热带喀斯特森林多花白头树木质部年内生长动态及其对环境因子的响应
2024-01-17刘亚男韦小练范泽鑫陈礼敏林友兴付培立
刘亚男,韦小练,范泽鑫,陈礼敏,林友兴,付培立
1 中国科学院西双版纳热带植物园,热带森林生态学重点实验室,勐腊 666303 2 中国科学院大学,北京 100049 3 云南哀牢山森林生态系统国家野外科学观测研究站,景东 676209 4 中国科学院核心植物园,勐腊 666303 5 三峡大学生物与制药学院,宜昌 443002
热带森林保存了59%的全球森林碳库,在增强全球陆地碳吸收和维持碳平衡方面具有重要作用[1-2]。近年来,亚马逊森林的碳汇持续下降,随着热带森林的碳汇趋向饱和,热带森林可能由碳汇转变为碳源[3]。木质部生长是森林生态系统固碳的重要方式,形成层细胞通过分裂和分化产生木质部细胞并表现为树木的径向生长,其生长过程受诸多环境因子的影响[4]。通常认为,热带地区的树木生长主要受水分状况的影响。在泰国的研究发现,形成层活动与相对湿度呈显著正相关[5]。在厄瓜多尔南部湿润的山地雨林中,形成层活动也主要受水分限制[6]。在巴西东南部的半落叶林内,雨季的树木径向增量显著大于旱季[7]。然而,热带树木生长不仅受水分影响,还与温度和其它环境因子有关。在巴西东南部的研究表明,热带雨林树木的径向生长和形成层活动与温度和日长呈显著的正相关关系[8-9]。但也有研究发现,热带树木的形成层活动与温度不相关[10]。因此,树木生长与环境因子的关系十分复杂,热带地区有关木质部生长及其调控因子的研究相对较少,理解热带树木年内生长动态及其对环境因子的响应尚且具有一定挑战性[11-12]。
喀斯特是一种脆弱的生境,由于土壤浅薄、保水能力差,植物在喀斯特生境主要利用土壤水和储存在基岩中的水,生长在喀斯特生境的植物容易受水分胁迫的影响[13-14]。在西双版纳地区,喀斯特主要分布于海拔600-1600 m的东南地区[14]。与同区域的热带季节雨林相比,西双版纳喀斯特森林的植物具有更强的叶片保水能力和更高的光合水分利用效率[15-16]。对热带喀斯特森林树木径向生长的研究表明,树木的径向生长与降水和相对湿度显著正相关[17-19]。与热带季节雨林相比,喀斯特森林红椿(Toonaciliata)的径向生长对干旱更加敏感[17]。但是,目前对热带喀斯特森林树木木质部生长的研究尚且有限,并且不同树种对环境因子的响应不同[17,20]。监测热带喀斯特森林树木木质部生长及其对环境因子的响应,不仅可以揭示树木的年内生长动态,而且可以预测气候变化背景下热带喀斯特森林的生长趋势。多花白头树(Garugafloribundavar.gamblei)为橄榄科植物,分布在亚洲热带地区[21],是西双版纳热带喀斯特森林的常见落叶树种。本研究采用生长环和微树芯两种方法监测了多花白头树在2020-2021年的树干径向变化和木质部生长动态,然后分别将月径向生长量和木质部生长与环境因子进行相关分析,旨在(1)揭示热带喀斯特森林多花白头树树干径向变化和木质部生长的年内动态;(2)明确热带喀斯特森林多花白头树木质部生长对环境因子的响应。
1 材料与方法
1.1 研究样地
本研究在云南省西双版纳傣族自治州勐仑镇中国科学院西双版纳热带植物园绿石林景区(21°54′N, 101°16′E, 海拔680 m)中进行(图1)。该地区位于云南南部,地处热带边缘,1963-2019年的平均温度为22.0℃,平均年降水量为1479.6 mm,受热带季风气候的影响,有明显的旱季(11月-次年4月)和雨季(5-10月)之分。绿石林景区内石灰岩丛生,呈现喀斯特地貌,其植被属于热带季节性湿润林,呈半常绿季相,具有一定比例的落叶树种[22]。

图1 研究样地的地理位置和野外实验照片Fig.1 Location of study site and pictures of field experiment
1.2 树木生长动态的监测
选择3棵树干直立、生长状况良好且胸径相似的多花白头树(平均胸径:(58.9±3.8)cm),在胸高处安装生长环,标记初始刻度线(图1)[23]。安装生长环前,去除部分树皮,降低树皮含水量对树干径向变化的影响。在2020年2月至2021年12月用游标卡尺测量树干胸围变化。2020年的2、10和12月分别在月初和月末各测量一次树干胸围变化,其它月份则在月初测定一次。2021年的1、5和6月仅在月初测定一次,其它月份分别在月初和月末各测定一次。由于受天气的影响,2021年 8月没有测量。
