水稻DBB基因家族的生物信息学分析
2023-12-29郝爱平韩雪杨薛巨坤魏继承任如意
郝爱平,韩雪杨,薛巨坤,魏继承,任如意
(牡丹江师范学院生命科学与技术学院,黑龙江 牡丹江 157011)
【研究意义】锌指转录因子是植物中最重要的家族之一,参与植物的生长发育代谢及抵抗生物和非生物胁迫途径。锌指TF根据其个体成员的结构和功能特征分为几个亚家族。其中,B-box蛋白(B-box proteins,BBX蛋白)由于具有多种功能,近年来备受关注。BBX蛋白包含1个或2个靠近N-末端的保守B盒结构域,有些在靠近C-末端的地方有一个额外的CCT(CONSTANS,CO-like和TOC1)保守结构域。B-box分为两类,分别为B-box1(B1)和B-box2(B2)。2个B-box保守结构域根据其一致序列和锌结合残基之间的距离被识别[1]。进化过程中的片段复制和删除事件导致2个B盒结构域中的一致序列存在差异[2]。高度保守的CCT结构域由42~43个氨基酸组成,对功能性转录和核蛋白运输的调节非常重要[3]。在拟南芥中,已鉴定出14个 BBX 蛋白,这些成员根据 B-BOX 结构域和 CCT 结构域的存在分为5个亚组[4]。越来越多的证据表明植物 BBX 蛋白在多种生理和生化过程中发挥着关键作用,例如开花诱导、光形态发生、避荫反应、类胡萝卜素生物合成、生物和非生物应激反应[5]。【前人研究进展】BBX蛋白参与植物激素信号反应。脱落酸 (ABA) 是一种植物激素,当植物受到逆境胁迫时会被激活[6]。微矩阵分析检测到蔷薇中BBX基因的表达模式对ABA的响应不同[7]。此外,在番茄中发现BZS1/BBX20还参与油菜素类脂 (BR) 和光通路的信号转导[8]。马铃薯BBX蛋白在赤霉素 (GA) 信号通路中发挥潜在作用[9]。BBX18蛋白可以通过上调GA的表达水平来增强梨树下胚轴的生长,同时在不同光照情况下能够促进相关代谢基因的活性[10]。当白菜经生长素、GA和细胞分裂素处理时,不同的白菜BBX基因表达水平不同,并且白菜BBX基因在其启动子中具有激素响应顺式作用元件[11]。BBX 蛋白在果实发育中也具有重要作用,在花青素和类胡萝卜素的生物合成中,MdBBX22蛋白参与 MdHY5介导的信号转导并调节苹果皮中花青素的积累[12],SlBBX20蛋白通过直接激活乙烯合成相关基因的表达促进番茄叶绿体发育和类胡萝卜素积累[13]。表明,BBX蛋白可能参与植物激素信号传导。BBX基因也显示它们在非生物应激反应中的作用。在拟南芥中,初步鉴定耐盐蛋白 (STO,AtBBX24) 可以提高酵母细胞耐盐性[14],也可在高盐度条件下促进拟南芥根的生长。STO蛋白可以作为广泛范围应激相关基因的调控因子[15]。AtBBX18蛋白在光形态形成和提升热耐受性方面都具有重要作用[16]。在菊花中,CmBBX24蛋白除了具有延迟开花的作用外,还可以提高植物对寒冷或干旱的耐受性[17]。此外,与野生型植物相比,AtBBX24基因的过表达增强了耐盐性,并显著增加拟南芥的根长度[18]。【本研究切入点】水稻是世界上种植最广泛的粮食作物之一,深入分析水稻BBX基因家族并挖掘发育和胁迫相关重要基因对水稻育种意义重大,并且水稻中的BBX基因家族研究还未见报道。【拟解决的关键问题】通过对水稻的DBB(Double B-box)基因家族进行系统的生物信息学鉴定并研究其在不同组织中的表达情况,为后续深入研究水稻DBB基因家族功能提供基础。
