深圳双扇蕨依存环境的植物群落学特征
2024-01-02马钰琦李宇惠顾钰峰王发国
马钰琦 李宇惠 林 琳 沈 悦 顾钰峰 王发国*
(1.中国科学院华南植物园,广东省应用植物学重点实验室,广州 510605; 2.深圳市生态环境局大鹏管理局,深圳 518116; 3.深圳市兰科植物保护研究中心,深圳 518114)
深圳双扇蕨(Dipteris shenzhenensis)为双扇蕨科(Dipteridaceae)双扇蕨属(Dipteris)植物,是2020年于深圳市大鹏新区被发现的新物种[1]。该种形态上与中华双扇蕨(D.chinensis)相似,特征不同在于:根状茎密被开张的黑色长鳞片,叶柄为栗色,叶片背部颜色为灰白色,有2个基部具阔翅相连的扇形羽片。该种目前被国际自然保护联盟(IUCN)列为极度濒危(CR)物种,保护状况不容乐观,其状况包括:(1)仅在深圳市东部山地发现,种群面积小,个体数量不多;(2)陆栖于岩石,喜好酸性土壤,对生长环境要求高,人工栽培和繁殖都很困难;(3)生长在低海拔地区,易受到人为活动干扰;(4)关于该种研究少,自新种发现后再无其他研究报道。本研究通过探讨深圳双扇蕨依存的植物群落学特征,掌握该濒危种生存环境条件、所处群落稳定性,基于生态位分析、种间关系分析,直观反映该种在群落中的生态特点,为进一步调查该种分布,营造适合该种生存的植物群落以及提出合理保护措施提供理论和实践指导。
1 研究区概况
研究区位于深圳大鹏半岛杨梅坑,该地区处于七娘山与老虎坐山之间,距离海边较近,面临大亚湾,与惠州巽寮湾隔海相望。气候属亚热带季风气候,全年温暖,日照充足,降水充沛。年平均气温22 ℃,年日照时间2 011 h,日照率高达47%。年降雨量2 280 mm 左右,4—9 月有大量降水,属高温多雨地区[2]。土壤主要由赤红壤、红壤和冲击土组成,属于酸性土壤,土质黏重且含有机质不多[3]。植被类型属于亚热带常绿季雨林,同时有面积较大的亚热带常绿阔叶林,植被茂密且多样性丰富。
2 研究方法
1.1 样地调查
深圳双扇蕨群落学特征调查在研究区西南部沟谷中沿杨梅坑河进行,沿途共寻有深圳双扇蕨分布集中区域3 个,分别设20 m×20 m 样地。记录样地经纬度、海拔、坡向、坡度、郁闭度。将所有样地分割为4 个10 m×10 m 区域进行样方调查。记录样方内乔、灌木植物物种名称、胸径、高度、冠下高、冠幅等以及小灌木及草本植物物种名称、高度、盖度。各样地基本信息如表1 所示。
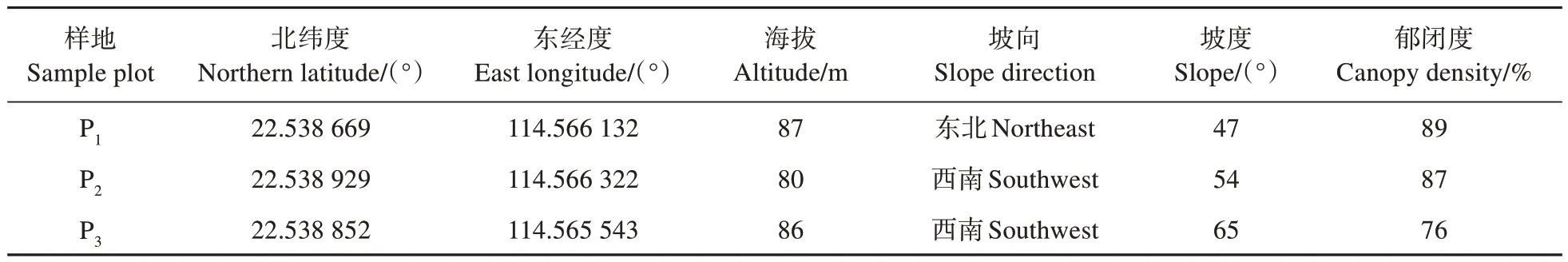
表1 杨梅坑河各样地基本信息Table 1 Basic information of test plot in Yangmeikeng river
1.2 数据处理
1.2.1 重要值
选用重要值表示物种的优势程度,计算方法如下[4-5]:
相对多度=(某一种植物的个体总数量/同一生活型植物个体总数量)×100% (1)
频度=(该种植物出现的样地数量/所调查的样地总数量)×100% (2)
相对频度=(一个种的频度/所有种的频度和)×100% (3)
相对显著度=(该种所有个体胸面积之和/所有种个体胸面积总和)×100% (4)