使用微型生长锥(Trephor, 意大利)在2020年2月至2021年12月每两周沿树干一侧在胸高处呈“Z”型螺旋上升采集微树芯(图1)[24],其中2021年1、5和6月仅采样一次,2月没有采样。将采集的微树芯放入50%酒精溶液中固定,经乙醇甘油溶液软化、不同浓度梯度的酒精脱水、柠檬烯透明、浸蜡、石蜡包埋、切片,制作永久装片,并使用带有偏光模块的显微镜(DM2500, Leica, 德国)拍照。每张照片中选择三个位置用ImageJ(https://imagej.nih.gov/ij/)分别测量形成层区域、扩大细胞区域、细胞壁加厚区域和成熟细胞区域的宽度(图2)。多花白头树的早材木质部细胞腔大、细胞壁薄,晚材木质部细胞腔小、细胞壁厚,年轮边界处具有排列整齐的薄壁细胞(图2)。

图2 多花白头树的微树芯解剖图Fig.2 Microcoring anatomical pictures of Garuga floribunda var. gambleiPH:韧皮部;CZ:形成层区域;EZ:扩大细胞区域;WZ:细胞壁加厚区域;MZ:成熟细胞区域;红色虚线表示年轮边界;刻度尺为200 μm
1.3 气象数据的获取
本研究使用的气象数据均来自中国科学院西双版纳热带雨林生态系统定位研究站(以下简称版纳生态站)。光合有效辐射、降水、相对湿度、日平均气温、日最低气温和日最高气温数据来自版纳生态站的综合气象观测场,距绿石林景区约3 km。20 cm土壤含水量的数据来自版纳生态站石灰山季节雨林调查点,距研究样地较近。
1.4 数据分析
将生长环监测的树干胸围动态转化为树干半径变化,使用McMahon和Parker提出的逻辑斯蒂模型(Logistic model)模拟树干径向变化,判断生长开始和结束时间[25]。逻辑斯蒂模型假设径向生长达到生长曲线下渐近线的2.5%时为生长开始时间,径向生长达到生长曲线上渐近线的97.5%时为生长结束时间。
广义可加模型(Generalized additive models, GAMs)为数据驱动模型,可以对木质部生长的动态变化进行拟合[26]。本研究使用广义可加模型拟合木质部生长过程中各分化阶段的动态过程。由于每次采样的木质部生长速率不同,采用前一年的年轮宽度对木质部生长的各分化阶段进行标准化[27]。同时,将扩大细胞的首次出现定义为木质部生长开始,细胞壁加厚的停止定义为木质部生长结束。
分别将每月和采样前10天的光合有效辐射、降水、相对湿度、日平均气温、日最低气温、日最高气温、饱和水汽压亏缺和20 cm土壤含水量的均值与月径向生长量和扩大细胞区域宽度以及细胞壁加厚区域宽度做Spearman 相关。饱和水汽压亏缺的计算采用“plantecophys”包中的RHtoVPD函数,根据温度和相对湿度计算饱和水汽压亏缺[28]。所有的数据分析和作图均在R 4.0.5中进行[29],并用Adobe Illustrator 2020对图进行修改。
2 结果与分析
2.1 环境因子的变化
研究区域的温度、降水、相对湿度和20 cm土壤含水量具有明显的季节性(图3)。2020年的平均气温为23℃,2021年的平均气温为22.7℃;2020年的总降水量为1206 mm,2021年的总降水量为1272 mm。这两年的年降水量均低于该地区的多年平均降水量。2020年雨季的降水为1045.2 mm,2021年雨季的降水为958.8 mm。与2021年相比,2020年的1-3月更为干旱,相对湿度在2020年3月底达到最低。20 cm土壤含水量的变化与降水变化相对一致,均呈雨季高、旱季低的趋势。
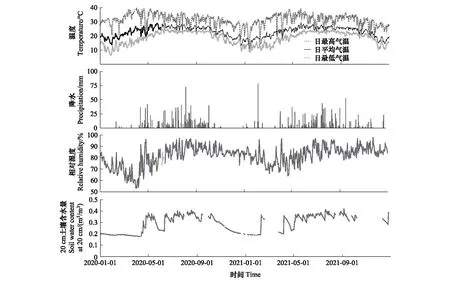
图3 西双版纳勐仑地区2020-2021年温度、降水、相对湿度以及20 cm土壤含水量的日变化Fig.3 Variations in daily temperature, precipitation, relative humidity and soil water content at a depth of 20 cm during 2020-2021 in Menglun, Xishuangbanna