1 材料与方法
1.1 数据来源
用于生物信息学分析的水稻全蛋白序列、全基因组序列以及GFF3文件下载自http://asia.ensembl.org/index.html,拟南芥DBB基因的蛋白序列下载自库TAIR(https://www.arabidopsis.org/index.jsp)。
1.2 水稻DBB基因家族成员的鉴定
根据拟南芥(Arabidopsisthaliana)已经鉴定出的DBB基因及其编码的蛋白序列,利用Pfam(http://pfam.xfam.org/)数据库和Hmmer 3.0软件构建隐马氏模型序列,对水稻全蛋白序列进行检索和去冗余,得到候选蛋白序列,其中阈值设为 E<1e-5。利用PfamScan (https://www.ebi.ac.uk/Tools/pfa/pfamscan/)和SMART (http://smart.embl-heidelberg.de/)在线平台对水稻所有候选基因的氨基酸序列结构域进行鉴定,含有DBB结构域的蛋白即为水稻DBB蛋白家族成员。并根据水稻DBB基因家族在染色体上的定位进行命名。
1.3 水稻DBB蛋白理化性质分析
利用ExPASy(https://web.expasy.org/protparam/)在线工具分析水稻DBB蛋白的物理和化学性质,包括氨基酸数目(NA)、分子量大小(MW)和等电点(pI)等;利用网站WoLF PSORT(https://wolfpsort.hgc.jp/)对其进行亚细胞定位预测分析。
1.4 水稻DBB基因染色体定位
水稻DBB基因染色体信息从基因注释文件中获得,整理成MAP文件,上传到MapInspect得到染色体位置可视化图片。
1.5 水稻DBB家族基因多重序列比对及结构域分析
使用DNAMAN软件对OsDBB氨基酸序列进行多重序列比对。利用NCBI和TBtools软件对OsDBB蛋白序列的保守结构域进行预测。
1.6 水稻DBB基因结构和蛋白保守基序预测
分别从水稻全基因组序列和基因注释GFF3文件中筛选OsDBB基因家族的 DNA和CDS序列,利用Gene Structure Display Server(GSDS)(http://gsds.gao-lab.org/index.php)在线网站对该基因家族的结构进行分析,并利用MEME Suite5.3.3(https://meme-suite.org/meme/)网站对水稻DBB蛋白保守基序进行预测,借助TBtools软件对该基因家族的保守基序进行可视化分析,其中motif最大数目为10。
1.7 水稻和拟南芥DBB基因家族系统发育树的构建
