黄土丘陵区恢复草地土壤团聚体组成及全氮分布特征
2024-01-17阿合江赛力克余舟昌刘育书郝洪剑田兴芳周荣磊
阿合江·赛力克,孙 琳,王 绒,余舟昌,刘育书,郝洪剑,田兴芳,周荣磊,张 伟
西北农林科技大学草业与草原学院,杨凌 712100
土壤团聚体是土壤颗粒通过有机质胶结而成的团块,是土壤基本结构单元,其稳定性与土壤氮储量密切相关[1]。不同粒级对土壤孔隙度、水土保持能力、土壤生物活动等具有不同的作用,并且会受到内部的有机物[2]、微生物群落因素影响[3],这样会导致不同粒级当中氮累积量与其对应的粒级稳定性产生差异,这种不同粒径氮的累积差异会对全土氮储量贡献产生影响[4]。所以从团聚体尺度研究不同粒径分布与其中氮累积的变化,对于揭示土壤氮动态和稳定性机理具有重要意义。
草地生态系统作为陆地生态系统重要的组成部分,其固氮能力和动态变化在全球氮循环中扮演着非常重要的作用[5]。在国家实施“退耕还林(草)”及封山禁牧政策以来,黄土高原退化天然草地得以恢复,植被覆盖度大幅提升,生态环境大为改善,势必会对土壤氮储量及土壤团聚体的形成和稳定性产生影响[6]。目前,学术界在草地恢复对土壤团聚体氮固存的影响研究方面主要存在两种不一致的观点。大量研究认为,草地恢复首先增加土壤大粒级团聚体(>0.25 mm)氮含量,微粒级团聚体(<0.25 mm)氮含量的增加相对较慢,且大粒级团聚体对土壤氮累积的相对贡献大于微粒级团聚体[4,7];但是,也有学者发现草地恢复首先增加土壤微粒级团聚体氮含量,并且土壤微粒级团聚体对土壤氮累积的贡献大于大粒级团聚体[8]。因为团聚体的形成和稳定是复杂的过程,依赖于与植物属性(例如植被类型、植物根系)和土壤属性(例如土壤有机质、微生物活性)相关的各种生物和非生物因素[2,9]。因此,基于不同恢复年限草地土壤团聚体构成及其氮含量差异,阐明不同粒级团聚体对土壤氮固持的相对贡献及其影响因素,对于揭示草地恢复过程中土壤氮的动态特征、累积规律和稳定性维持机理具有重要意义。
黄土高原丘陵区撂荒草地是经过长期的恢复和保护,在该区形成了时间跨度大、保存完整、受干扰较小的天然草地恢复系列,为开展长期草地恢复中土壤氮动态及稳定机制研究提供了理想的平台[10]。有关该区域草地恢复对团聚体氮累积动态及稳定性的研究较少,且尚不清楚长期草地恢复过程中,植物、微生物等对土壤团聚体氮累积动态及稳定性的影响。因此,本研究以10a、20a、30a、40a和50a恢复草地和农田(CK)为研究对象,探究草地自然恢复过程中土壤团聚体氮累积的变化,以及分析植物叶片碳氮含量、凋落物、根系和团聚体稳定性等与团聚体氮累积的关系,明确自然草地恢复中土壤团聚体分布动态及氮累积与影响因素,为指导土壤养分管理和生态可持续发展提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省延安市安塞区五里湾流域(36°51′-36°53′N,109°20′-109°22′E)(图1)。该区气候类型属于暖温带半干旱大陆性季风气候,年平均气温为8.8°C,年平均降水量535 mm,降雨集中在7-9月,约占全年降雨量的70%。无霜期在4月初至9月末之间,研究区主要的土壤类型为黄绵土,其粘聚力弱、渗透性高、保水性低,导致该区域土质疏松、抗侵蚀能力差、土壤贫瘠[11]。过去几十年,该地区实施了退耕还林还草措施,以恢复退化的生态系统,植被类型由传统的农田转变为撂荒草地,从中国科学院地理科学与资源研究所数据共享中获取基于Landsat 8遥感影像的栅格数据,截至2021年,安塞县五里湾撂荒草地的面积有184.18 hm2,占该流域总面积约40%,是该地区主要的土地利用类型[12]。撂荒草地主要的优势种有茵陈蒿(Artemisiacapillaries)、铁杆蒿(Artemisiastechmanniana)、兴安胡枝子(Lespedezadavurica)、白羊草(Bothriochloaischaemum)、本氏针茅(StipabungeanaTrin)、甘草(Glycyrrhizauralensis)(表1)。
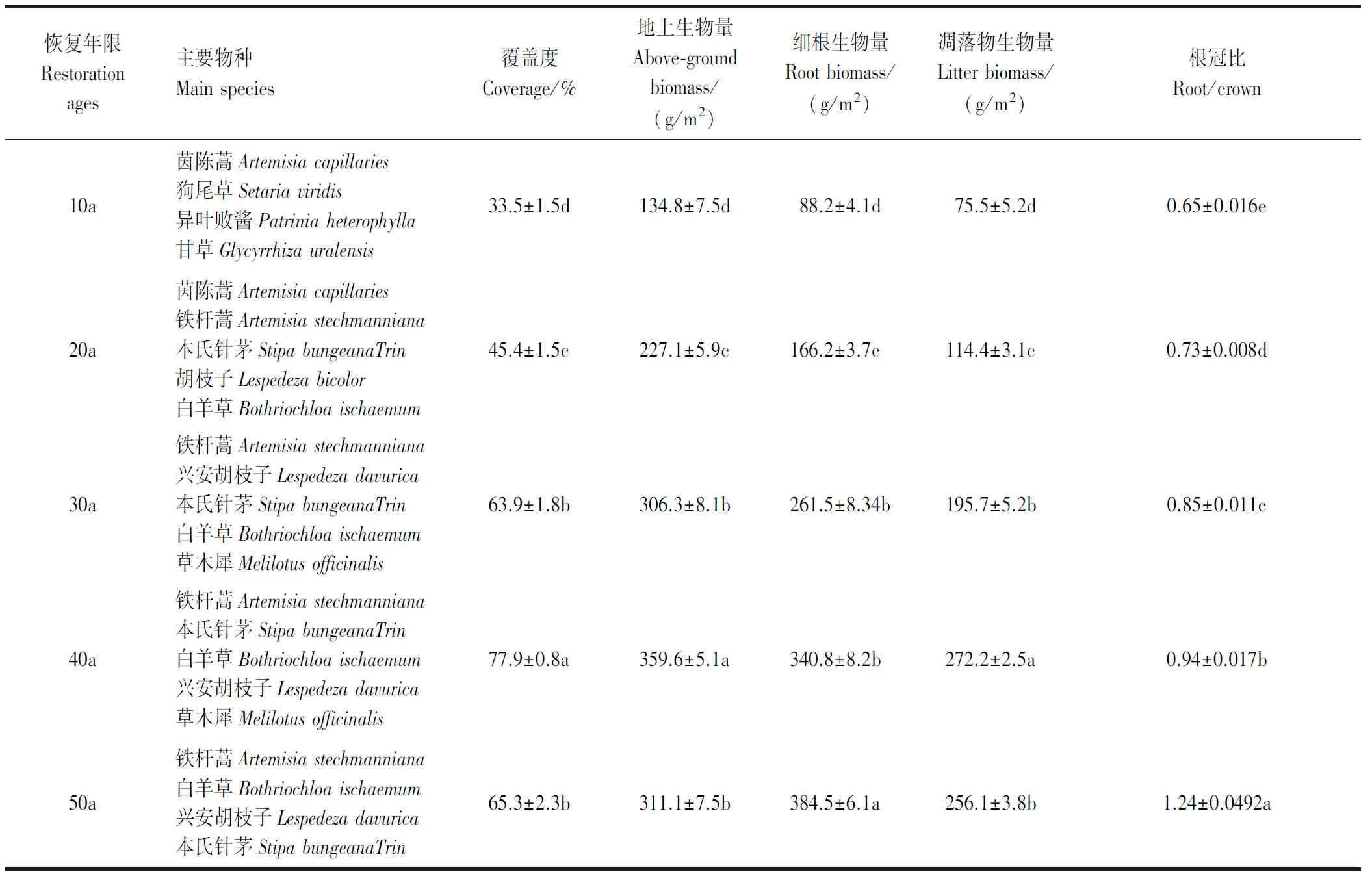
表1 草地不同恢复年限主要物种、盖度和产量特征Table 1 Main species, cover and yield characteristics of grassland at different restoration ages