相对盖度=(每个物种的盖度/所有物种的盖度和)×100% (5)
乔木种的重要值=(相对多度+相对频度+相对显著度)÷3 (6)
灌木(草本)种的重要值=(相对多度+相对频度+相对盖度)÷3 (7)
1.2.2 多样性指数
比较Margalef 丰富度指数(E)[6]、Simpson 多样性指数(D)[7]、Shannon-Wiener 多样性指数(H)[8]、Pielou 均匀度指数(J)[9]研究不同群落间结构及物种多样性差异,计算公式如下:
式中:S为样方内的植物种类总和;N为样方内所有植物的个体数量总和;Pi为种i的个体占所有种的个体数量的比例。
1.2.3 年龄结构
依据优势种胸径划分不同立木级,进而对群落年龄结构进行分析,立木级划分标准如下[10]:Ⅰ级小树,胸径<2.5 cm;Ⅱ级壮树,2.5 cm≤胸径<7.5 cm;Ⅲ级大树,7.5 cm≤胸径<22.5 cm;Ⅳ级老树,胸径≥22.5 cm。
1.2.4 生态位
生态位反映物种在群落中的地位和状态,是分析物种间生存机制、竞争关系和群落动态演替等方面的重要理论[11]。本研究采用生态位宽度、生态位重叠度和生态位相似度描述生态位特征。生态位宽度指物种利用各种资源总和的幅度。采用Colwell加权修正的Levins指数(BCL,i)[12]表示,公式为:
式中:Pij为种i在资源位j的重要值占其在所有资源位置重要值总和的比例;r为样方数量。
生态位重叠度采用Pianka 指数(OP,ih)[13],公式为:
式中:Phj为种h在资源位j的重要值占其所在所有资源重要位总和的比例。
生态位相似度采用Schoener 指数(OS,ih)[14],公式为:
1.2.5 种间联结性
种间联结性用来描述不同物种在空间分布上的相互作用关系,可以用来分析群落发育过程中种间相互关系及表征优势种分布,进一步研究群落结构、功能及其演替趋势[15]。采用AC 联结系数(AC)[13]来衡量种间联结性,公式如下:
式中:a为植物A 与植物B 同时出现的样方数量,b为只有植物B 出现的样方数量,c为只有植物A 出现的样方数量,d为植物A 与植物B 都不出现的样方数量。AC在[-1,1]范围内有效,AC=0,种间完全独立,AC越趋向1,种间正联结程度越高,AC越趋向-1,种间负联结性越强。
1.3 数据分析
所有数据分析过程在R4.2.2 软件中完成,通过vegan 包进行多样性数值的计算[16],通过spaa 包进行生态位与种间联结性有关计算,然后利用ggplot2包实现数据可视化。
2 结果与分析
2.1 深圳双扇蕨群落组成分析
调查表明,深圳双扇蕨占优势的群落类型是该地区典型的亚热带常绿阔叶林,根据群落优势种组成,命名为:大头茶(Gordonia axillaris)+谷木叶冬青(Ilex memecylifolia)+吊钟花(Enkianthus quinqueflorus)+豺皮樟(Litsea rotundifoliavar.oblongifolia)+深圳双扇蕨+芒萁(Dicranopteris pedata)群落,根据研究对象以及群落特征种为深圳双扇蕨,将该群落称作深圳双扇蕨群落。