三江源区鱼类多样性调查及保护对策
2024-01-01陈锋袁婷熊满堂赵先富马沛明朱滨张志永刘晖董方勇李键庸



摘要:了解三江源区鱼类多样性及其资源现状,为三江源区生态系统保护和流域管理提供科学依据。2022年8月开展了长江源、黄河源、澜沧江源鱼类调查,分析了3个水系鱼类组成与分布,探讨了鱼类多样性特征,研究了三江源地区鱼类群落的空间格局。结果表明:三江源区共调查到鱼类18种,其中长江源9种,黄河源8种,澜沧江源6种;根据相对重要性指数,长江源优势种3种,依次为细尾高原鳅(Triplophysa stenura)、小头高原鱼(Herzensteinin microcephalus)、裸腹叶须鱼(Ptychobarbus dipogon),黄河源优势种2种,包括麻尔柯高原鳅(T. markehenensis)和黄河裸裂尻鱼(Schizopygopsis pylzovi),澜沧江源优势种4种,依次为前腹裸裂尻鱼(S. anteroventris)、裸腹叶须鱼、东方高原鳅(T. orientalis)和细尾高原鳅;三江源区鱼类多样性相对较低,其中澜沧江源最高,长江源次之,黄河源最低。对鱼类种类组成进行聚类分析,结果表明长江源和澜沧江源相近,黄河源则与长江源、澜沧江源差距较大。为切实保护好三江源区鱼类资源,建议加强本底调查,开展关键栖息地生态环境监测和保护,构建三江源区水生生物资源数据库,关注气候变化对水生态环境的影响,预防和控制外来鱼类入侵。
关键词:鱼类多样性;空间格局;优势种;保护对策;三江源区
中图分类号:S931.1" " " " 文献标志码:A" " " " 文章编号:1674-3075(2024)05-0001-08
三江源区位于我国西部、青藏高原腹地、青海省南部,是长江、黄河和澜沧江的源头汇水区,地理坐标为北纬31°39′~36°12′,东经89°45′~102°23′,总面积36.3万km2,平均海拔3 600 m以上,大部分地区海拔4 000 m以上,有些山峰海拔甚至高于6 000 m(青海省环境保护厅,2017)。三江源地区以其独特的地理位置和海拔高度,拦截了西南气流带来的大量水汽而形成充沛降水,为长江、黄河、澜沧江的发源分别提供了总水量的25%、49%和15%(陈孝全和苟新京,2002;青海省环境保护厅,2017),被称为“中华水塔”。三江源是世界上高原生物多样性最集中的地区之一,是我国最重要、影响范围最大的生态调节区,也是全国生态系统最脆弱的地区,在全国生态文明建设和保护制度改革研究中具有极其重要地位(李志强等,2013;Tao et al,2018)。
三江源地处高海拔地区,地域辽阔,境内河流180余条,湖泊1 800多个,丰富的水体资源和复杂的水文、气候条件为水生生物提供了赖以生存的自然条件,源区内水生生物资源丰富,是长江、黄河、澜沧江上游珍稀濒危水生生物的天然集中分布区域,具有很高的经济、科研和文化价值(李志强等,2013)。目前,关于三江源地区的研究主要集中在气候变化(李林等,2006;Liu et al,2011)、流域生态要素结构与空间分布演化(Li et al,2014;李冠稳等,2021)、生态功能价值(刘敏超等,2005;郑德凤等,2020)、生态恶化与保护等方面(郭忠胜等,2009;陈兴和余正勇,2022),对水生生物研究相对较少。