图1 研究区地理位置图Fig.1 Geographical location map of the study area
1.2 试验设计
通过查阅当地林业部门造林资料,于2021年对研究区域实地考察后在五里湾流域内选取了干扰程度较低的不同年限撂荒草地,分别选择10a、20a、30a、40a和50a撂荒草地,以农田为对照(CK)。撂荒草地在恢复前均为农田,且与对照农田一样,主要种植玉米(Zeamays)和马铃薯(Solanumtuberosum),农作措施为玉米马铃薯轮作,种植期间为雨养农田,不进行人为管理措施,作物收获后移除全部地上植物,不进行土壤管理措施。撂荒草地各个恢复年限以及对照农田分别选择3块立地条件相似(海拔、坡向、坡位、坡度、土壤类型和地表结皮等)的独立重复样地,共计18个样地。每个样地内分别设置3个20 m×20 m的标准样方,便于进行后续的调查、采样以及指标测定,共计54个标准样方。
1.3 植被调查及植物样品采集
在每个20 m×20 m标准样方中按“S”形随机选取10个1 m×1 m的小样方,使用样方法调查记录样方内所有出现的草本植物的植物名,使用标尺测量样方内每株植物的高度,同种植物的盖度及密度,确定优势物种。在小样方中刈割植被称取地上生物量,使用根钻(内径为10 cm)钻取0-20 cm土层土芯3个,混合成为一个复合样本,淘洗过筛后分离出细根,收集小样方内的未分解的凋落物,将所有称重的植物、细根和凋落物样本在75°C下烘至恒重,称量记录。将干燥的植物样品粉碎,测定有机碳(OC)和全氮(TN)的含量。
1.4 土壤样品采集
在移除全部地上植被的10个1 m×1 m的小样方中选取5个,用铝盒(长×宽×高为:24 cm×15 cm×10 cm)取0-20 cm深度采集原状土样,带回实验室后将原状土壤沿其自然结构纹路轻掰成小土块,并将同一标准样方内的5份样品充分混匀,用于后续团聚体相关指标分析。用直径为5 cm的土钻在每个采样点钻取2个土壤样品,将同一标准样方内的10个土壤样品充分混匀后分成两份,一份用于测定土壤含水量、pH等基本理化性质。用环刀在每个采样点取1个土样,用于测定土壤容重。
1.5 植物多样性指数计算
草地植被多样性指数:本研究采用Margalef指数(M)、Shannon-Wiener指数(H)和Pielou均匀度指数(E)来表示群落的物种多样性[13-14],计算公式如下:
M=(S-1)/lnN
(1)
(2)
E=H/lnS
(3)
式中,N为全部种的个体总数;S为物种总数;Ni为i种的个体数;Pi为种i的多度比例,Pi=Ni/N。
1.6 理化性质和养分测定
土壤含水量采用烘干称重法测定,在105℃烘干至恒重后称重;土壤容重采用环刀法测定[15];使用pH计(PHS-3C)测量土壤pH(土壤∶水为1∶2.5);采用MS2000型激光粒度仪测定土壤颗粒组成(粘粒,粉粒,砂粒)(Malvern Mastersizer 2000, Malvern, Worcestershire, UK);植物、凋落物、细根和土壤中的有机碳(OC)含量均采用重铬酸钾-浓硫酸(K2Cr2O7-H2SO4)外加热容量法测定[15-16],全氮(TN)含量均采用硫酸钾:五水硫酸铜(K2SO4∶CuSO4·5H2O)(质量比10∶1)作为催化剂,并用浓硫酸(H2SO4)消煮后用全自动凯氏定氮仪(KDY-9830)测定[17]。
1.7 数据处理
团聚体粒级分级:土壤团聚体采用分级湿筛法,收集4个粒级团聚体土壤,分别为> 2mm、0.25-2 mm、0.053-0.25 mm和<0.053 mm。将土壤团聚体稳定性指标采用平均重量直径(MWD)、几何平均直径(GMD)[18]。分别按照以下公式计算:
(4)

(5)
式中,di是筛分出来的任意粒径范围内团聚体的平均直径(mm);wi是任意粒径范围内团聚体的质量占土样干重的百分比(%)。
土壤全氮储量(kg/m2)计算公式为:
(6)
式中,TNs是土壤全氮储量;STN是土壤全氮含量(g/kg);D是土壤深度(cm);BD为土壤容重(g/cm3)。
土壤各粒级团聚体氮储量(g/m2)计算公式为:
(7)
式中,ANSi为各粒级团聚体氮储量;ANCi为第i粒级团聚体氮含量(g/kg);wi为第i粒级占比(%)。