群落共有维管植物49科73属85种,其中石松类和蕨类植物7 科9 属12 种,裸子植物2 科2 属2 种,被子植物40科62属71种。群落主要组成:茜草科(Rubiaceae)7 种,冬青科(Aquifoliaceae)5 种,樟科(Lauraceae)4 种,大戟科(Euphorbiaceae)3 种,桃金娘科(Myrtaceae)3 种,豆科(Fabaceae)3 种,山茶科(Theaceae)3 种,莎草科(Cyperaceae)3 种,里白科(Gleicheniaceae)3 种,鳞 始 蕨 科(Lindsaeaceae)3 种,夹竹桃科(Apocynaceae)3 种。该群落物种构成具有明显亚热带地区性质,符合该地区热带与亚热带过渡的特点。
植物生活型指植物对特定生境长期适应而在外貌反映出来的类型,可反映群落垂直结构与外貌特征。生活型按垂直结构可分为乔木层、灌木层、草本层3 个层次。深圳双扇蕨群落乔木层共29 种植物,以大头茶为优势种,谷木叶冬青与竹叶青冈(Cyclobalanopsis neglecta)次之。乔木层可分为2个亚层。第一亚层高度10~14 m,以上述3 种植物为主,第二亚层在10 m 以下,以大头茶、软荚红豆(Ormosia semicastrata)及谷木叶冬青为主,其他有鼠刺(Itea chinensis)、降真香(Acronychia pedunculata)、岭南山竹子(Garcinia oblongifolia)等。灌木层共有植物59种,以吊钟花和豺皮樟为优势种,其他有鼠刺、降真香、狗骨柴(Diplospora dubia)、桃金娘(Rhodomyrtus tomentosa)、粗叶木(Lasianthus chinensis)及上层乔木的幼树,如:谷木叶冬青、大头茶等。草本层植物约38 种,以深圳双扇蕨、芒萁为主,其他有鳞籽莎(Lepidosperma chinense)、乌毛蕨(Blechnum orientale)、粤里白(Diplopterygium cantonense)等,层间植物以藤本植物为主,如土茯苓(Smilax glabra)、罗浮买麻藤(Gnetum luofuense)等。
对深圳双扇蕨群落中植被高度与个体数量关系进行分析,采取分类的方法[17],标准为:A 类,树高<4 m;B类,4 m≤树高<8 m;C类,8 m≤树高<12 m;D类,12 m≤树高<16 m;E类,16 m≤树高<20 m。结果表明个体数量随植被高度变化呈现“倒J”型分布(见图1),即随植被高度增加,植株个体逐渐减少。“倒J”型分布特征是热带和亚热带地区植物群落中常见的分布[18-19],符合当地地理特点,同时该分布表明群落中幼树、小树多,有良好的更新能力,说明群落的生态演替阶段可能位于先锋期或稳定期,排除了衰老期的可能。

图1 深圳双扇蕨群落树高与个体数量关系A.未分类;B.已分类。Fig.1 Relationship between tree height and individual number of D. shenzhenensis community A.Unclassified;B.Classified.