三江源地区鱼类多为高寒地带种类,生长速度慢,繁殖力低,种群再生能力相对弱,受气候变化和人类活动影响较大,鱼类资源总体呈现出下降的趋势,部分种类被列为濒危物种。鱼类是三江源食物网中重要组成部分,对生态系统健康和稳定起着关键作用,鱼类资源的减少会破坏食物网的结构,进而威胁生态系统稳定性。然而,关于三江源地区高寒鱼类群落研究不多,相关研究基础相对薄弱,针对整个三江源区鱼类资源的现状报道更是少见,研究主要聚焦在分类学(武云飞和吴翠珍,1992;李柯懋等,2009)、区划(武云飞等,1994)和流域内部分区域(长江源、黄河源和澜沧江源)的鱼类群落专题调查(唐文家等,2006;陈锋等,2012;李柯懋等,2012;唐文家等,2012;熊芳园等,2022)等方面。因此,有必要对三江源区水域生态系统开展基础调查研究工作。
2022年8月,笔者对三江源区长江、黄河、澜沧江3个水系的鱼类资源进行了初步调查,分析了该区域不同水系鱼类组成与分布情况,探讨了鱼类多样性特征,通过积累一手鱼类群落数据,为后续深入研究提供基础,进而为三江源地区生态系统保护和流域管理提供科学依据。
1" "材料与方法
1.1" "样点设置
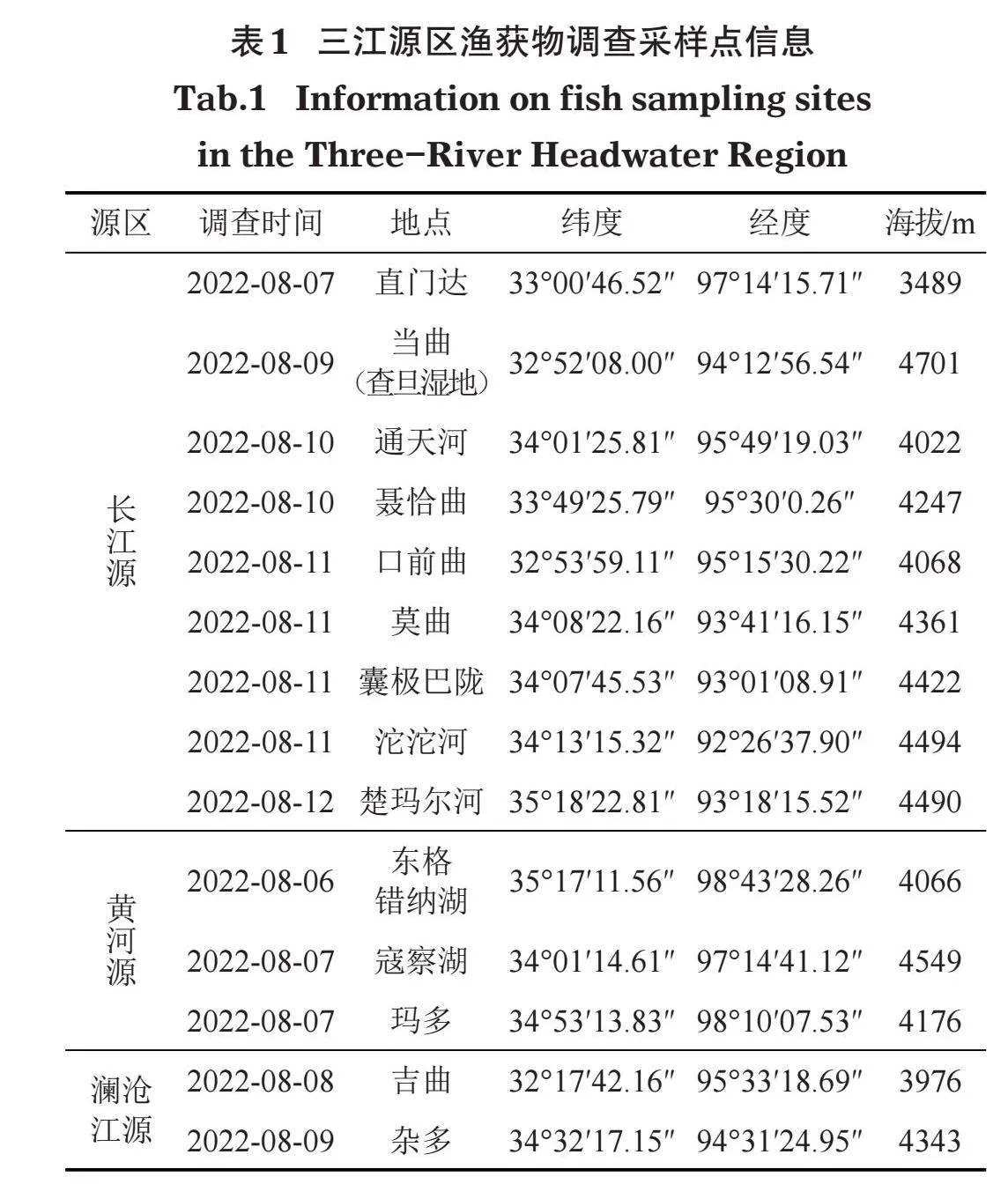
2022年8月,对长江源、黄河源、澜沧江源进行了考察,通过实地采样,分析三江源地区鱼类群落的空间格局。其中长江源采样点9个,均为河流生境;黄河源3个,2个为高原湖泊生境,1个为河流生境;澜沧江源2个,均为河流生境(表1)。
1.2" "样品采集
参照《内陆水域渔业自然资源调查手册》(张觉民和何志辉,1991),采用抄网、刺网、地笼等渔具采集长江源、黄河源、澜沧江源鱼类样品。其中,抄网网头直径40 cm,网目0.5 cm;刺网网目2~5 cm,网高1.5 m,长10 m;地笼长8 m,宽35 cm,高35 cm。参考《西藏鱼类及其资源》(西藏自治区水产局,1995)、《青藏高原鱼类》(武云飞和吴翠珍,1992)、《中国淡水鱼类检索》(朱松泉,1995)、《中国动物志硬骨鱼纲鲤形目(中卷)》(陈宜瑜,1998)、《中国动物志硬骨鱼纲鲤形目(下卷)》(乐佩琦,2000)对采集的渔获物样品鉴定到种,并测量体长(0.1 cm)、体重(0.1 g)等指标。
1.3" "数据处理
运用R 4.1.0软件eulerr包绘制3个水系鱼类组成韦恩图,运用层次聚类(聚合算法为非加权组平均法,UPGMA)对鱼类组成空间分布进行分析,基于Bray-curtis相异系数,使用R 4.1.0软件stats包中的hclust()实现,距离指数基于vegan包中的vegdist()计算而来。
利用Pinkas相对重要性指数(index of relative importance,IRI)来评估鱼类在各水域生态环境中的优势程度(Pinkas,1971),采用Shannon-Wiener指数(Wilhm,1968)、Pielou指数(Pielou,1975)和Margalef种类丰富度指数(Margalef,1957)研究鱼类群落多样性,计算公式为:
IRI = (N + B)×F×10000" " " " " " " " " " ①
式中:N为某一种类的数量占总数量的百分比,也称丰度占比;B为某一种类的重量占总重量的百分比,也称生物量占比;F为某一种类在采样中出现的次数占总捕捞次数的百分比。相对重要性指数IRI≥1 000的种类为优势种,100≤IRIlt;1 000的种类为常见种,10≤IRIlt;100的种类为一般种,IRIlt;10的种类为偶见种。
Shannon-Wiener多样性指数H′:
H′ = –Σ(Ni/N)ln(Ni/N)" " " " " " " "②
Pielou均匀度指数J′:
J′ = H′/lnS" " " " " " " " " " " " " "③
Margalef物种丰富度指数D:
D = (S–1)/lnN" " " " " " " " " " "④
式中:S为种类数,单位为种;Ni为种类i的个体数,N为渔获物总个体数,单位为尾。
2" "结果与分析
2.1" "鱼类种类组成与分布
本研究在三江源区共采集到鱼类609尾,隶属1目2科2亚科6属18种(表2)。渔获物均属于鲤形目,其中鲤科裂腹鱼亚科5属7种,占总种类数的38.89%,鳅科条鳅亚科1属11种,占总种类数的61.11%。
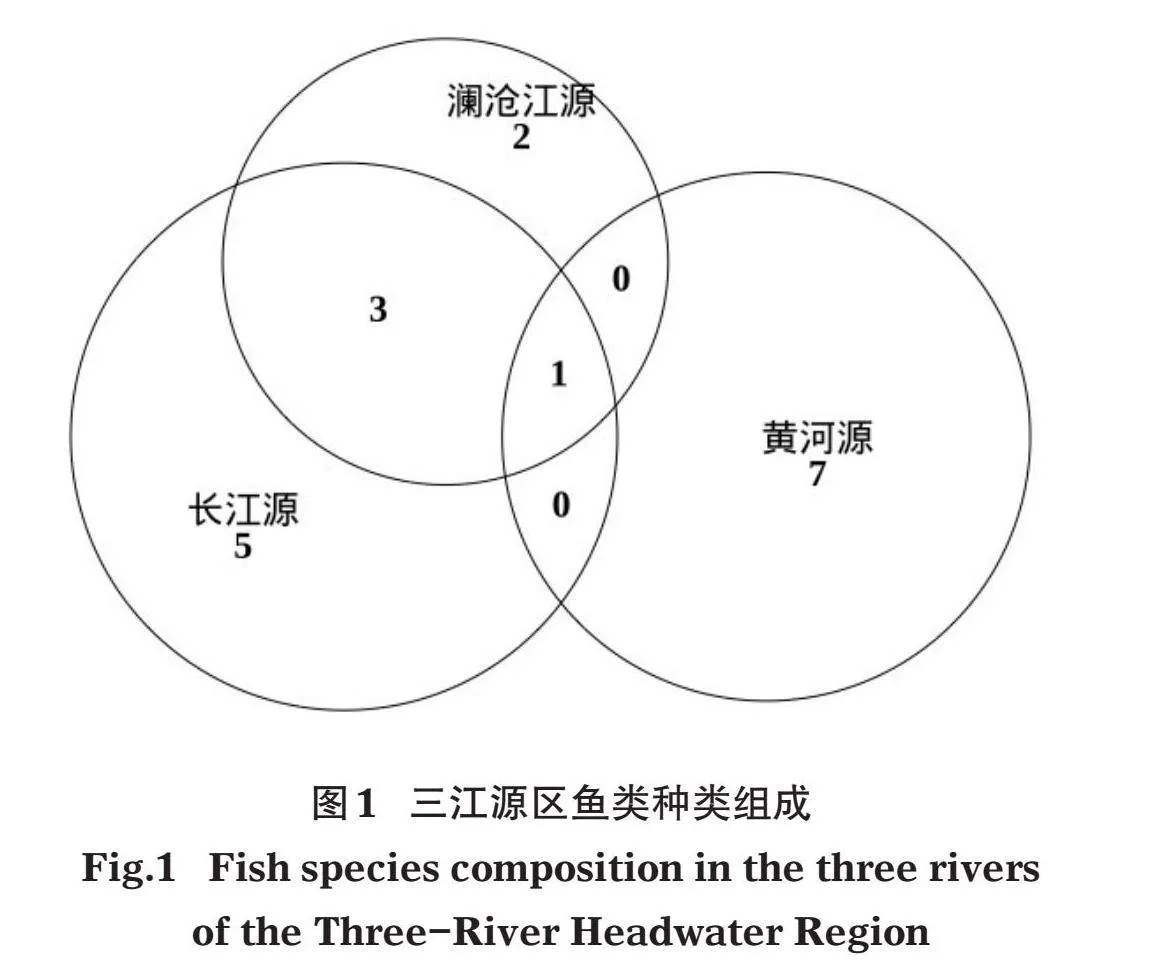
从分布区域来看,长江源区共采集到449尾、9种,黄河源区采集到115尾、8种,澜沧江源区采集到45尾、6种(表2、图1)。长江源、黄河源、澜沧江源均采集到的鱼类仅1种,即东方高原鳅(Triplophysa orientalis);仅分布于长江和澜沧江水系的鱼类为3种,分别是裸腹叶须鱼(Ptychobarbus dipogon)、细尾高原鳅(T. stenura)和前鳍高原鳅(T. anterodorsalis);黄河源鱼类组成与长江、澜沧江差异较大,除东方高原鳅外未发现共同种(表2)。
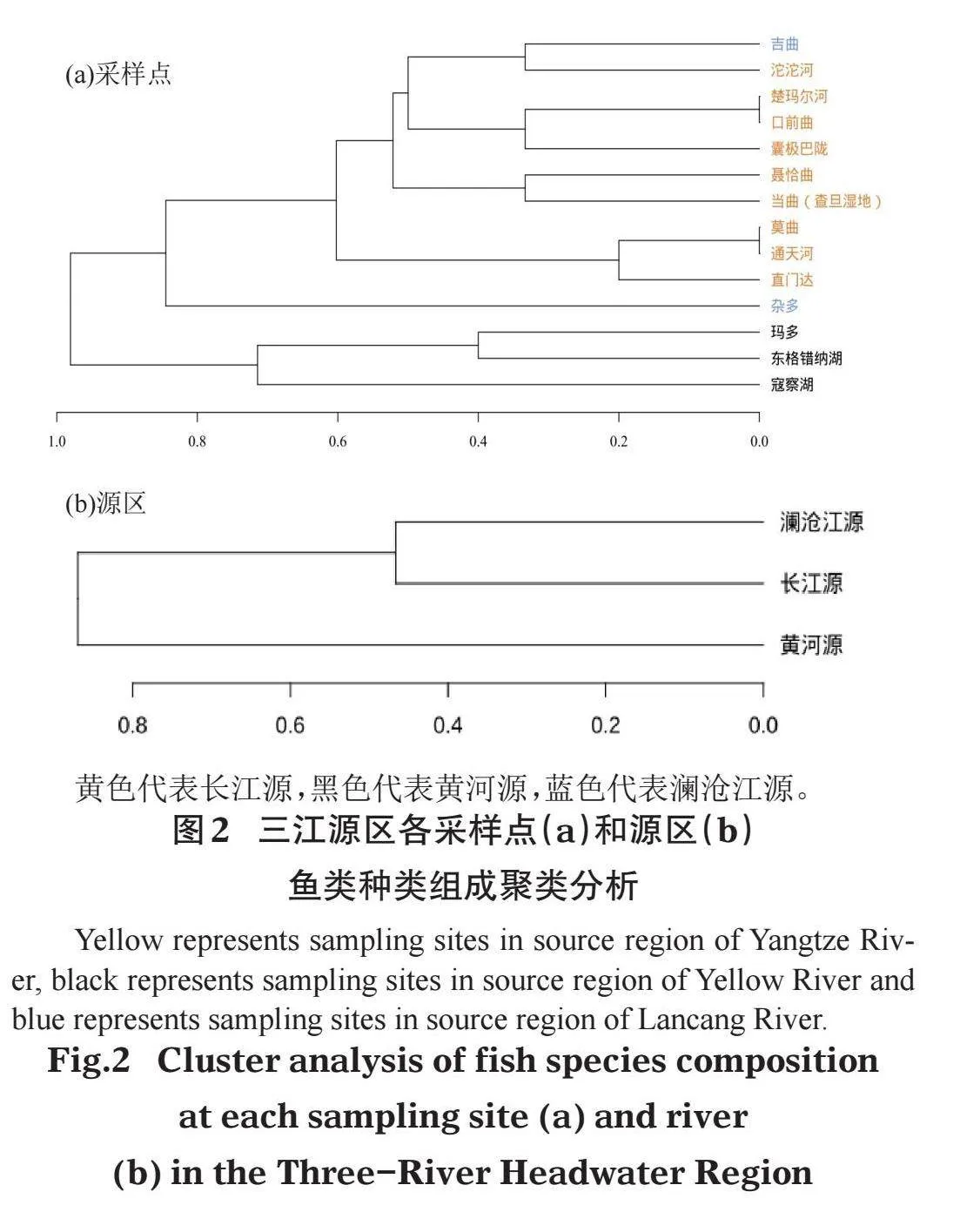
对各采样点和各水系鱼类种类组成进行聚类分析。按采样点来看(图2a),黄河源的东格错纳湖、寇察湖和玛多鱼类组成情况相近,聚为一大类;澜沧江源的吉曲与长江源的沱沱河鱼类组成相近,并与余下8个采样点聚为一小类,杂多样点的鱼类组成情况与长江源样点和澜沧江源的吉曲聚为一大类,表明长江源鱼类与澜沧江源相近。按河流水系来看(图2b),也呈现相似的结果,长江源和澜沧江源相近,黄河源则与长江源、澜沧江源差距较大。
2.2" "优势种组成
运用相对重要性指数(IRI)对三江源区鱼类优势种进行分析(表3),结果表明,3个水系优势种组成情况差异较大,其中长江源优势种3种,依次为细尾高原鳅、小头高原鱼(Herzensteinin microcephalus)、裸腹叶须鱼,在该水系的丰度占比分别为41.65%、34.52%和5.79%,生物量占比分别为4.58%、26.16%和60.56%;黄河源优势种2种,依次为麻尔柯高原鳅(T. markehenensis)和黄河裸裂尻鱼(Schizopygopsis pylzovi),丰度占比为68.70%和13.91%,生物量占比为44.78%和21.56%;澜沧江源优势种最多,有4种,依次为前腹裸裂尻鱼(S. anteroventris)、裸腹叶须鱼、东方高原鳅和细尾高原鳅,丰度占比为17.78%、15.56%、24.44%和26.67%,生物量占比为67.30%、20.54%、9.64%和1.27%。