各粒级团聚体氮储量变化的驱动因素可分为各粒级TN含量的变化F1和各粒级占比的变化F2[19-20],因此假设一个特定的粒级氮储量的变化是该粒级TN含量的变化主导驱动则F1>F2,反之,一个特定的粒级氮储量的变化是该粒级占比的变化主导驱动则F1 F1=M×ΔC (8) F2=ΔM×C (9) 式中,M为草地恢复前团聚体占比,ΔM为草地恢复引起的团聚体占比的变化值,C为草地恢复后的团聚体氮含量,ΔC为草地恢复引起的团聚体氮含量变化差值。 所有数据的分布采用Shapiro-Wilk检验,所有变量均服从正态分布,并满足进一步方差分析的假设。同一指标(植物地上生物量、植物和土壤OC、TN、团聚体稳定性、粒度分布、各粒级TN含量和储存量)在不同年限间(CK、10a、20a、30a、40a和50a)差异显著性采用单因素方差分析(One-way ANOVA,LSD)进行检验(P<0.05)。通过冗余分析,探讨了团聚体氮积累与植物指标和土壤理化性质的关系。通过Spearman相关分析,确定了土壤全氮储量、团聚体稳定性和各粒级氮储量之间的联系。使用Origin 2021软件进行统计分析和绘图,使用Canoco 5进行冗余分析并绘图。 2.1.1不同恢复年限草地群落物种组成和生物量特征及多样性特征 如表1所示,5个不同恢复年限草地群落主要物种属菊科、禾本科、豆科和败酱科,10a至50a的恢复过程中,草地群落经历了茵陈蒿→茵陈蒿+铁杆蒿→铁杆蒿+兴安胡枝子→铁杆蒿+本氏针茅→铁杆蒿+白羊草的演替过程。此外,植物群落特征在草地恢复过程中不断变化,其中草地群落覆盖度、地上生物量和凋落物生物量变化范围分别为33.5-77.9%、134.8-359.6 g/m2和75.5-272.2 g/m2,且恢复0-40a均呈现增加趋势,并在40a达到最大值,40-50a呈现降低趋势。与覆盖度、地上生物量和凋落物生物量变化规律不同,草地群落细根生物量和根冠比随着恢复年限的增加呈显著增加趋势,与恢复10a相比,恢复20a、30a、40a和50a草地群落地下生物量和根冠比分别增加了88.4%、196.5%、286.4%、335.9%(地下生物量)和12.3%、30.8%、44.6%、90.8%(根冠比)。在50a的恢复过程中,草地群落物种多样性发生明显变化(图2),且均呈现先上升后下降的趋势,且最大值出现在30a。通过分析可知恢复10-50a草地群落多样性指数满足抛物线拟合方程,分别为:y=-0.01952x2+1.3981x-4.133(R2=0.779)(物种数)、y=-0.00151x2+0.10604x+1.882(R2=0.828)(Margalef指数)、y=-0.00236x2+0.1728x+0.36533(R2=0.835)(Shannon-Wiener指数)和y=-0.00038x2+0.02857x+0.63343(R2=0.894)(Pielou均匀度指数)。 图2 不同恢复年限草地群落多样性指数Fig.2 Diversity index of grassland communities in different restoration ages 2.1.2不同恢复年限草地的植被、根系和凋落物养分特征 如表2所示,不同恢复年限草地植被的地上部分、根系和凋落物OC和TN含量变化明显。不同恢复年限草地植物、细根和凋落物碳含量变化范围分别为363.7-456.6 g/kg、329.17-427.37 g/kg和296.92-404.89 g/kg。而植物、细根和凋落物TN含量变化范围分别为12.05-24.51 g/kg、9.74-20.1 g/kg和7.36-18.23 g/kg。草地植物、细根和凋落物碳TN含量随年限增加呈现先上升后下降的趋势,不同的是植物、细根和凋落物OC含量最高值出现在40a,而TN含量最高值出现在30a。草地植物、细根和凋落物碳氮比变化范围分别为17.45-30.34、19.85-33.85和20.75-40.39。且随恢复年限增加呈现先下降后上升的趋势,最小值均出现在30a。 表2 不同恢复年限草地的植被、根系和凋落物养分Table 2 Plant, Root and Litter nutrients of grassland at different restoration ages 不同恢复年限草地土壤理化性质与农田相比有显著的差异(表3)。其中土壤pH和土壤容重逐年降低,恢复10-50a草地与农田相比pH和容重降幅范围分别为0.67%-4.29%和6.29%-17.32%。