利用MEGA11.0软件中的Align by Clustal W工具对水稻和拟南芥DBB蛋白序列进行多重序列比对,各个参数设定为默认值,并用其 Phylogeny工具以 Neighbor-Joining 法构建系统发育进化树,Bootstrap值设定为1000。
1.8 水稻DBB基因启动子区域的顺式调控元件分析
利用TBtools软件提取OsDBB基因CDS序列上游2000 bp序列,通过PlantCare数据库(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对OsDBB基因的顺式调节元件进行分析。
1.9 水稻DBB基因在不同组织中的表达模式
利用BAR数据库(https://www.bar.utoronto.ca/)检索水稻不同生长阶段各组织器官中水稻DBB基因的表达水平,并构建热图,筛选在水稻生长发育中起关键作用的DBB基因。
2 结果与分析
2.1 水稻DBB基因家族成员的鉴定
使用14个拟南芥DBB蛋白序列作为查询序列,对水稻基因组数据利用Pfam(http://pfam.xfam.org/)数据库和Hmmer3.0软件构建隐马氏模型序列,对水稻全蛋白序列进行检索和去冗余,得到候选蛋白序列,对假定的OsDBB蛋白质序列进行SMART和Pfam工具测试DBB结构域的存在。鉴定的13个OsDBB均含有2个DBB结构域。
2.2 水稻DBB蛋白理化性质分析
对OsDBB蛋白进行理化性质(表1)分析,OsDBB蛋白氨基酸数量在181(OsDBB6)~ 378(OsDBB8),分子量在21.62(OsDBB4)~39.40(OsDBB8)KDa,等电点 (pI) 在4.93(OsDBB2)~5.98(OsDBB1),且全部为酸性蛋白pI<7.5,OsDBB蛋白不稳定系数为41.76 ~66.62,其中所有OsDBB蛋白为不稳定蛋白(II>40),脂肪系数在54.63(OsDBB8)~79.5(OsDBB6),平均亲水性值在-0.137(OsDBB13)~-0.685(OsDBB12),且亲水性值都小于0,表明OsDBB蛋白均为亲水性蛋白。水稻DBB蛋白在理化性质上存在差异,说明这些成员可能在水稻生长发育过程中起着不同的调控作用。
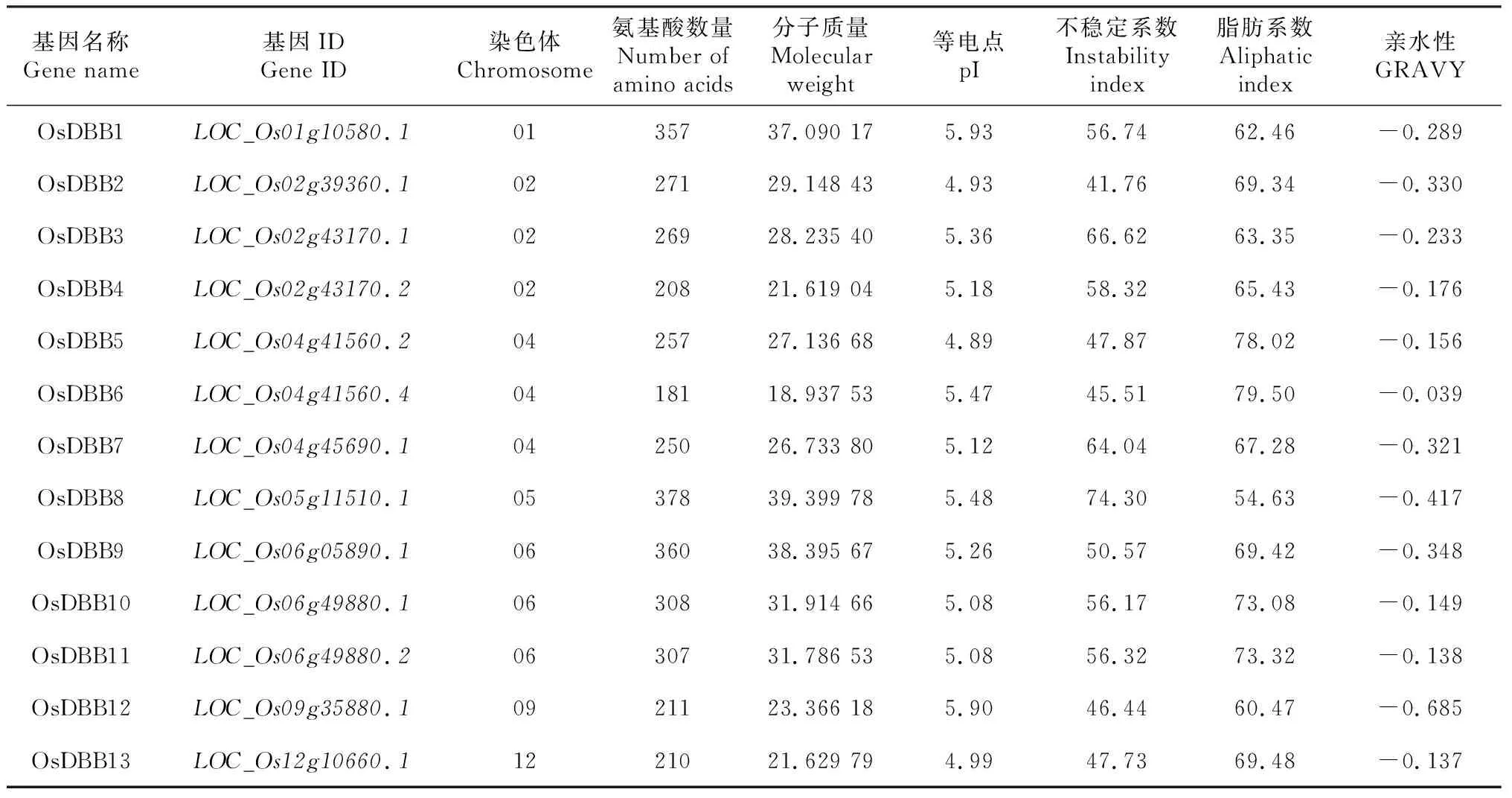
表1 OsDBB蛋白理化性质分析Table 1 Analysis of physicochemical properties of OsDBB proteins
2.3 水稻DBB基因染色体定位
根据水稻基因组中的注释信息,发现13个水稻DBB基因分布在12条水稻染色体中的7条上(图1)。每个染色体DBB基因数量为1或3个,其中Chr01、Chr05、Chr09和Chr12号染色体上分别含有1个OsDBB基因,Chr02、Chr04和Chr06号染色体上分别含有3个OsDBB基因。
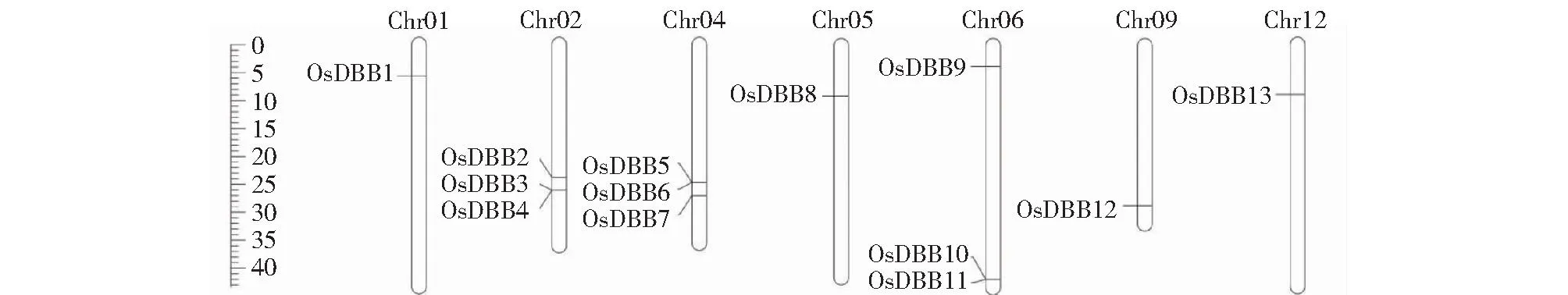