2.2 树干径向变化
多花白头树的树干径向变化在不同年份和不同个体的总体趋势相对一致,但不同个体径向增量的变异较大,同一个体在不同年份的径向增量也存在差异(图4)。通过比较树干径向变化的月动态发现(图4),多花白头树在2020年和2021年的径向增量均主要集中在6-9月,其中6月的径向增量最大。受树干水分亏缺的影响,树干径向增量在旱季会出现负值。

图4 多花白头树2020-2021年的树干径向变化和径向生长月动态Fig.4 Stem radius variation and monthly radial growth of Garuga floribunda var. gamblei during 2020-2021D1、D2、D3分别表示不同植株;曲线为GAMs拟合的平滑趋势线;图中数据为平均值±标准误(n=3)
逻辑斯蒂模型拟合的结果表明,多花白头树在2020年和2021年的生长开始时间分别为4月底(DOY:114.7±26.7)和4月中旬(DOY:103.7±11),生长停止时间分别为8月初(DOY:215±51.2)和8月底(DOY:232±18.7)。2020年的生长开始时间晚于2021年,但2020年的生长结束时间早于2021年。多花白头树在2021年的生长持续时间更长,但2020年的径向生长量((3.30±2.91)mm)大于2021年((2.36±0.81)mm)。
2.3 木质部生长
用GAMs分别拟合木质部生长的各分化阶段(图5)。结果表明,形成层区域宽度的变化没有明显趋势,扩大细胞区域宽度和细胞壁加厚区域宽度均呈先增加后减少的“钟形”曲线模式,成熟细胞区域宽度变化呈“S”型曲线。扩大细胞区域在2020年2月底出现,6月中下旬出现最大峰值,10月之后消失;细胞壁加厚区域在2020年4月初出现,7月中下旬出现最大峰值,9月底消失。扩大细胞区域和细胞壁加厚区域的最大峰值出现时间不同,细胞壁加厚区域的最大峰值约滞后1个月。由于2021年5-6月的采样间隔较长,扩大细胞和细胞壁加厚细胞出现的时间间隔较大。扩大细胞在2021年4月中旬出现,细胞壁加厚细胞则在5月底出现,并且扩大细胞区域的最大峰值不明显,细胞壁加厚区域的最大峰值出现在2021年6月中下旬,扩大细胞在9月之后消失,细胞壁加厚细胞在9月较低直至10月消失。无论是扩大细胞区域还是细胞壁加厚区域,它们在2020年的宽度均大于2021年的宽度。受采样位置的影响,成熟细胞区域的宽度在木质部生长结束后仍然不断波动。

图5 多花白头树2020-2021年形成层区域、扩大细胞区域、细胞壁加厚区域和成熟细胞区域的宽度变化Fig.5 The variations in the thickness of cambial zone, enlarging zone, wall-thickening zone and mature zone of Garuga floribunda var. gamblei during 2020-2021图中数据为平均值±标准误(n=3)
通过比较多花白头树在2020年和2021年的木质部生长物候参数发现(图6),多花白头树在2020年和2021年的木质部生长开始时间分别为3月底(DOY:87±45)和4月中旬(DOY:103.7±11.5),2020年和2021年的木质部生长结束时间分别为9月底(DOY:269.7±30)和10月中旬(DOY:288±10.4)。多花白头树在2020年的木质部生长开始时间和结束时间均稍早于2021年,但在统计学上均不具有显著差异。2020年的木质部生长量为(2.87±1.46)mm,2021年的木质部生长量为(2.98±1.02)mm,二者也不存在显著差异。

图6 多花白头树2020-2021年木质部生长物候参数的箱型图Fig.6 Boxplots of the xylem growth phenology parameters of Garuga floribunda var. gamblei during 2020-2021
2.4 木质部生长对环境因子的响应