土壤含水量变化范围为11.57%-17.01%,恢复0-20a呈现下降趋势,20-50a呈现上升趋势,最低值和最高值分别出现在20a和50a。草地恢复显著增加了土壤粘粒含量,恢复10-50a草地与农田相比土壤粘粒含量增加了6.52%-64.84%。草地恢复过程中土壤粉粒含量呈现先上升后下降的趋势,在30a达到最高值,与之相反土壤沙粒含量呈现先下降后上升的趋势。草地恢复过程中土壤有机碳含量呈现增加趋势,恢复10-50a草地与农田相比土壤有机碳含量增加了37.14%-332.38%。草地恢复过程中的土壤TN含量呈现先上升后下降的趋势,恢复40a达到最大值,而草地恢复中TN储量同土壤氮含量变化相似在40a达到最高值,恢复10-50a草地与农田相比TN储量值增加范围为8.10-118.92%。 表3 不同恢复年限土壤理化性质Table 3 Physicochemical properties of soil at different restoration ages 有图3所示,草地恢复影响了土壤团聚体分布特征。在农田地团聚体分布特征为微团聚体(0.053-0.25 mm)>中团聚体(0.25-2 mm)=粘粉粒团聚体(<0.053 mm)>大团聚体(> 2mm)。在草地恢复10-40a之间土壤微团聚体占据主导地位,但其他粒级团聚体分布各不相同,在10a中团聚体>大团聚体>粘粉粒团聚体,20a中团聚体=大团聚体>粘粉粒团聚体,30a和40a大团聚体>中团聚体>粘粉粒团聚体,到了50a大团聚体>微团聚体>中团聚体>粘粉粒团聚体。50a的恢复过程中,草地土壤大团聚体和中团聚体占比呈上升趋势,草地恢复50a大团聚体占主导地位,占比为36%。与之相反微团聚体在草地恢复过程比例下降。而粘粉粒团聚体在草地恢复过程中呈现先下降后上升的趋势。土壤团聚体稳定性指标MWD和GMD随草地恢复年限增加而增加(图3)。草地恢复10a、20a、30a、40a和50a土壤团聚体MWD和GMD指数分别比农田提高50.12%、96.66%、155.07%、188.79%、210.69%和14.60%、25.77%、39.73%、49.55%、55.48%。土壤团聚体稳定性排序依次为50a>40a>30a>20a>10a>ck(P<0.05)。 图3 团聚体粒级分布和稳定性Fig.3 Particle size distribution and stability of aggregate图中不同大写字母表示不同恢复年限之间团聚体占比差异;不同小写字母表示各恢复年限当中团聚体占比以及稳定性差异 由图4可知,草地恢复使各粒级团聚体总TN含量和TN储量明显发生变化。农田、10a、20a、30a、40a和50a各粒级团聚体TN含量变化范围分别为0.2-0.33 g/kg、0.21-0.27 g/kg、0.34-0.43 g/kg、0.55-0.64 g/kg、0.65-0.75 g/kg和0.63-0.71 g/kg,中团聚体TN含量最高。大团聚体TN含量变化范围为0.24-0.67 g/kg,随恢复年限呈现先下降后上升的趋势,在10a为最小值。中团聚体和微团聚体在草地恢复过程中的TN含量变化,均呈现先下降后上升再降低的趋势,变化范围分别为0.27-0.74 g/kg和0.24-0.73 g/kg最低值和最高值分别出现在10a和40a。与上述三个粒级团聚体不同,粘粉粒团聚体TN含量变化范围为0.2-0.64 g/kg,呈现先上升后下降的趋势,在40a为最大值。农田、10a和20a各粒级团聚体TN储量均呈现微团聚体>中团聚体>大团聚体>粘粉粒团聚体的特征,在草地恢复30a和40a大团聚体TN储量高于中团聚体和粘粉粒团聚体TN储量,TN储量依然低于微团聚体。不同的是50a团聚体TN储量依次为大团聚体>微团聚体>中团聚体>粘粉粒团聚体。而大团聚体TN储量逐年增加,恢复10-50a与农田相比TN储量增加范围为70.19-1491.30%。中团聚体TN储量在50a恢复过程当中呈现先上升后下降的趋势,在40a为最大值,与农田相比恢复10-50a草地TN储量增加范围为32.44-353.62%。微团聚体TN储量在0-10a、10-30a和30-50a分别呈现先下降后上升再下降的趋势。