2.3" "珍稀特有鱼类
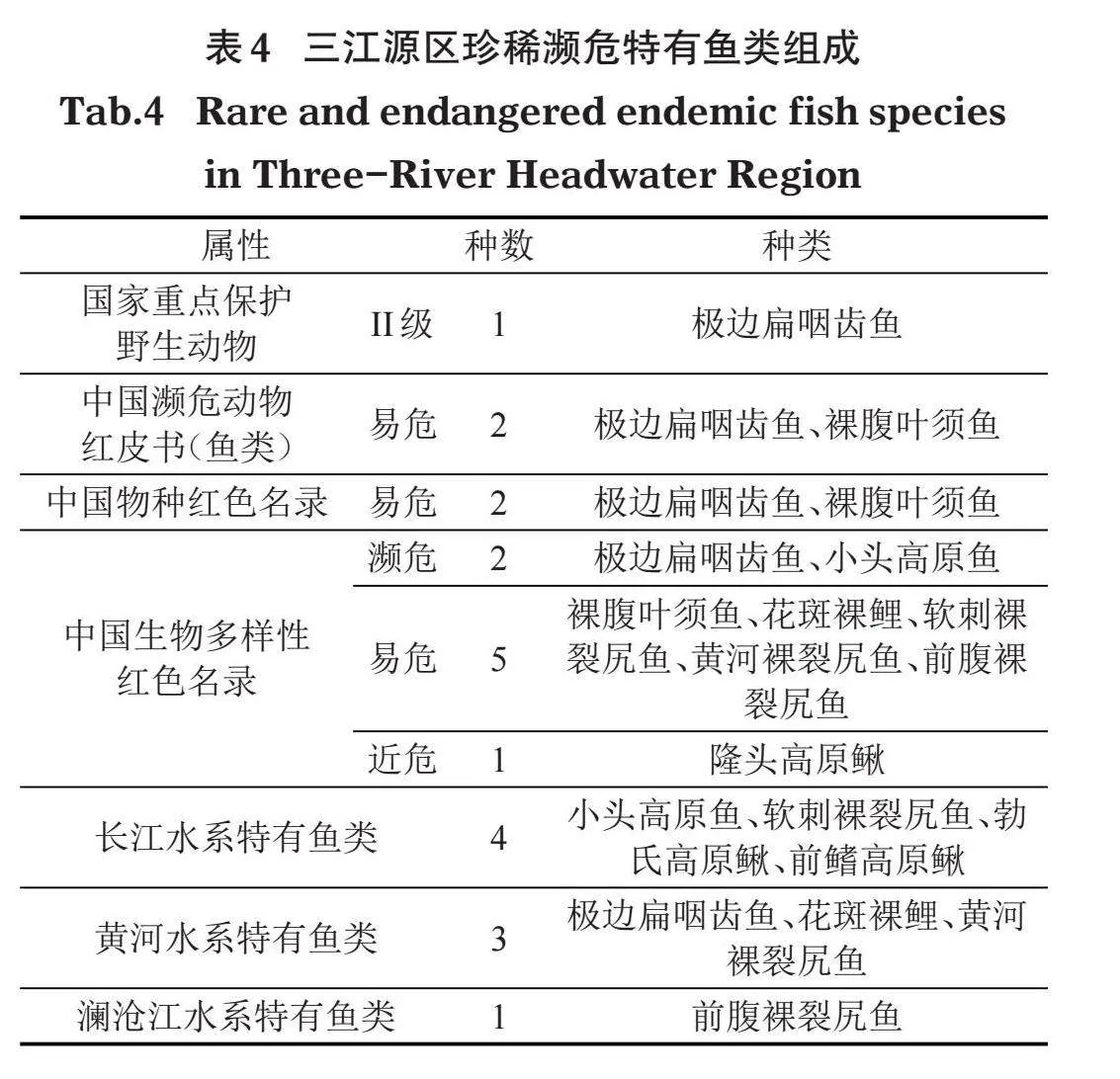
本次考察共采集到国家级重点保护鱼类1种,即极边扁咽齿鱼(Platypharod extremus)(国家林业和草原局和农业农村部,2021);列入《中国濒危动物红皮书(鱼类)》(乐佩琦和陈宜瑜,1998)、《中国物种红色名录》(汪松和解焱,2004)、《中国生物多样性红色名录——脊椎动物卷》(蒋志刚,2021)的鱼类共计8种,分别为极边扁咽齿鱼、小头高原鱼、裸腹叶须鱼、花斑裸鲤(Gymnocypris eckloni)、软刺裸裂尻鱼(S. malacanthus)、黄河裸裂尻鱼、前腹裸裂尻鱼、隆头高原鳅(T. alticeps)。长江水系特有鱼类4种,分别为小头高原鱼、软刺裸裂尻鱼、勃氏高原鳅(T. bleekeri)、前鳍高原鳅;黄河水系特有鱼类3种,分别为极边扁咽齿鱼、花斑裸鲤、黄河裸裂尻鱼;澜沧江水系特有鱼类1种,为前腹裸裂尻鱼(表4)。上述种类去重后共计10种,分别为极边扁咽齿鱼、裸腹叶须鱼、小头高原鱼、花斑裸鲤、软刺裸裂尻鱼、黄河裸裂尻鱼、前腹裸裂尻鱼、隆头高原鳅、勃氏高原鳅、前鳍高原鳅,占本次调查总种类数的55.6%,珍稀濒危特有种类比例较高,特别是裂腹鱼亚科的7种均为珍稀濒危特有鱼类。
2.4" "鱼类多样性
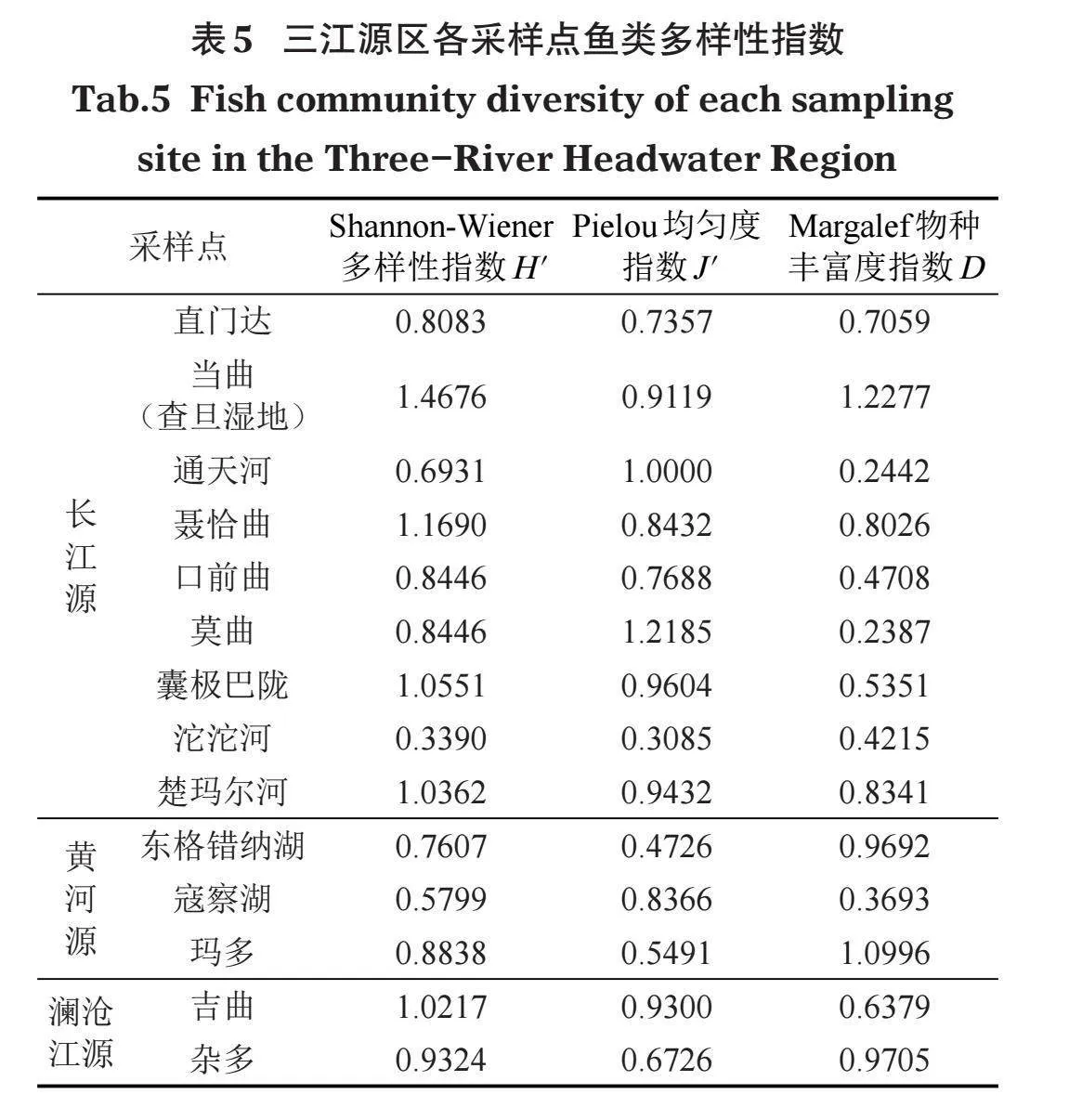
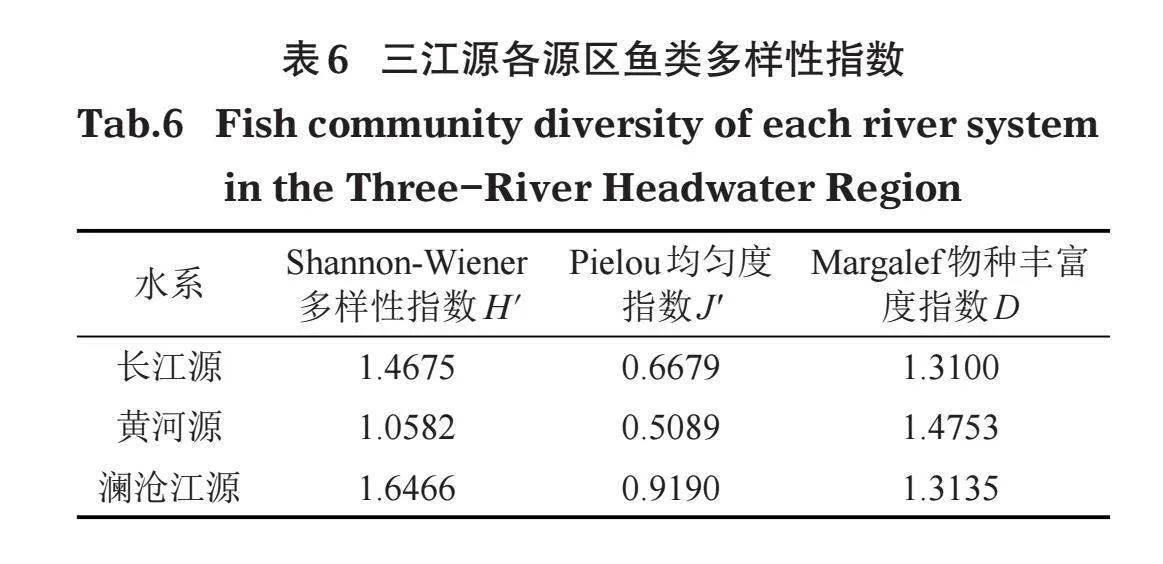
分别对各采样点及各水系渔获物多样性进行统计(表5),按采样点,Shannon-Wiener多样性指数前3的依次是当曲(查旦湿地)、聂恰曲、囊极巴陇,其次为寇察湖,再次东格错纳湖,最低的是沱沱河;Pielou均匀度指数前3的依次莫曲、通天河、囊极巴陇,其次为东格错纳湖,再次玛多,最低的是沱沱河;Margalef物种丰富度指数前3的依次是当曲(查旦湿地)、玛多、杂多,其次为通天河,再次为寇察湖,最低的是莫曲。
按河流水系(表6),Shannon-Wiener多样性指数由高到低依次为澜沧江源、长江源、黄河源,Pielou均匀度指数由高到低依次为澜沧江源、长江源、黄河源,Margalef物种丰富度指数由高到低依次为黄河源、澜沧江源、长江源。
3" "讨论
3.1" "三江源区鱼类组成特点
三江源区鱼类历史记录种类数为50种,其中长江源区鱼类22种,黄河源区鱼类24种,澜沧江源区鱼类9种(李柯懋等,2009;高桂香,2011;李志强等,2013)。本文调查到鱼类18种,其中长江源区9种,黄河源区8种,澜沧江源区6种。本次调查区域并未全面覆盖三江源区,主要在海拔4 000 m以上,相当于源头的源头,且采样强度不大,采集到的种类较少,但总体上也能反映出三江源更高海拔区不同水系的鱼类多样性。
