群体感应淬灭酶AiiO的克隆表达及其发酵工艺优化
2024-01-08田唱,郭婉萍,陈阳,赵晶,陈明
田 唱, 郭 婉 萍, 陈 阳, 赵 晶, 陈 明
( 1.大连工业大学 生物工程学院, 辽宁 大连 116034;2.大连民族大学 生命科学学院, 辽宁 大连 116600 )
0 引 言
酰基高丝氨酸内酯(N-acyl-homoserine lactones,AHLs)是革兰氏阴性菌群体感应系统(Quorum sensing,QS)中广泛存在的一种信号分子,当AHLs浓度超过一定阈值时,能够与相应的受体蛋白结合,进而调控多种生理功能,如毒力基因的表达、生物膜的形成等[1]。群体感应淬灭酶能够通过酶促反应降解信号分子AHLs,保持信号分子浓度低于阈值,进而有效干扰细菌的QS系统,抑制病原菌的致病性[2-3]。目前,已经有很多群体感应淬灭酶被分离鉴定,高丝氨酸内酯酶和酰化酶是群体感应淬灭酶中重要的两类。群体感应淬灭酶作为一种研究最多也是最有效的淬灭途径,在克隆表达[4]、分离纯化[5]、酶学性质[6]、作用机理[7]、抗菌活性[8]、工业应用[9]等方面研究成果丰富。AiiO酶于苍白杆菌Ochrobactrumsp. A44中被发现[10],属于AHL酰化酶中的α/β水解酶折叠酶系[4],能够降解酰基侧链长度为4~14的AHLs及其相应的3OH-Cx-HSLs[11]。国内外对AiiO酶的相关报道比较少,高效表达AHL酰化酶AiiO有利于后续的病原菌防控制剂开发及其在致病菌中的应用研究。目前,常规表达外源蛋白的方法是利用大肠杆菌表达系统和IPTG诱导的方式。但是,课题组前期采用这种方式诱导表达AiiO可溶性表达量很低,无法满足后续研究的需要。
乳糖是乳糖操纵子的天然诱导剂,相较于常用的IPTG诱导剂昂贵的价格和潜在的毒性,在工业化生产中更加具有应用前景。林晓栩等[12]利用乳糖诱导重组环糊精葡萄糖基转移酶(CGTase)表达,胞外酶活达19.87 U/mL。