图1 OsDBB家族基因染色体定位分析Fig.1 Localization analysis of OsDBB family gene
2.4 水稻DBB家族基因多重序列比对及结构域分析
利用DNAMAN软件对OsDBB蛋白序列进行多重序列比对发现,该家族基因成员相似性达32.46%,且保守区域主要集中在N端(图2)。同时对OsDBB蛋白序列的结构域(图3)分析发现,该家族主要含Bbox1-BBX-like和BBOX 2个结构域,且主要位于OsDBB蛋白序列的N端,与序列同源性比对结果一致。

图2 水稻DBB氨基酸多重序列比对Fig.2 Multiple sequence alignment of rice DBB amino acid
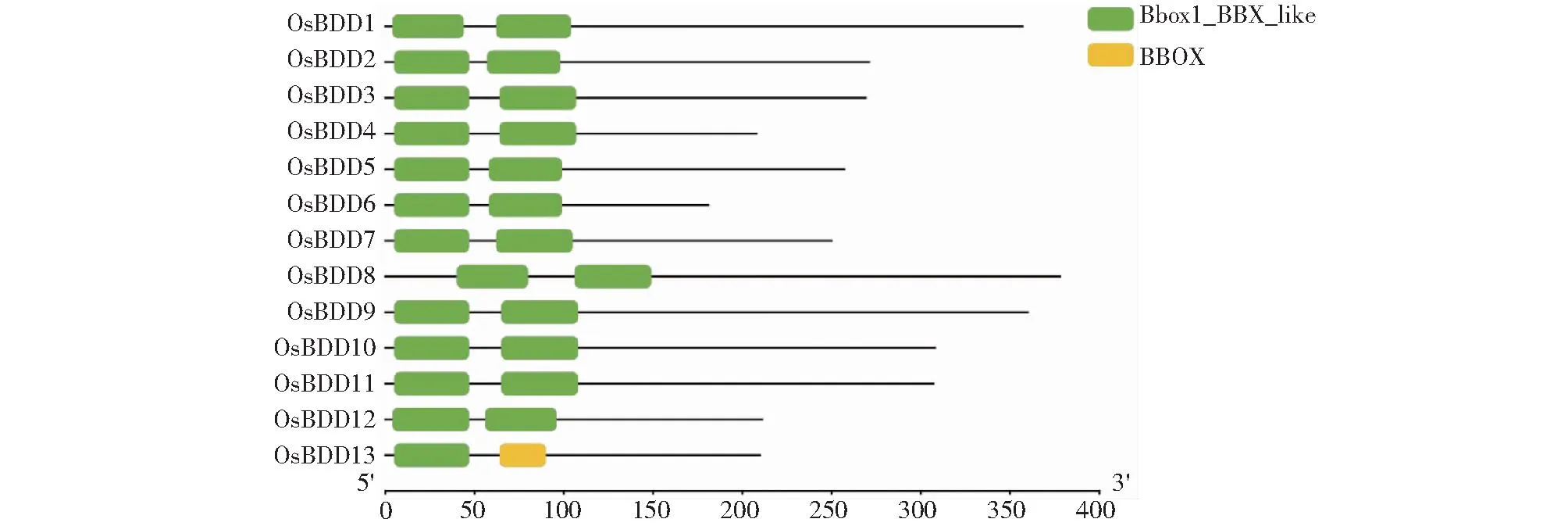
图3 水稻DBB基因结构域Fig.3 Rice DBB gene domain
2.5 水稻DBB基因结构和蛋白保守基序的分析
从图4-A可知,OsDBB基因家族成员的内含子和外显子数目存在一定的差异,具体表现在OsDBB1、OsDBB2、OsDBB6、OsDBB10、OsDBB11和OsDBB13各含有2个外显子和1个内含子,而OsDBB12含有5个外显子和4个内含子,其余基因均含有3个外显子和2个内含子。基因聚类(图4-B)发现,水稻13个DBB基因中有5个旁系基因对,分别为OsDBB1和OsDBB8、OsDBB10和OsDBB11、OsDBB3和OsDBB4、OsDBB5和OsDBB6、OsDBB12和OsDBB13,每个旁系基因对的基因结构相似,推测基因功能类似。

A:OsDBB基因结构分析;B:OsDBB基因进化树;C:OsDBB基因家族motif预测;D:motif氨基酸序列分析 A:OsDBB gene structure analysis; B:The Phylogenetic tree of OsDBB gene; C:Motif prediction of OsDBB gene family; D:Sequence analysis of motif amino acid.图4 水稻DBB基因结构和motif预测Fig.4 Rice DBB gene structure and motif prediction
从图4-C可知,OsDBB基因家族成员的motif数量及排列均存在一定的差异,其中motif 1在13个OsDBB蛋白中均含有分布;而motif 2分布在除OsDBB12蛋白外的其他12个OsDBB蛋白序列中,且主要分布在蛋白质序列的N端;motif 3和motif 4主要分布在OsDBB2、OsDBB3、OsDBB4和OsDBB7蛋白序列中;motif 4主要位于蛋白质序列的C端;motif 5主要分布在OsDBB3、OsDBB4和OsDBB9蛋白质序列中;motif 6、motif 7和motif 8主要分布在OsDBB10和OsDBB11的蛋白质序列中,motif 9在OsDBB2和OsDBB5都有分布;motif 10则分布在OsDBB1、OsDBB5、OsDBB6、OsDBB8和OsDBB9蛋白质中。另外,每个motif的序列和长度均存在差异(图4-D),其中motif 9含有18个氨基酸,motif 1、motif 2和motif 6均含有50个氨基酸,其余氨基酸数目21~45个不等。