三江源地区鱼类种类组成存在显著的地域差异,其中长江源和澜沧江源相近,黄河源则与长江源、澜沧江源差距较大。据报道,3大水系共有种类为2种,包括梭形高原鳅和修长高原鳅(李柯懋等,2009;高桂香,2011;李志强等,2013),与历史研究不同,本调查中3大水系共有种为东方高原鳅。已有研究发现除上述3种共有种外,仅在长江源和澜沧江源发现的共有种还有3种,分别是裸腹叶须鱼、细尾高原鳅和小眼高原鳅(李柯懋等,2009;高桂香,2011;李志强等,2013),本调查中仅存在于2大水系的共有种3种,其中2种与早期研究相同(裸腹叶须鱼和细尾高原鳅),第3种是前鳍高原鳅,小眼高原鳅在澜沧江源区未发现,这可能与调查频次和点位相对较少有关。长江源和黄河源也存在2种共有种,即斯氏高原鳅和东方高原鳅(李柯懋等,2009;高桂香,2011;李志强等,2013),其中本次3大水系调查均采集到东方高原鳅,但未发现斯氏高原鳅,除东方高原鳅外,本次调查未发现长江源和黄河源的鱼类共有种。
总体而言,三江源3大水系鱼类群落多样性较平原地区低(Lin et al,2021;Liu et al,2021),且不同水系鱼类群落多样性存在一定空间差异(李柯懋等,2009;高桂香,2011;李志强等,2013),造成这一结果的主要因素包括海拔、水温、水质等(Barletta et al,2005;Pico et al,2019),此外,本次调查范围和频次有限,调查结果尚不能反映总体情况,后续深入研究需开展不同季节和多年度鱼类资源调查,同时注意扩大调查范围,增加调查点位。
3大水系鱼类优势种组成也具有一定差异。高原鳅属鱼类由条鳅亚科原始类群随着青藏高原隆起演化而来,本次调查中3大水系优势种中均有高原鳅属鱼类,其适应于青藏高原高寒环境,是世界上已知分布海拔最高的鱼类,主要分布于青藏高原及其周边地区(武云飞和吴翠珍,1992;陈宜瑜等,1996)。
除麻尔柯高原鳅外,黄河裸裂尻鱼也是黄河源区优势种,这2种鱼的丰度占比、生物量占比和IRI值均显著高于其他鱼类。黄河水系特有鱼类极边扁咽齿鱼属于常见种,被《中国濒危动物红皮书(鱼类)》(乐佩琦和陈宜瑜,1998)和《中国物种红色名录》(汪松和解焱,2004)列为易危物种,此外,极边扁咽齿鱼被列入《国家重点保护野生动物名录》二级。
细尾高原鳅和裸腹叶须鱼是长江源区和澜沧江源区共同优势种。细尾高原鳅主要分布于青藏高原和四川西部境内的长江、澜沧江、怒江、雅鲁藏布江等水系,栖息在浅水卵石下,以底栖硅藻和水生昆虫等为食(武云飞和吴翠珍,1992),本次调查中,以长江源的通天河和当曲(查旦湿地)丰度最高。裸腹叶须鱼分布于金沙江水系、澜沧江、怒江上游干支流等,是长江源和澜沧江源大型鱼类(武云飞和吴翠珍,1992;熊芳园等,2022),在长江源的直门达、当曲(查旦湿地)、囊极巴陇和澜沧江源的吉曲、杂多均有一定资源量,其中当曲(查旦湿地)的裸腹叶须鱼资源量显著高于其他采样点。受捕捞活动和环境恶化等因素影响,裸腹叶须鱼种群也受到一定威胁,被《中国濒危动物红皮书(鱼类)》(乐佩琦和陈宜瑜,1998)和《中国物种红色名录》(汪松和解焱,2004)列为易危物种。