2.2 深圳双扇蕨群落重要值分析
2.2.1 乔木层物种重要值分析
由表2可知,乔木层有植物21种,16种植物重要值大于1,按从大到小排序依次为大头茶、谷木叶冬青、竹叶青冈、鼠刺、软荚红豆、豺皮樟、岭南山竹子、粘木(Ixonanthes chinensis)、亮叶柃(Eurya nitida)、五列木(Pentaphylax euryoides)、吊钟花、光叶山矾(Symplocos lancifolia)、密花树(Myrsine seguinii)、降真香、酸味子(Antidesma japonicum)、岗松(Baeckea frutescens),其中大头茶重要值为24.754 0,明显高于其他物种,占据乔木层优势。谷木叶冬青与竹叶青冈占次优势,二者重要值分别为14.050 8和13.872 0。

表2 深圳双扇蕨群落乔木层物种特征值Table 2 Characteristics values of the species in the arbor layer of D. shenzhenensis community
2.2.2 灌木层物种重要值分析
由表3 可知,灌木层共有植物49 种,种类构成较为复杂,谷木叶冬青及大头茶为上层乔木植物小苗,二者重要值约占整个灌木层总重要值18%。其次灌木植物吊钟花与豺皮樟的重要值较高,分别为5.941 3与5.675 2。

表3 深圳双扇蕨群落灌木层物种特征值Table 3 Characteristics values of the species in the shrub layer of D. shenzhenensis community
2.2.3 草本层物种重要值分析
由表4 可知,草本层植物共34 种,构成比乔木层复杂,比灌木层简单。重要值大于1 的有14 种,深圳双扇蕨与芒萁重要值为18.639 5 与18.320 5,占绝对优势,其次为鳞籽莎、乌毛蕨、粤里白、土茯苓与罗浮买麻藤。以上7 种植物重要值占草本层总重要值的64%,是草本层的主要组成部分。

表4 深圳双扇蕨群落草本层物种特征值Table 4 Characteristics values of the species in the herb layer of D. shenzhenensis community
2.3 深圳双扇蕨群落物种多样性分析
深圳双扇蕨群落丰富度指数、多样性指数以及均匀度指数均在生活型上呈现“中间高两头低”态势。丰富度由高到低依次为:灌木层、草本层、乔木层。多样性指数由高到低依次为:灌木层、草本层、乔木层。均匀度指数由高到低依次为:灌木层、乔木层、草本层。
将深圳双扇蕨群落各生活型物种指数与杨梅坑山地植物群落动态演替研究中血桐(Macaranga tanarius)-九节(Psychotria asiatica)-半边旗(Pteris semipinnata)群落(血桐群落)和大头茶-栀子(Gardenia jasminoides)-团叶陵齿蕨(Lindsaea orbiculata)群落(团叶陵齿蕨群落)各指数[20]比较(见表5)。发现:(1)深圳双扇蕨群落整体Margalef 丰富度指数小于血桐群落与团叶陵齿蕨群落,表明前者物种丰富度水平比后者低。(2)群落间Simpson多样性指数与Shannon-Wiener 多样性指数通过t检验进行相似性判断[21]。对数据样本进行服从高斯分布前置性检验,结果P均小于0.05,可以进行t检验。t检验结果显示在显著水平α=0.05 条件下各组间物种多样性差异不显著(见图2)。(3)Pielou均匀度指数深圳双扇蕨群落各生活型均高于其他2个群落,但该群落各生活型之间均匀度数值差距大导致总体值小于其他2个群落,说明在深圳双扇蕨群落中物种的空间分布水平并不一致,水平方向分布较垂直方向分布更为均匀。