2.6 水稻DBB蛋白多序列比对及系统发育树
聚类分析结果(图5)显示,27个DBB蛋白分为Group 1、Group 2、Group 3和Group 4 4个亚族,每个亚族均含有拟南芥DBB蛋白,但数量均存在一定差异,其中Group 4亚族中成员最多,含有13个DBB家族成员,水稻和拟南芥分别含有7和6个DBB成员,Group 1亚族含有3个AtDBB,无水稻DBB基因家族成员,是含DBB基因家族成员最少的亚族,而Group 2和Group 3各含4和7个DBB基因家族成员,其中2个亚族均含有2个AtDBB基因。表明,水稻DBB大多数家族成员与拟南芥DBB家族成员进化关系较近,推测其功能相似。

图5 水稻和拟南芥DBB基因家族系统发育分析Fig.5 Family phylogenetic analysis of DBB gene in rice and A.thaliana
2.7 水稻DBB基因启动子区域的顺式调控元件分析
在13个水稻OsDBB基因启动子区域中共预测27种顺式作用元件(图6)。其中,参与生长发育的顺式作用元件有7种,分别为ARE、CAT-box、GC基序、GCN4基序、O2-site、RY和TGA元件;参与植物激素和非生物胁迫的顺式作用元件分别有4种(TATC-box、CGTCA基序、ABRE和TGACG基序)和6种(TCA元件、CCAAT-box、LTR、MBS、MRE和富含TC的基序),参与光响应的有10种(ATCT基序、Box4、GATA基序、G-box、GT1基序、I-box、Sp1、TCCC基序、TCT和GTGGC基序)。在预测的植物激素反应元件中,ABRE最丰富,参与脱落酸反应;TGACG和CGTCA基序次之,且数量相等,均参与茉莉酸甲酯(MeJA)的调控途经。说明,OsDBB基因可能参与脱落酸和茉莉酸甲酯调控途径。在确定的6种与非生物胁迫相关的顺式作用元件中,主要涉及干旱胁迫(MBS),参与低温反应(LTR)和防御和压力反应(富含TC的重复)的顺式作用元件。说明,OsDBB基因可能参与对逆境的反应。

图中颜色表示顺式作用元件的数量,颜色越深数量越多,其中白色表示无相关顺式作用元件; ARE:参与无氧诱导ABRE:参与脱落酸反应的顺式调节元件。 AuxRR-core:参与生长素反应的顺式调节元件。 CGTCA 基序:参与茉莉酸甲酯 (MeJA) 反应的顺式作用调节元件。 GARE-motif:赤霉素反应元件。 TGACG 基序参与 MeJA 反应的顺式调节元件。 TGA-元件:生长素反应元件。 ERE:顺式乙烯反应元件。 P-box:赤霉素反应元件。 