除细尾高原鳅和裸腹叶须鱼外,小头高原鱼是长江源区的又一优势种(熊芳园等,2022),主要分布于长江源各支流及湖泊中,以藻类、植物碎屑和水生昆虫等为食,个体大、分布广(武云飞等,1994)。当曲(查旦湿地)小头高原鱼生物量最高,其次是沱沱河。
澜沧江源区鱼类优势种最多,除上述2种优势种外,还有前腹裸裂尻鱼和东方高原鳅。有学者指出前腹裸裂尻鱼仅分布于澜沧江上游干支流(武云飞和吴翠珍,1992),本文结果符合这一观点,但仅在杂多调查到,这可能与澜沧江源区调查点位较少有关。东方高原鳅是栖息于溪流缓流或浅水多砂砾及水草处的底层鱼类,分布于长江和黄河干支流和附属水体(李亮涛等,2016),但本次在澜沧江源区也调查到东方高原鳅,是本次调查在该水系发现的新纪录种。
从本次调查结果看,长江水系与澜沧江水系的鱼类组成有较大共性,但黄河水系与长江和澜沧江水系的鱼类组成存在着较大差异,这也反映出在历史上长江与澜沧江之间较黄河可能曾有着更为广泛的交流和联系。从生物地理学角度分析,三江源地区人烟稀少,鱼类群落受人为因素影响较少,该地区鱼类主要受水系和海拔高度限制,青藏高原的隆升导致鱼类生存环境相应发生巨大变化,存活并延续下来的鱼类积累了适应环境的基因和特征(曹文宣等,1981;何德奎等,2003;张弥曼和DeSui,2016),不同水系受青藏高原隆起影响形成了不同环境特征,进而导致长江源、澜沧江源和黄河源形成了不同的鱼类群落组成和演化特征。因此,建议采用分子生物学等方法进行深入研究,探讨三江源的生物地理格局及其形成过程。
3.2" "三江源鱼类群落多样性研究与保护展望
三江源区人类活动较少,鱼类资源保存良好,水生生物保存相对完整(申志新和简生龙,2012)。近年来气候变化是人类关注的热点,可能对三江源鱼类群落产生影响,但三江源区水生生物资源调查较少,对珍稀特有鱼类的生物学基础理论和物种保护对策及措施研究等相对滞后。因此,亟需加强对三江源区的本底调查,关注气候变化对三江源水环境和水生生物的影响。
(1)加强本底调查,构建三江源区水生生物资源数据库。持续开展三江源地区多指标的水生生物本底调查,建立标准化科学监测体系,调查样点覆盖三江源区不同水系干支流、重要湖泊、湿地等不同水体类型,调查频率根据季节变化开展,积累长期基础调查资料,构建三江源区水生生物资源数据库,了解三江源区水生生物种类组成、分布和变化趋势,为流域水生生物多样性保护对策研究提供基础数据支撑。
(2)开展关键栖息地生态环境监测和保护。在鱼类繁殖场、索饵场、越冬场及洄游通道等重要栖息地设置监测点,动态监测栖息地生态环境和鱼类资源变化,加强对栖息地的保护力度,保障鱼类关键生活史不受干扰。
(3)构建三江源地区鱼类种质资源库。利用分子生物学方法分析不同水系鱼类遗传多样性,并采用现代技术保存三江源区鱼类种质资源样品,保存形式包括分子、细胞、组织、活体及种群样本等。
(4)关注气候变化对水生态环境的影响。三江源地区气候变化是当前的研究热点,全球变暖导致积雪融化,流域内河流、湖泊等水位升高,甚至出现咸水湖湖水漫溢,直接影响水生生物的群落结构。因此,需构建气候变化背景下水环境因子和鱼类群落结构变化预测模型,加强气候变化对水环境因子和鱼类种群结构影响研究。