不同恢复年限草地粘粉粒团聚体TN储量在10a为最低值,20a、30a、40a和50a与10a相比粘粉粒团聚体TN储量分别增加了65.36%、224.18%、325.49%和357.79%。 图4 不同恢复年限草地团聚体全氮含量(点线图)和储量(直方图)Fig.4 Total nitrogen content (line chart) and stock (histogram) of grassland aggregates at different restoration agesTNs:全氮储量 total nitrogen stocks;TN:全氮含量 total nitrogen;图中不同小写字母表示各粒级团聚体在不同恢复年限的全氮储量存在显著差异(P<0.05) 在草地恢复过程中,草地植被和土壤理化性质对团聚体组分、稳定性和氮累积产生影响。图5为基于冗余分析的不同年限恢复草地植被特征和土壤理化性质与团聚体组分及稳定性和TN储量的相关性,其结果表明,植被、根系、凋落物生物量和碳含量与大团聚体、中团聚体、粘粉粒团聚体占比和团聚体稳定性呈正相关关系,植被、根系、凋落物氮含量也与大团聚体、中团聚体、粘粉粒团聚体占比和团聚体稳定性相关。不同的是微团聚体占比与上述植被特征呈现负相关关系,与植物碳氮比相关。如图5所示各粒级TN储量除植被、凋落物碳氮比之外的植被特征有相关关系,其中植被、根系、凋落物生物量和碳含量与大团聚体、中团聚体、粘粉粒团聚体TN储量相关,而微团聚体TN储量与植被、根系、凋落物氮含量相关。草地恢复过程当中大团聚体、中团聚体、粘粉粒团聚体占比和团聚体稳定与土壤氮含量、有机碳含量、含水量和粘粒含量相关。微团聚体占比与土壤pH、容重和砂粒含量相关。微团聚体TN储量与粉粒、土壤氮、有机碳和粘粒含量相关,大团聚体、中团聚体、粘粉粒团聚体TN储量与土壤TN、有机碳、粘粒、粉粒含量和含水量相关,其中大团聚体、中团聚体氮储量与土壤氮含量相关。 图5 植被特征和土壤理化性质与团聚体组分和全氮储量关系Fig.5 Vegetation characteristics and soil physicochemical properties in relation to aggregates fraction and TN storageMac:大团聚体占比 macro aggregates content;Mec:中团聚体占比 medium aggregates content;Mic:微团聚体占比 micro aggregates content;S+Cc:粘粉粒团聚体占比 silt+clay aggregates content;MWD:平均重量直径mean weight diameter;GMD: 几何平均直径geometric mean diameter;MaTNs:大团聚体氮储量 macro aggregates total nitrogen stocks;MeTNs:中团聚体氮储量 medium aggregates total nitrogen stocks;MiTNs:微团聚体氮储量 micro aggregates total nitrogen stocks;S+CTNs:粘粉粒团聚体氮储量 silt+clay aggregates total nitrogen stocks;PlantB:植物生物量 plant biomass;PlantC:植物碳含量 plant carbon;PlantN:植物氮含量 plant nitrogen;RootB:根系生物量 root biomass;RootC:根系碳含量 root carbon;RootN: 根系氮含量 root nitrogen;LitterB:凋落物生物量 litter biomass;LitterC:凋落物碳含量 litter carbon;LitterN:凋落物氮含量 litter nitrogen;BD:容重 bulk density;SWC:土壤含水量 soil water content;Clay c: 粘粒含量 clay content;Silt c:粉粒含量 silt content;Sand c:砂粒含量 sand content;SOC:土壤有机碳含量 soil organic content;STN:土壤全氮含量 soil total nitrogen content 相关性分析表明(图6),土壤TN储量与团聚体分布和稳定性显著相关(P<0.05)。