图2 杨梅坑不同群落间多样性系数t检验Fig.2 t-test of diversity coefficient among different communities in Yangmeikeng

表5 杨梅坑部分群落内各生活型生物多样性指数Table 5 Biodiversity index of various life types in some communities in Yangmeikeng
2.4 深圳双扇蕨群落年龄结构分析
年龄结构分析选取深圳双扇蕨群落优势物种大头茶、谷木叶冬青、竹叶青冈为代表(见图3)。群落年龄结构特征表现为:占优势的乔木树种没有Ⅳ级老树,大头茶以Ⅱ级壮树和Ⅲ级大树为主,Ⅰ级小树也有,说明在演替中Ⅱ级壮树和Ⅲ级大树可以得到补充,处于稳定阶段。谷木叶冬青以Ⅱ级壮树为主,Ⅲ级大树与Ⅰ级小树少,在演替中,该种群未来发展潜力小,树种补充力弱,有衰退趋势。竹叶青冈Ⅱ级壮树和Ⅲ级大树与Ⅰ级小树较少,在群落演替中状态良好,属于稳定状态。总体来说,该群落以Ⅱ级壮树和Ⅲ级大树为主,处于稳定演替状态,演替方向正向。

图3 深圳双扇蕨群落优势乔木年龄结果分析Fig.3 Age analysis of dominant trees in D. shenzhenensis community
2.5 深圳双扇蕨群落草本层生态位分析
2.5.1 生态位宽度分析
生态位宽度取决于生态环境的异质性以及植物对环境的适应和利用力。生态位宽度越大,植物对环境的适应性越强,生态位宽越小,则对生境要求越高。按生态位宽度大小可分为广生态位(BCL,i>0.6)、中生态位(0.6≥BCL,i>0.2)和窄生态位(BCL,i≥0.2)3种类型[22]。
选取深圳双扇蕨群落草本层重要值大于1 的11种优势草本植物进行生态位宽度计算(见表6)。从大到小排序依次为芒萁(0.757 6)、乌毛蕨(0.276 8)、扇叶铁线蕨(0.221 1)、土茯苓(0.213 0)、黑莎草(0.212 1)、深圳双扇蕨(0.203 4)、团叶陵齿蕨(0.197 2)、鳞籽莎(0.142 0)、铁芒萁(0.108 6)、石松(0.106 0)、粤里白(0.104 8)。其中芒萁生态位宽度最高,为广生态位植物,对调查环境有很强的适应性,在群落竞争中占有显著优势,在群落演替中不会轻易被其他植物取代。乌毛蕨、扇叶铁线蕨、土茯苓、黑莎草、深圳双扇蕨占有中生态位,环境适应能力适中,对资源的竞争能力远不如芒萁,但高于其他6 种窄生态位草本植物,在群落演替中存在被取代的风险。
2.5.2 生态位重叠度与相似度分析
生态位重叠度表示种对于同一生态资源的竞争力,重叠度低,表示2 种植物对同一生态资源竞争关系小,反之则对同种生态资源竞争关系强。生态相似度表示种对对同一环境的适应性,相似度越高表示2种植物趋同适应性越强,反之则趋异适应性强。
深圳双扇蕨群落草本层55个种对生态位重叠度与相似度见表7。结果说明:所有种对生态位重叠度平均值为0.187。重叠度在[0,0.100)有29对,占总对数52.7%;在[0.100,0.500)有20 种,占总对数36.3%;在[0.500,1.000)有6 种,占总对数10.9%。重叠度由大到小排名前3 的种对为石松-鳞籽莎(0.977)、团叶陵齿蕨-乌毛蕨(0.880)、团叶陵齿蕨-粤里白(0.868),这些种对对周围环境需求相似,竞争激烈。39 个种对重叠度不为0,占总对数70.9%,说明群落优势草本间大部分都存有重叠现象。