本研究利用构建的重组工程菌E.coliBL21(DE3)-pET-28a(+)-aiiO进行乳糖自诱导发酵,并对该发酵过程中诱导温度、氮源浓度、乳糖添加时机和乳糖浓度等因素进行优化,以期提高目的蛋白AiiO的表达量。
1 材料与方法
1.1 菌种及质粒
小麦苍白杆菌(Ochrobactrumtritici)、质粒pET-28a,本实验室保存;大肠杆菌E.coliDH5α、BL21(DE3),北京全式金生物技术有限公司。
1.2 培养基
LB液体培养基(g/L):蛋白胨10,酵母提取物5,氯化钠10。
高密度增殖培养基(g/L):葡萄糖5,磷酸氢二钠3.55,磷酸二氢钾3.40,氯化铵2.68,硫酸钠0.71,七水合硫酸镁0.50,柠檬酸三钠0.30,六水合琥珀酸钠2.50、三氯化铁(0.1 mol/L) 1 mL。
复合自诱导培养基(g/L):蛋白胨(T)10,酵母粉(Y)5,甘油26,乳糖2,磷酸氢二钠3.56,磷酸二氢钾3.40,氯化铵2.68,硫酸钠0.71,七水合硫酸镁0.50,葡萄糖0.50,柠檬酸三钠0.30,六水合琥珀酸钠5.40,三氯化铁(0.1 mol/L) 1 mL。
1.3 重组表达载体pET-28a(+)-aiiO的构建
根据GeneBank中公布AiiO蛋白的基因碱基序列,以及载体pET-28a(+)中的多克隆酶切位点,利用NCBI设计上下游引物,引物序列分别为F:5′-GGGAATTCCATATGATGAAATCCC ATGAAAT-3′(下划线处为NdeI位点);R:5′-C GGAATTCTTAAGCCGTGCAGTC-3′(下划线处为EcoRI位点)。提取O.tritici基因组,以此为模板扩增aiiO(810 bp),采用酶切酶连的方法将扩增产物与pET-28a载体连接。将连接产物化学转化E.coliDH5α感受态细胞,挑取阳性重组子,对重组质粒pET-28a(+)-aiiO进行双酶切验证。选取验证正确的重组质粒送生工生物工程(上海)股份有限公司进行测序。
1.4 重组蛋白AiiO的诱导表达及发酵条件优化