其中大、中和粘粉粒团聚体占比以及团聚体稳定性指数与土壤氮储量呈现显著正相关(P<0.05),而与微团聚体占比呈现显著负相关(P<0.05)。土壤TN储量与大、中和粘粉粒团聚体TN储量呈现显著正相关(P<0.05),但与微团聚体TN储量没有相关性。两个团聚体稳定性指数除微团聚体TN储量之外均与团聚体分布和团聚体氮储量相关,其中与微团聚体占比呈负相关。 图6 土壤全氮储量与团聚体分布和稳定性以及团聚体全氮储量关系Fig.6 Relationship between soil TN stock and distribution and stability of aggregates and TN stock of aggregatesSTNs:土壤全氮储量 soil total nitrogen stocks;*表示因子间显著相关(P<0.05),**表示因子间极显著相关(P<0.01) 表4和图7为团聚体的动态变化对于土壤全氮储量的贡献率,由表3可知,在草地恢复过程中,随年限增加土壤全氮储量变化量增加,恢复10-50a与农田相比变化量范围为0.003-0.045 kg/m2。其中大团聚体对土壤全氮变化的贡献率较大,而中团聚体贡献率较小,微团聚体对土壤全氮储量变化的贡献为负值,值得注意的是粘粉粒团聚体在恢复10a贡献值为负值,但随恢复年限增加呈现上升的趋势。在草地恢复过程中大团聚体对土壤全氮储量变化的贡献率最大,在10a时达到165.49%,而后接近稳定在70.16%-72.95%之间。此外,在草地恢复过程中大团聚体F1与农田对照相近而F2显著高于F1和农田,中团聚体F1在10a时低于农田,再随恢复年限增加呈现先上升后下降的趋势,而F2始终大于F1和农田。微团聚体F2显著低于农田,但在10a时高于F1,而后逐渐降低,F1呈现大于F2的趋势。 表4 不同恢复年限草地土壤氮储量以及团聚体氮储量对土壤氮储量变化的贡献率Table 4 Contribution of soil N stocks and aggregate N stocks to changes in soil N stocks in grasslands with different restoration ages 图7 不同恢复年限草地土壤全氮储量变化的两个驱动因子Fig.7 Two driving factors for changes in soil TN stocks in grasslands with different restoration agesF1表示由团聚体TN含量变化所驱动的土壤TN储量变化; F2表示由团聚体占比变化所驱动的土壤TN储量变化 植被恢复过程中,地上植物群落特征会随恢复年限发生改变[21]。本研究表明,草地恢复过程中,菊科、豆科和禾本科植物为研究区主要优势物种(表1),这与黄土高原植被分布特征相吻合[22]。这主要是由于独特的生物学特性使这三科植物对本研究区的生态环境有很强的适应性[23]。一方面菊科、禾本科和豆科植物通过增加根系生物量来吸收更多的水分[24],同时通过增加根冠比来减少水分散失(表1),另一方面在干旱胁迫下这三科植物通过渗透调节保持体内水分,并增加SOD和丙二醛含量来抵御干旱胁迫[25],并且豆科植物可与根瘤菌共生来增加耐旱性[26]。虽草地恢复过程中主要优势种为这三科,但不同植物在不同恢复阶段的重要值存在差异[10]。本研究结果显示,在草地恢复10-50a间,草地群落经历了茵陈蒿→茵陈蒿+铁杆蒿→铁杆蒿+兴安胡枝子→铁杆蒿+本氏针茅→铁杆蒿+白羊草的演替过程(表1),该结果与董凌勃等[10]在黄土高原的研究结果基本一致。这是由于各不同植物在受到光照、水分、土壤等环境因素以及种间和种内竞争作用影响[27],其中茵陈蒿在恢复前20a能促进铁杆蒿、胡枝子和白羊草幼芽和根系生长,但在恢复30a后铁杆蒿通过化感作用来抑制其他植物生长发芽[24]。 草地群落物种组成在恢复过程中的变化使不同恢复年限草地群落多样性发生变化,本研究中,随着恢复年限的增加草地群落物种数、Margalef丰富度指数、Shannon多样性指数和Pielou均匀度指数呈现出先增后减趋势(图2),乔文静等[22]在黄土高原研究也有相同的结果,这是由于在草地恢复10-30a间,草地群落的生物量增加(表1)、总蒸腾量增多,致使土壤含水量逐渐降低(表3),进而导致30a之后部分植物受水分限制而退出群落,导致物种多样性减少。