(5)预防和控制外来鱼类入侵。三江源区局部放生活动可能是外来鱼类引入的主要途径,目前已有长江源和黄河源外来鱼类报道(唐文家和何德奎,2015),需采取严格的外来物种监管措施,加强外来鱼类入侵危害科普宣传,严禁外来鱼类引种、养殖和放生。
参考文献
曹文宣, 陈宜瑜, 武云飞, 等, 1981. 裂腹鱼类的起源和演化及其与青藏高原隆起的关系[M]// 中国科学院青藏高原综合科学考察队.青藏高原隆起的时代、幅度和形式问题. 北京:科学出版社:118-130.
陈锋, 常剑波, 郭玉, 等, 2012. 长江源区水生态调查及分析[C]//2012中国·青海绿色经济投资贸易洽谈会绿色发展高峰论坛暨三江源生态文明专题论坛论文集:84-93.
陈孝全, 苟新京, 2002. 三江源自然保护区生态环境[M]. 西宁:青海人民出版社.
陈兴, 余正勇, 2022. 三江源国家公园生态保护研究进展与展望[J]. 国土资源科技管理, 39(2):13-24.
陈宜瑜, 1998. 中国动物志硬骨鱼纲鲤形目(中卷)[M]. 北京:科学出版社.
陈宜瑜, 刘焕章, 陈毅峰, 1996. 青藏高原动物地理区的地位和东部界线问题[J].水生生物学报, 20(2):97-103.
高桂香, 2011. 三江源区鱼类及其它水生生物基本情况介绍[J]. 青海农牧业, (3):1.
郭忠胜, 马耀峰, 张志明, 等, 2009. 黄河源区气候变化及人为扰动的生态响应[J]. 干旱区资源与环境, 23(6):78-84.
国家林业和草原局, 农业农村部, 2021-02-09. 国家重点保护野生动物名录[EB/OL]. [2023-02-20]. https://www.gov.cn/xinwen/2021-02/09/content_5586227.htm.
何德奎, 陈毅峰, 陈宜瑜, 等, 2003. 特化等级裂腹鱼类的分子系统发育与青藏高原隆起[J]. 科学通报, 48(22):2354-2362.
蒋志刚, 2021. 中国生物多样性红色名录:脊椎动物卷[M]. 北京:科学出版社.
乐佩琦, 2000. 中国动物志硬骨鱼纲鲤形目(下卷)[M]. 北京:科学出版社.
乐佩琦, 陈宜瑜, 1998. 中国濒危动物红皮书:鱼类[M]. 北京:科学出版社.
李冠稳, 高晓奇, 肖能文, 2021. 基于关键指标的黄河流域近20年生态系统质量的时空变化[J]. 环境科学研究, 34(12):2945-2946.
李柯懋, 申志新, 陈燕琴, 等, 2012. 黄河班多段鱼类多样性初步研究及保护对策[J]. 水生态学杂志, 33(4):104-107.
李柯懋, 唐文家, 关弘韬, 2009. 青海省土著鱼类种类及保护对策[J]. 水生态学杂志, 30(3):32-36.
李亮涛, 杨学芬, 杨瑞斌, 等, 2016. 雅鲁藏布江中游东方高原鳅的年龄与生长特性[J]. 华中农业大学学报, 35(6):117-123.
李林, 李凤霞, 郭安红, 等, 2006. 近43年来“三江源”地区气候变化趋势及其突变研究[J]. 自然资源学报, 21(1):79-85.
李志强, 王恒山, 祁佳丽, 等, 2013. 三江源鱼类现状与保护对策[J]. 河北渔业, (8):24-30,38.