将测序正确的重组质粒pET-28a(+)-aiiO转化到E.coliBL21(DE3),接种于高密度增殖培养基(含50 μg/mL卡那霉素)中,37 ℃、180 r/min过夜活化后按照0.1%接种量转接于复合自诱导培养基,37 ℃、250 r/min条件下培养。
采用单因素试验依次对发酵过程中诱导温度、氮源浓度、乳糖添加时机和乳糖浓度进行优化,比较发酵过程中甘油消耗、菌体干重、AiiO表达量,确定最优发酵条件。
1.5 重组蛋白AiiO的纯化
收集菌液,4 ℃、6 000 r/min离心15 min,将收集的菌体悬浮于Tris-HCl缓冲液中进行超声破碎,功率40%,工作2 s停3 s,共破碎40 s,再于4 ℃、13 000 r/min离心30 min,收集上清液。对收集的上清液进行亲和层析纯化、10 U/mg 凝血酶20 ℃水浴过夜酶切、凝胶过滤层析。
1.6 分析方法
1.6.1 甘油消耗量
取1 mL发酵液,6 000 r/min离心15 min,收集上清液,0.22 μm无菌滤膜过滤后的清液作为样品,选用北京普利莱基因技术有限公司的甘油测定试剂盒对甘油质量浓度进行测定。
1.6.2 菌体干重
取50 mL发酵液,4 ℃、6 000 r/min离心15 min,弃上清,沉淀用Tris-HCl缓冲液洗涤两次,80 ℃烘干至质量稳定,记录菌体干重。
1.6.3 目的蛋白AiiO含量
对细胞破碎液全蛋白和其离心后上清液进行SDS-PAGE检测。以已纯化且已知浓度的AiiO为标准上样,利用Quantity One软件对电泳图进行分析,计算目的蛋白含量。
2 结果与讨论
2.1 重组表达载体及工程菌的构建
将PCR扩增得到的目的基因aiiO(810 bp)切胶纯化后,用1 %琼脂糖凝胶电泳检测,结果如图1(a)所示。在750~1 000 bp存在单一条带,其大小与理论值相符,表明aiiO基因扩增成功。连接后的质粒经双酶切电泳验证,结果如图1(b)所示,在5 369和810 bp附近各有一条酶切条带,符合载体pET-28a(+)和目的基因aiiO的大小,初步判断为阳性克隆,送至生工生物工程(上海)股份有限公司测序。将测序正确的质粒导入大肠杆菌E.coliBL21(DE3)中,成功构建重组工程菌E.coliBL21(DE3)-pET-28a(+)-aiiO。