GC-motif:参与特定缺氧诱导的增强子样元件。 LTR:参与低温反应的顺式作用元件。 MBS:与干旱诱导相关的 MYB 结合位点。 STRE:压力应元素。富含 TC 的重复序列:参与防御和压力反应的顺式作用元件。 WUN-motif:伤口反应元素。 MYC:参与干旱胁迫的顺式作用元件。 W框:参与糖代谢和植物防御信号的顺式作用元件。 DRE核心:脱水反应元件。 ARE:厌氧诱导的顺式调节元件。The color in the figure indicates the number of cis acting elements. The darker the color, the more the number. Among them, white indicates that there are no relevant cis acting elements;ARE: Involved in anaerobic induction. ABRE: Cis-regulatory element involved in abscisic acid reaction. AuxRR-core: A cis-regulatory element involved in the auxin reaction. CGTCA motif: Cis-acting regulatory element involved in the reaction of methyl jasmonate (MeJA). GARE-motif: Gibberellin reaction element. The TGACG motif is a cis-regulatory element involved in MeJA response. TGA-element: Auxin reaction element. ERE: Cis-ethylene reaction element. P-box: Gibberellin reaction element. GC-motif: Enhancer like element involved in specific hypoxia induction. LTR: Cis-acting element involved in low temperature reaction. MBS: MYB binding site associated with drought induction. STRE: Stress element. Tc-rich repeat sequences: Cis-acting elements involved in defense and stress responses. WUN-motif: Wound response element. MYC: Cis-acting element involved in drought stress. W frame: Cis-acting element involved in glucose metabolism and plant defense signaling. DRE core: Dehydration reaction element. ARE: Anaerobically induced cis-regulatory element.