刘敏超, 李迪强, 温琰茂, 等, 2005. 三江源地区生态系统生态功能分析及其价值评估[J]. 环境科学学报, 25(9):1280-1286.
青海省环境保护厅, 2017. 2016青海省环境状况公报[R]. 西宁:青海省环境保护厅.
申志新, 简生龙, 2012. 三江源区水生生物状况及保护对策[J]. 青海农林科技, (3):37-40.
唐文家, 崔玉香, 赵霞, 等, 2012. 青海省澜沧江水系水生生物初步调查[J]. 水生态学杂志, 33(6):20-28.
唐文家, 何德奎, 2015. 青海省外来鱼类调查(2001-2014年)[J]. 湖泊科学, 27(3):502-510.
唐文家, 申志新, 简生龙, 2006. 青海省黄河珍稀濒危鱼类及保护对策[J]. 水生态学杂志, 26(1):57-60.
汪松, 解焱, 2004. 中国物种红色名录[M]. 北京:高等教育出版社.
武云飞, 吴翠珍, 1992. 青藏高原鱼类[M]. 成都:四川科学技术出版社.
武云飞, 吴翠珍, 于登攀, 1994. 青海可可西里地区的鱼类区系和地理区划[J]. 高原生物学集刊, 12:127-142.
西藏自治区水产局, 1995. 西藏鱼类及其资源[M]. 北京:中国农业出版社.
熊芳园, 刘晗, 陆颖, 等, 2022. 长江源区河流及典型湖泊丰水期水质与鱼类群落分析[J]. 中国环境监测, 38(1):86-94.
张觉民, 何志辉, 1991. 内陆水域渔业自然资源调查手册[M]. 北京:中国农业出版社.
张弥曼, DeSui Miao, 2016. 青藏高原的新生代鱼化石及其古环境意义[J]. 科学通报, 61(9):981-995.
郑德凤, 郝帅, 吕乐婷, 等, 2020. 三江源国家公园生态系统服务时空变化及权衡-协同关系[J]. 地理研究, 39(1):64-78.
朱松泉, 1995. 中国淡水鱼类检索[M]. 南京:江苏科学技术出版社.
Barletta M, Barletta-Bergan A, Saint-Paul U, 2005. The role of salinity in structuring the fish assemblages in a tropical estuary[J]. Journal of Fish Biology, 66(1):45-72.
Li X L, Perry G, Brierley G, et al, 2014. Quantitative assessment of degradation classifications for degraded alpine meadows (heitutan), Sanjiangyuan, western China[J]. Land Degradation amp; Development, 25(5):417-427.
Lin L, Deng W, Huang X, et al, 2021. Fish taxonomic, functional, and phylogenetic diversity and their vulnerabilities in the largest river in southeastern China[J]. Ecology and Evolution, 11:11533-11548.
Liu C, Zhu X, Bai Y, 2011. A study on runoff response to climate change and its prediction in the headwater region of the yellow river over the recent 49 years[J]. Resources Science, 33(7):1236-1241.
Liu H, Guo C, Qu X, 2021. Fish diversity, endemism, threats, and conservation in the Jinsha River basin (upper Yangtze River), China[J]. North American Journal of Fisheries Management, 41(4):967-984.
Margalef R, 1957. Information theory in ecology[J]. General System, 3:37-71.
Pico J A F, Carpio A J, Tortosa F S, 2019. Turbidity: A key factor in the estimation of fish species richness and abundance in the rocky reefs of Ecuador[J]. Ecological Indicators, 111:106021.
Pielou E C, 1975. Ecological diversity[M]. New York:Wiley.
Pinkas L, Oliphant M S, Iverson I, 1971. Food habits of albacore, bluefin tuna, and bonito in California water[J]. Fish Bulletin, 152:1-105.
Tao J, Kennard M J, Jia Y T, et al, 2018. Climate-driven synchrony in growth-increment chronologies of fish from the world's largest high-elevation river[J]. Science of the Total Environment, 645:339-346.
Wilhm J L, 1968. Use of biomass units in Shannon's formula[J]. Ecology, 49(1):153-156.
(责任编辑" "熊美华)
Fish Diversity Investigation in the Three-River Headwater Region
and Strategies for Conservation
CHEN Feng, YUAN Ting, XIONG Man‐tang, ZHAO Xian‐fu, MA Pei‐ming, ZHU Bin,
ZHANG Zhi‐yong, LIU Hui, DONG Fang‐yong, LI Jian‐yong
(Institute of Hydroecology, Ministry of Water Resources and Chinese Academy of Sciences,
Wuhan" "430079, P.R. China;Key laboratory of Ecological Impacts of Hydraulic-Projects and Restoration
of Aquatic Ecosystem of Ministry of Water Resources, Wuhan" "430079, P.R. China)
Abstract:The Three-River Headwater Region (TRHR) is the source of the Yangtze River, Yellow River and Lancang River. In this study, we analyzed the fish resource status in the TRHR, focusing on species composition, diversity and the spatial distribution of fish communities. The aim of the study was to provide scientific evidence for ecosystem protection and basin management in the TRHR. The study was based on a fishery resource investigation conducted at 14 sampling sites in the TRHR in August 2022. A total of 18 fish species were collected, including 9 species from Yangtze River, 8 species from Yellow River and 6 species from Lancang River. There was only one species (Triplophysa orientalis) common to all three rivers, and three species (Ptychobarbus dipogon, T. stenura and T. anterodorsalis) common to Yangtze River and Lancang River. Species composition in the Yellow River varied significantly from the other two rivers. According to the index of relative importance (IRI), there were 3 dominant species in the Yangtze River (T. stenura, Herzensteinin microcephalus and P. dipogon), 2 dominant species in the Yellow River (T. markehenensis and Schizopygopsis pylzovi) and 4 dominant species in the Lancang (S. anteroventris, P. dipogon, T. orientalis and T. stenura). Fish community diversity in the TRHR was relatively low. The highest fish diversity was in the source region of Lancang River, followed by the source region of Yangtze River and then Yellow River. Cluster analysis of fish species composition shows that the fish community in the source region of Yangtze River was similar to that in Lancang River and clearly different from the source region of Yellow River. In order to effectively protect fish resources in the TRHR, we recommend strengthening background investigations, ecological monitoring and habitat protection, building a database of fishery resources, attending to the impact of climate change on the aquatic ecological environment and preventing the invasion of exotic fish species.
Key words: fish diversity; spatial pattern; dominant species; conservation strategies; Three-River Headwater Region