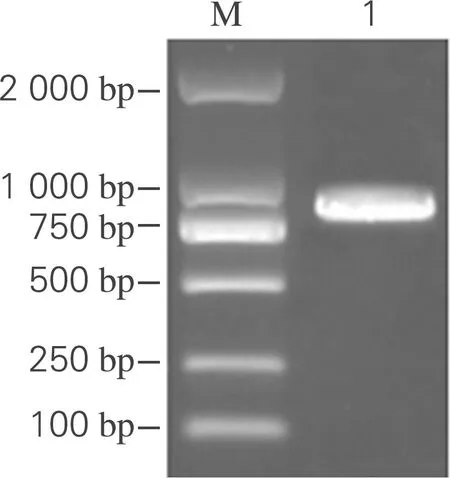
(a) PCR扩增目的基因
2.2 培养条件的优化
2.2.1 诱导温度
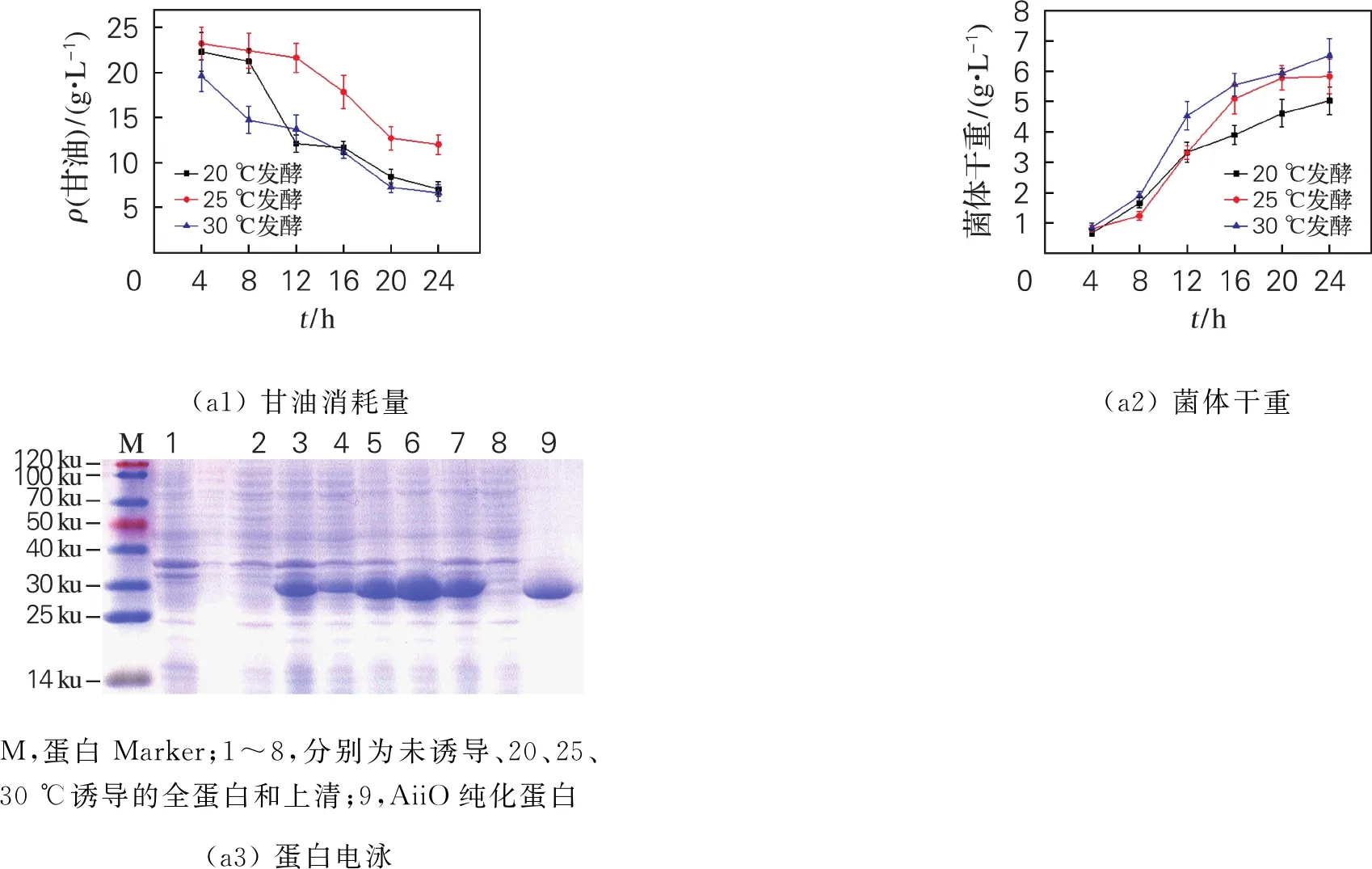
菌体转接至复合自诱导培养基,37 ℃培养2~3 h,OD600在0.6~0.8,分别于20、25、30 ℃进行诱导培养。由图2(a)可知,在诱导温度30 ℃时,0~8 h内甘油消耗较快,诱导结束时菌体干重最大。但在30 ℃条件下,胞内可溶性目的蛋白AiiO浓度极低,可能是因为菌体生长过快,导致包涵体的形成。20 ℃的低温诱导条件下虽然有利于肽链的正确折叠,但由于菌体生长缓慢,最终胞内可溶性AiiO浓度也较低。25 ℃培养菌体诱导产酶,胞内可溶性目的蛋白AiiO产量最高,因此选择25 ℃为最适诱导温度。
(a) 诱导温度
2.2.2 初始氮源质量浓度
利用不同质量浓度氮源(T 10 g/L+Y 5 g/L,T 15 g/L+Y 7.5 g/L,T 20 g/L+Y 10 g/L)的复合自诱导培养基,37 ℃培养菌体OD600为0.6~0.8,25 ℃进行诱导,至OD600趋于稳定。由图2(b)可以看出,在氮源质量浓度选择T 20 g/L+Y 10 g/L时,菌体干重高于其他,同时甘油消耗量也最大。由表1可知,随着氮源质量浓度的增大,胞内可溶蛋白AiiO产量随之增加,在T 20 g/L+Y 10 g/L条件下,胞内可溶性AiiO产量最高,达到276.04 mg/L。因此,选取T 20 g/L+Y 10 g/L为最适氮源质量浓度。
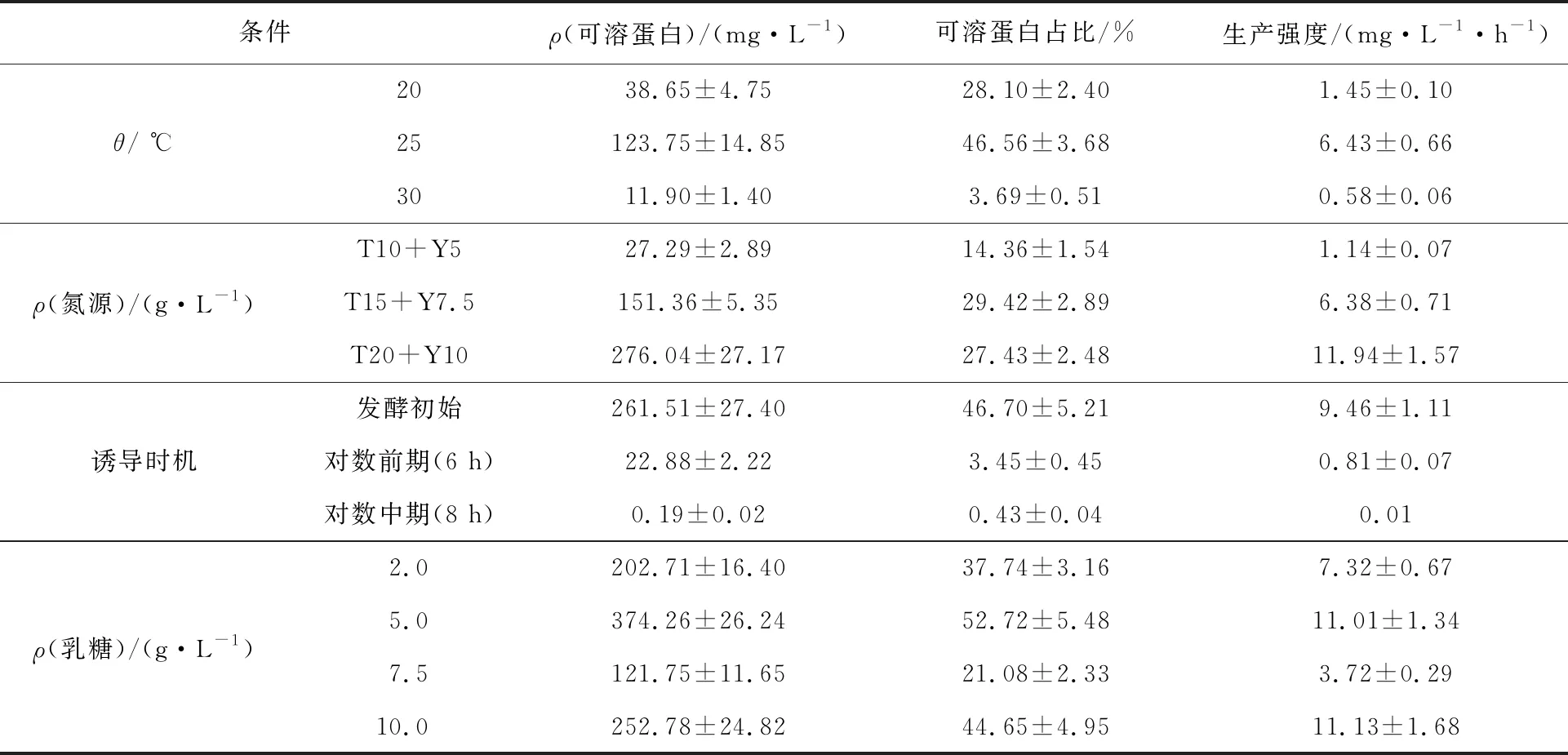
表1 不同条件下发酵产AiiO蛋白结果
2.2.3 添加乳糖诱导时机