深圳双扇蕨与其他10 种优势草本均有重叠,与芒萁(0.410)重叠度最强,竞争关系最显著。次为扇叶铁线蕨(0.210)、乌毛蕨(0.183)、黑莎草(0.115),有较明显的竞争关系,但远不如与芒萁对环境资源的竞争激烈。与其他6 种优势草本重叠度不足0.1但大于0,说明种对间竞争关系弱,但仍存有对资源的少量竞争。所有种对生态位相似度平均值为0.154。相似度在[0,0.100)有28 种,占总对数50.9%;在[0.100,0.500)有23 种,占总对数41.8%;在[0.500,1.000)有4 种,占总对数7.2%。相似度由大到小排名前3 的种对为石松-鳞籽莎(0.842)、团叶陵齿蕨-乌毛蕨(0.663)、团叶陵齿蕨-粤里白(0.634),这些种对对环境有很强的趋同适应性。深圳双扇蕨与芒萁(0.232)、乌毛蕨(0.171)、扇叶铁线蕨(0.167)、黑莎草(0.143)均有趋同适应态势,但未表现出很强的趋同性。与其他6种优势草本优势度不足0.1,表现出趋异适应。

表7 深圳双扇蕨群落优势草本种对生态位重叠度与相似度Table 7 Niche overlap and similarity of dominant herbaceous species of D. shenzhenensis community
2.5.3 种间联结性分析
深圳双扇蕨群落草本植物AC 联结系数计算结果显示(见图4):正联结对数32 对,8 对正联结系数较高(r≥0.67),占总对数14.5%;9 对正联结系数一般(0.67>r≥0.33),占总对数16.4%;15 对正联结系数趋向独立(0.33>r≥0),占总对数27.2%,负联结对数23 对,4 对负联结系数程度趋向对立(0>r>-0.33),占总对数7.2%;2 对负联结系数一般(-0.33>r≥-0.67),占总对数3.6%;17 对负联结系数很强(-0.67>r),占总对数30.9%。深圳双扇蕨与鳞籽莎、石松表现强的正种间联结关系,与乌毛蕨、粤里白、扇叶铁线蕨表现出一般正联结关系,与团叶陵齿蕨表现出趋向独立的正联结关系,与土茯苓、黑莎草、铁芒萁表现出趋向独立的负联结关系,与芒萁表现出一般的负联结关系。

图4 深圳双扇蕨群落优势草本植物种间联结性Fig.4 Interspecies association of dominant herbaceous plants in D. shenzhenensis community
3 讨论
3.1 深圳双扇蕨群落演替水平
通过分析,判断深圳双扇蕨群落处于自然演替的稳定阶段,演替方向正向发展,理由如下:(1)重要值分析中大头茶、谷木叶冬青与竹叶青冈在乔木层中总占52%,在灌木层中总占21%,都有较高占比,且重要值皆远超其他物种,占有绝对的优势地位,很难在短时间经过自然演替被其他物种所取代。(2)将深圳双扇蕨与同地区的血桐群落和团叶陵齿蕨群落进行多样性指数的相似性分析,发现3个群落生物多样性水平没有显著差异,群落结构复杂,组织水平高,处于演替的稳定阶段,演替方向向正向发展。(3)该群落占据绝对优势的物种大头茶年龄结构以Ⅱ级壮树与Ⅲ级大树为主,Ⅰ级小树数量也较多,为后续壮树与大树的更新提供了可能性,大头茶为先锋树种,适应力强,耐旱耐寒,若非自然灾害或人为干扰,该种会保持一个发展的状态,优势地位难以撼动;谷木叶冬青Ⅰ级小树少,后续更新能力差,但Ⅱ级壮树数量较多,在较短时间的演替过程中Ⅱ级壮树会优先发展为Ⅲ级、Ⅳ级树木,可以保持现有优势地位,但超过大头茶的可能性不大。由于更新能力差,长远来看该物种的优势地位可能会由其他物种所取代;竹叶青冈Ⅰ级小树多,Ⅱ级壮树和Ⅲ级大树少,说明处于发展状态,具备成为该群落优势种的潜力。该群落没有Ⅳ级老树,在短时间内如果无自然灾害、人为干扰则优势物种不会轻易发生改变,演替处于相对稳定阶段。