将生长环在2020-2021年监测的月径向生长量与环境因子做Spearman相关(图7)。结果表明,2020年和2021年的月径向生长量均与降水显著正相关,2021年的月径向生长量还与光合有效辐射、日平均气温以及日最低气温显著正相关。
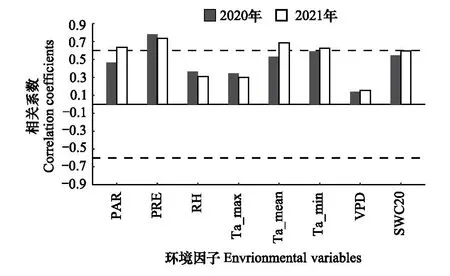
图7 多花白头树2020-2021年月径向生长量与环境因子的相关系数Fig.7 Correlation coefficients between monthly radial growth of Garuga floribunda var. gamblei and environmental variables during 2020-2021PAR:光合有效辐射;PRE:降水;RH:相对湿度;Ta_max:日最高气温;Ta_mean:日平均气温;Ta_min:日最低气温;VPD:饱和水汽压亏缺;SWC20:20 cm土壤含水量;图中虚线为相关系数的临界值(P<0.05)
分别将2020-2021年扩大细胞区域宽度和细胞壁加厚区域宽度与采样前10天的环境因子做Spearman相关(图8)。结果表明,2020年和2021年的扩大细胞区域宽度与降水、日平均气温和日最低气温显著正相关,2020年的扩大细胞区域宽度还与日最高气温显著正相关。2020年和2021年的细胞壁加厚区域宽度与降水、相对湿度、日平均气温、日最低气温和20 cm土壤含水量显著正相关,并且2021年的相关系数更高。

图8 多花白头树2020-2021年扩大细胞区域宽度和细胞壁加厚区域宽度与环境因子的相关系数Fig.8 Correlation coefficients between the thickness of enlarging zone and wall thickening zone of Garuga floribunda var. gamblei and environmental variables during 2020-2021
3 讨论
结果表明,热带喀斯特森林多花白头树木质部开始生长时间为3月底至4月初,结束生长时间为9月底至10月中旬。韦小练等发现,该区域热带季节雨林多花白头树形成层活动的开始时间为3月初,结束时间为9月中下旬,与本研究结果相对一致[30]。对该区域红椿的研究表明,树木的径向生长主要发生在3-10月,受水分亏缺的影响,热带喀斯特森林的生长持续时间小于热带季节雨林[17]。此外,多花白头树在2020年和2021年的最大径向生长速率均发生在6月,扩大细胞区域也在6月出现峰值。对该区域热带季节雨林多花白头树径向生长和形成层活动的研究表明,其最大径向生长速率也出现在6月[30]。西双版纳地区的旱季(11月-次年4月)降水仅为全年15%左右,该地区树木的径向生长主要集中在雨季,径向生长速率在雨季初期(5-6月)达到最高。另一方面,最大生长速率也可能受日照长度的影响,瑞士温带森林树木径向生长的最大速率主要出现在日照最长的夏至日前后[31]。与2021年相比,多花白头树不同个体在2020年的生长变异较大,2020年的径向生长量略高于2021年。但是2020年和2021年的木质部生长量较为接近,这可能是因为生长环测定的径向生长还包含了树皮以及水分收缩膨胀的影响,并且树干径向变化不能直接反映细胞壁加厚阶段的信息[32]。
扩大细胞区域和细胞壁加厚区域在2020年的最大峰值出现时间不同,细胞壁加厚区域最大峰值的出现时间约滞后1个月。Cuny等人通过比较针叶树扩大细胞和细胞壁加厚细胞的年内动态发现,扩大细胞区域的峰值出现时间早于细胞壁加厚区域,即树干木质部生物量的积累滞后于树干胸围增加,这种滞后现象普遍存在于寒温带、亚高山和地中海森林[33]。然而,对西双版纳热带季节雨林多花白头树[30]和红椿[17]的研究发现,扩大细胞区域和细胞壁加厚区域的最大峰值不存在时滞。在热带喀斯特森林中,红椿的这种滞后现象也并不明显[17]。在本研究中,多花白头树扩大细胞区域和细胞壁加厚区域在2021年的峰值也未出现明显时滞,因此热带地区树木径向生长和生物量积累是否存在时滞仍需进一步探究。