此外,恢复30a后优势物种竞争、化感作用的增强,部分优势物种占据更广生态位使其他草本植物正常生长受到抑制[23]。草地群落演替和多样性的变化除了使物种变化的同时改变了植被养分含量。本研究结果显示,不同恢复年限草地的地上植被、根系和凋落物TN含量随恢复年限呈先增后减的趋势(表2),这是由于植被恢复前30年,土壤条件不断改善[28],豆科植物禾本科等植物的生物量不断增加,导致植被碳、氮等养分含量增加。但是,30a后菊科和禾本科植物占主导地位,豆科植物减少,其中菊科和禾本科含氮量较低,但作为C4植物的禾本科含碳量较高[29],进而导致植被、根系和凋落物碳含量增加而氮含量下降。 植被恢复过程中,地上植物和地下土壤的协同作用会改变土壤养分状况[11,28,30]。本研究结果显示,随草地恢复年限增加土壤PH和容重呈下降趋势,粘粒含量呈上升趋势(表3),表明植被恢复使土壤条件得到不断改善[31]。这是由于草地恢复过程中植被覆盖度和凋落物增加,导致土壤表层覆盖物增多,土壤水分蒸发减少,另外有机质的输入使土壤有机酸含量上升[32],进而降低土壤PH。植物根系生物量增多,导致土壤孔隙度增加,使土壤容重下降。而根系分泌物增加以及微生物分解增加了土壤有机质,再加上土壤保水性增强,导致土壤粘性增加,使土壤质量得到改善[11]。不同的是土壤含水量在恢复过程中先下降后上升(表3),这是由于恢复初期草地群落的地下生物量增加,根系吸水增加,而地上覆盖度增多导致了总蒸腾量增多,进而使土壤含水量逐渐降低,但30a之后草地物种多样性降低,耐旱的优势物种占据群落主导地位,水分蒸散减少,并且凋落物覆盖增加,土壤保水性增强,所以在草地恢复30-50a土壤含水量逐渐上升。草地恢复过程当中土壤有机碳含量增加(表3),这是由于植被恢复过程地上植被增多,其中含C量较高的禾本科植物增多可将含C营养物质归还与土壤[22],同时凋落物和根系生物量增加,以及它们分解速率加快[33],增加了土壤有机碳(OC)含量,同时充足的C资源使微生物生长和代谢需求得到了满足,增强了土壤固碳能力。而土壤全氮(TN)含量呈现先增加后降低的趋势(表3),在30a之前豆科植物变多,通过固氮作用和凋落而归还于土壤的N增多,进而增加土壤TN含量,但30a之后菊科和禾本科植物占群落主导地位,其中菊科植物含N量较低,加上豆科植物减少,归还于土壤的N下降,并且微生物活动的增加造成更多的N排放[34],导致土壤TN含量降低。 地上植物群落特征的结构组成和地下土壤理化性质的改变会影响土壤团聚体组成与分布[35]。本研究结果显示,随恢复年限的增加土壤大团聚体(>2mm)和中团聚体占比(0.25-2 mm)增加,而微团聚体占比(0.053-0.25 mm)显著减少(图3)。大团聚体和中团聚体形成与植被、根系和凋落物生物量和碳氮含量有关(图5),这是由于草地恢复过程中植被覆盖度、凋落物增加,减少了土壤水分蒸发散失并增加了腐殖质与腐殖酸[36],同时根系生物量增加,根系通过固结缠绕和释放分泌物,增强土壤抗侵蚀和土壤微小颗粒团聚胶结的能力,进而增加土壤大粒径团聚体的占比[37],李程程等[38]认为植被恢复过程中土壤质量的改善影响了团聚体组成,土壤当中水分以及碳氮含量的改变影响了土壤微生物和土壤酶活性,进而对小粒级进行聚集形成大粒级团聚体[1],这与本研究分析结果大团聚体和中团聚体形成与土壤有机碳、氮、碳氮比、含水量、粘粒和粉粒含量呈正相关(图5)一致。团聚体组成变化使团聚体稳定性发生变化,进而改变土壤稳定性[39]。本研究中,团聚体稳定性指数平均重量直径(MWD)和几何平均直径(GMD)均显著增加。这是由于土壤当中小的粘粉粒通过根系分泌物和微生物聚集,并向更大粒级团聚体转化[4,40-41],这表明团聚体稳定性在一方面与植被、根系和凋落物生物量和碳氮含量以及土壤有机碳、氮、碳氮比、含水量、粘粒和粉粒含量呈正相关(图5),另一方面与大团聚体、中团聚体和粘粉粒团聚体占比呈正相关性(图6)。 草地恢复过程中养分的输入与输出存在差异,进而对土壤TN在各粒级团聚体分布产生影响[42]。本研究结果显示,各粒级TN含量随恢复年限呈先下降后上升的趋势,而粒径小于2mm的团聚体TN含量又在40a后下降(图4)。