选择T 20 g/L+Y 10 g/L复合自诱导培养基,37 ℃培养菌体OD600为0.6~0.8,25 ℃继续培养,分别在发酵初始、对数前期(6 h)、对数中期(8 h)添加2 g/L乳糖诱导产AiiO。由图2(c)和表1可知,在菌体生长的对数中期添加乳糖,基本不能起到诱导作用;在对数前期添加乳糖,虽然在细胞破碎后的全蛋白中检测到较高浓度的AiiO,但胞内可溶性AiiO浓度很低。在发酵初始添加乳糖,胞内可溶性AiiO产量最高,说明乳糖最佳添加时机为发酵初始时。
2.2.4 乳糖添加量
在氮源选择T 20 g/L+Y 10 g/L、发酵初始添加诱导物乳糖、诱导温度25 ℃的优化条件下,考察不同乳糖添加量(2.0、5.0、7.5、10.0 g/L)对重组菌株发酵产目的蛋白AiiO的影响,结果见图2(d)和表1。可以看出,不同的乳糖添加量对重组菌株E.coliBL21(DE3)-pET-28a(+)-aiiO甘油消耗和菌体细胞生长的影响较小,4种乳糖添加量下甘油消耗和菌体干重变化趋势基本一致,说明重组菌株优先利用复合自诱导培养基中甘油、蛋白胨和酵母粉等碳氮源,乳糖作为一种不易被利用的碳源,其添加量多少对甘油消耗及菌体细胞生长基本没有影响。由表1可见,4种乳糖添加量下,细胞内可溶性目的蛋白AiiO(上清)产量表现出较大差异,乳糖添加量为5 g/L时,胞内可溶性AiiO产量最高,达到374.26 mg/L,明显高于其他3种。因此,选取诱导物乳糖的适宜添加量为5 g/L。
3 结 论
以常规诱导剂IPTG诱导重组菌株E.coliBL21(DE3)-pET-28a(+)-aiiO产AiiO蛋白,可溶性表达量极低。采用乳糖代替 IPTG为诱导剂,从诱导温度、氮源浓度、乳糖添加时机和乳糖添加量等4个方面进行优化。结果发现,相较于IPTG,以乳糖为诱导剂显著提高了AiiO蛋白的可溶性表达,在诱导温度25 ℃、氮源为20 g/L蛋白胨和10 g/L酵母粉、发酵初始添加5 g/L乳糖条件下,胞内可溶性目的蛋白AiiO产量达到374.26 mg/L,占胞内总目的蛋白的52.72%。