形成层细胞分裂后的细胞扩大需要膨压驱动并依靠游离的糖和氨基酸维持膨压,因此产生扩大细胞需要良好的水热条件[34]。Huang等发现,光周期与温度在调控北半球针叶树的木质部生长开始时间方面发挥了主要作用[35]。在我国祁连山中部的研究表明,祁连圆柏(Sabinaprzewalskii)木质部分化的开始时间主要受温度影响[36]。对中国亚热带地区杉木(Cunninghamialanceolata)和台湾松(Pinustaiwanensis)的研究发现,生长季初期的树干径向生长量与温度呈正相关[37-38]。在中国亚热带湿润季风气候区,春季温度的升高有助于提前打破樟树(Cinnamomumcamphora)形成层细胞的休眠[39]。这些结果与本研究中扩大细胞区域宽度以及细胞壁加厚区域宽度均和温度呈显著正相关的研究结果一致。落叶树种春季物候的启动与温度介导的植物体内淀粉和可溶性糖转换有关[40]。本研究发现2020年扩大细胞区域宽度与日平均气温和日最高气温的相关系数高于2021年,但与日最低气温的相关系数却低于2021年,这可能与2021年的平均温度(22.7℃)低于2020年(23.0℃)有关。多花白头树在2020年的木质部生长开始时间早于2021年,2020年1-3月的平均气温(20.1℃)大于2021年(18.9℃),2020年1-3月的降水量(22.6 mm)远低于2021年(138 mm),同时2020年1-3月的平均相对湿度(70%)低于2021年(78%)。因此,2021年1-3月较好的水分条件并没有使形成层活动提前,多花白头树在生长季早期的木质部生长启动可能还受温度或其它环境因子的影响。
热带喀斯特森林多花白头树的木质部生长主要受水分限制,多花白头树在2020年和2021年的月径向生长量、扩大细胞区域宽度和细胞壁加厚区域宽度均与降水呈显著正相关。在西双版纳的研究表明,热带喀斯特森林苦楝和红椿的径向生长也主要受水分限制[17,19]。Kaewmano等人对热带喀斯特森林红椿的研究还发现,扩大细胞区域宽度和细胞壁加厚区域宽度均与降水显著正相关,这与本研究结果一致[17]。细胞壁加厚区域的宽度除了与降水呈正相关,还与相对湿度和20 cm土壤含水量呈显著正相关。与扩大细胞区域相比,细胞壁加厚区域可能主要取决于碳水化合物的可利用性,而较好的水分条件有利于植物光合作用进行[33-34]。Wang 等人对我国西北地区三种针叶树的研究发现,水分状况较好时,植物在夏秋季积累的碳同化产物除了影响当年的树木生长,还可能影响次年树木生长[41]。对印度尼西亚四种阔叶树的研究表明,形成层活动主要受降水影响,旱季频繁的降水使树木形成层一直处于活跃状态[42]。然而,在降水充足、温度和光周期季节性明显的法国东北部,扩大细胞的产生与日长正相关,细胞壁加厚区域与温度耦合[33]。Zweifel等人认为,树木生长主要取决于水分条件,其次是碳同化过程[43]。本研究发现2021年的细胞壁加厚区域宽度与环境因子(尤其是20 cm土壤含水量)的相关系数比2020年高,这主要是受雨季降水状况的影响,2020年雨季的降水较多,多花白头树的木质部生长主要集中在雨季。2020年5-9月的总降水量为965 mm,有降水天数为103天,而2021年5-9月的总降水量为884mm,有降水天数为93天。虽然这两年生长季的降水均较多,由于喀斯特森林的保水能力较差,从而可能使2021年的细胞壁加厚区域宽度更容易受水分限制。
4 结论
本研究利用生长环和微树芯两种方法对热带喀斯特森林多花白头树的径向生长和木质部生长进行了连续两年的监测。热带喀斯特森林多花白头树木质部生长的开始时间为3月底至4月初,结束时间为9月底到10月中旬。多花白头树的年生长量为3 mm左右,径向生长速率在6月最高,扩大细胞区域也在6月出现峰值。多花白头树的木质部生长主要受水分影响,月径向生长量、扩大细胞区域宽度和细胞壁加厚区域宽度均与降水显著正相关,细胞壁加厚区域还与相对湿度和20 cm土壤含水量正相关。本研究有助于理解热带喀斯特森林树木生长对环境因子的响应,同时为热带树木年轮气候学提供参考。
致谢:感谢Shankar Panthi和杨绕琼师姐在地图绘制中给予的帮助,感谢马宏在野外实验中提供的帮助,感谢中国科学院西双版纳热带雨林生态系统定位研究站提供的气象数据。