这是由于草地恢复前10a,草地植物变多,土壤当中N被植物根系所吸收,因此各粒级团聚体TN含量减少,在10a之后,根系分泌物增加和凋落物在丰富的微生物作用下分解,同时各粒级当中微生物残体的增加[43],使各粒级TN含量上升,但在40a后,受到养分归还限制小于2mm粒径团聚体TN含量下降,而不同恢复年限草地各粒级全氮含量由高到低依次为:中团聚体>微团聚体>大团聚体>粘粉粒团聚体,这是由于中团聚体中有机物较多[2],且稳定性强,因此N持有能力较强,造成中团聚体当中TN含量较高。团聚体TN含量变化影响各粒级氮储量,本研究结果显示,粒径大于2mm的团聚体氮储量随恢复年限增加,而微团聚体氮储量呈先下降再上升后下降的趋势(图4)。这是由于恢复过程中的根系增多,土壤微小颗粒通过根系缠绕和分泌物聚集,形成稳定且具黏合特性的团聚结构[1],这保证了大粒径团聚体不被矿化或分解,且其黏性吸附更多的土壤颗粒,因此大粒径团聚体氮储量增加。微团聚体氮储量在恢复过程中先减少后增加,这是由于微团聚体氮储量与植被、根系和凋落物氮含量相关(图5),植被恢复使根系增多,根系更容易地从稳定性较低的团聚结构当中吸收含氮化合物[44],进而减少微团聚体氮储量,恢复10a后,豆科植物变多,氮归还增加,因此微团聚体氮储量增加,而微团聚体氮储量下降是因为30a后主要优势种菊科和禾本科本身含氮量较低,而豆科植物减少,并且微团聚体占比降低(图3),导致微团聚体氮储量减少。在农田以及草地恢复10-40a,微团聚体当中氮储量最高,但到50a大团聚体氮储量最高(图4),这是因为40a之前微团聚体占比虽有逐步降低但占比依然最大,到50a大团聚体占居主导地位(图3),这表明本研究符合TISDALL等[45]提出的团聚体的团聚体形成模型,随着团聚体粒径的增加,全氮的存储也会增加,因为粒径更大的团聚体是由小颗粒和有机结合剂组成的[46]。因此,大团聚体氮储量逐年增加。 不同恢复年限草地各粒级团聚体的氮储量对土壤氮储存有不同程度的贡献。本研究结果显示,土壤氮储量与大团聚体占比和氮储量呈现显著正相关(图6),恢复过程中土壤氮储量增加,且大团聚体对土壤氮储量的贡献率最大(表4),这是由于草地恢复过程中,含氮量较高的豆科植物变多,同时根系和凋落物的分解促进了土壤颗粒的黏合,因此大团聚体TN含量和数量增加,另外,恢复过程中人类活动的减少,以及土壤表面植被覆盖度和凋落物生物量增加减少了雨水的冲击,防止了土壤团聚结构被破坏,利于大团聚体的形成[47],进而增加了大团聚体氮储量。而大团聚体氮储量增多是由其本身占比和TN含量在恢复过程中的增加导致的,其中大团聚体占比的增加是大团聚体氮储量的增加主原因(图7)。这是由于植被恢复过程中大团聚体通过根系缠绕和有机物分解对土壤小颗粒黏合而成[1],并且在恢复过程中不断聚集,成为稳定性较强的结构,有效地避免了被微生物以及矿化过程所分解[48],因此微团聚体占比在不断减少,而大团聚体占比增加(图3),导致大团聚体氮储量在不断增加。总之,本研究中大团聚体占比的变化主导其氮储量的动态变化,而大团聚体氮储量是土壤氮储量变化的主要贡献粒级。 (1)黄土高原草地恢复过程中主要优势物种为菊科、禾本科和豆科植物,草地植物物种多样性随恢复年限的增加呈先增加后减少趋势,草地植物群落发生正向演替。 (2)草地恢复过程中土壤含水量、土壤OC和TN含量呈增加趋势,土壤pH和容重呈下降趋势,土壤质量得到有效改善。 (3)随恢复年限的增加,草地群落土壤大团聚体占比增加,微团聚体占比下降,土壤团聚体稳定性逐渐增强。 (4)草地群落土壤各粒级团聚体TN含量表现为中团聚体>微团聚体>大团聚体>粘粉粒团聚体,且均随随恢复年限的增加呈先降后升的趋势;TN储量表现为微团聚体>大团聚体>中团聚体>粘粉粒团聚体,且大、中团聚体均随恢复年限的增加呈增加趋势。 (5)草地恢复过程中土壤大团聚体对土壤TN储量的贡献最大,且其通过占比增加的方式促进土壤全氮储量增加。1.8 数据分析与作图
2 结果分析
2.1 不同恢复年限草地植物群落组成


2.2 不同恢复年限土壤理化性质特征

2.3 不同恢复年限草地团聚体粒级分布和稳定性特征

2.4 不同恢复年限草地团聚体氮含量与储量

2.5 植被特征和土壤理化性质与团聚体组分和TN储量关系

2.6 土壤TN储量与团聚体组分和TN储量的关系

2.7 土壤团聚体对土壤TN储量变化的贡献率


3 讨论
3.1 植被恢复对植物群落及其养分特征的影响
3.2 植被恢复对土壤理化性质及养分的影响
3.3 植被恢复对土壤各粒级团聚体组成与分布的影响
3.4 植被恢复对土壤各粒级团聚体全氮分布的影响
4 结论