3.2 深圳双扇蕨在群落中的生存状态
生态位与种间联结相关分析表明:(1)深圳双扇蕨在所调查群落草本层中重要值最高,占有优势地位,直观原因是其作为本次调查的研究物种,数量多、相对多度大以及其本身为大型蕨类,相对盖度大。作为对环境要求较高的珍稀濒危物种,该种并未出现在每个样方中,导致其相对频率较小;(2)深圳双扇蕨在优势草本植物中生态位宽度处于中生态位与窄生态位的交界,说明该植物对周围环境有要求,对该生境空间内环境资源利用能力不强,虽然该种在草本层重要值为最高但并不能称为广布种,在群落演替过程中存在被取代的风险;(3)生态重叠度与相似度分析表明,深圳双扇蕨与其他优势草本均有资源竞争现象,与芒萁环境资源竞争最强,与乌毛蕨、扇叶铁线蕨、黑莎草竞争次之,同时均与上述物种表现趋同适应,适应水平相近且均不强。当群落演替中环境资源充足时,深圳双扇蕨上述物种对环境表现趋同适应,但当环境资源随时间和空间变化而变得匮乏,其与上述植物表现出对资源的竞争,结合生态位宽度来看,该种在与上述物种竞争时并无优势,有被淘汰的风险;(4)深圳双扇蕨与鳞籽莎、石松、乌毛蕨、粤里白、扇叶铁线蕨、团叶陵齿蕨间存有正联结,说明与上述植物间存在依赖,在异质环境内几个物种有相似适应和反应,一种物种的存在对另一种物种可能是有利的,物种间可能存在互惠共生或资源互补。深圳双扇蕨与土茯苓、黑莎草、铁芒萁、芒萁存有负联结,说明在竞争资源中互斥,环境需求不相似,种间可能存在竞争。总体而言,深圳双扇蕨在群落中生存状态虽占有优势,但不能被称为广布种,该种对环境要求苛刻,与其他优势草本存在竞争,且竞争力不强,正联结植物数量少,重要值低,生态位宽度低,互惠共生及资源互补能力低。以上因素使得该濒危种在群落演替中有被淘汰的风险,需要采取保护手段。
4 保护建议
通过对深圳双扇蕨群落进行结构特征分析,发现该群落处于演替稳定阶段,具有较高的物种多样性和复杂的结构,演替向正向发展。这从一方面说明良好稳定的生态群落环境以及群落复杂的结构为深圳双扇蕨提供了生存条件;另一方面,也说明深圳双扇蕨对自然生长环境要求高。该种作为深圳东部大鹏半岛特有的濒危蕨类,其野外分布数量很少,均在坡度较陡的岩壁上生长,种群面积不大,生长地所处海拔均在100 m 以下,容易受到人为干扰。在调查地区的沟谷中,受到人为改造的地区均未发现该种的分布。通过对深圳双扇蕨的生存状态分析,发现该种对环境要求高,适应力弱,在群落中竞争力弱,演替中有被淘汰的风险,亟待保护。同时发现与该种可能有互惠共生及资源互补能力的植物鳞籽莎、石松、乌毛蕨、粤里白、扇叶铁线蕨、团叶陵齿蕨,在保护时可以提高资源在上述植物中的分配来间接作用于深圳双扇蕨对环境资源的利用。对于土茯苓、黑莎草、铁芒萁、芒萁等竞争物种,在资源充沛的条件下,竞争关系可以给深圳双扇蕨带来更好的发展空间,但在资源匮乏状态下则会加速该种的灭绝。通过分株繁殖的方式对深圳双扇蕨进行了繁育与迁地保护,但结果并不理想,成活率不高,后续考虑采用孢子繁殖的方式进行繁殖。综上,本研究建议采取就地保护的方法对该物种进行保护,在该种分布较密集的区域采取封山造林的措施,减少人为对该地环境的破坏,保护该地生物多样性,同时积极开展该物种的相关调查研究以及保育研究,抢救性地保护各种原生特有种及其原生地,在保证环境资源充足的条件下,人为干预地调配环境资源分配,营造适合该种生存状态。建立种质资源基因库,结合当地环境和科学研究探索最适合该种的珍稀濒危植物保护方法。