图6 水稻启动子顺式作用元件分析Fig.6 Analysis of rice promoter cis-acting elements
2.8 水稻DBB基因在不同组织中的表达模式
分析植物器官特异性基因表达模式可为基因在发育过程中的可能功能提供线索。由图7可知,所有水稻OsDBB基因在叶、根、种子和花序中的表达量较高。总体来看,所有水稻OsDBB基因茎尖分生组织的表达量低于其他组织,其中OsDBB1、OsDBB5、OsDBB8和OsDBB13基因在叶中表达量较高,OsDBB2、OsDBB7、OsDBB10、OsDBB11和OsDBB12在根中的表达量较高,OsDBB3、OsDBB4和OsDBB6在种子有较高的表达量,在花序中只有OsDBB9和OsDBB12表达。不同水稻OsDBB基因在不同组织中表达各不相同,并具有一定的特异性,表明OsDBB基因在水稻的生长发育过程中具有重要作用。

右侧标尺数值范围代表均一化后表达量的变化范围,红色表示高表达,蓝色表示低表达。The numerical range of the scale on the right represents the variation range of expression after homogenization, red indicates high expression and blue indicates low expression.图7 水稻DBB基因在不同组织表达差异分析Fig.7 Analysis of differences in expression of rice DBB gene in different tissues
3 讨 论
植物的转录因子(TFs)是一类通过调控下游基因转录而在生理生化过程中发挥重要作用的蛋白质。TF结构构建通常有4个主要域,所有这些域都是功能过程所必需的:DNA结合位点、转录激活域、寡聚化位点和核定位信号。其中,B-box(BBX)锌指家族的研究是近年来发展的领域。DBB基因家族是BBX的一个亚族,在植物生长发育和响应逆境胁迫中有重要作用。近年来,DBB基因家族成员已在多个植物种被鉴定,如橡胶树、芒果、红花和白菜中各鉴定出9、9、26和18个,并对其功能进行了报道,但关于水稻的报道较少。本研究根据拟南芥14个DBB基因家族成员,通过构建HMM文件对水稻DBB基因家族进行基因组水平的鉴定和分析,在水稻中鉴定出13个DBB基因,并根据染色体定位命名为OsDBB1~OsDBB13,发现水稻DBB基因家族成员数量与其他物种的数量相近,说明DBB基因家族在进化过程中相对保守。根据OsDBB和AtDBB蛋白保守序列的同源性和亲缘性,将其分为4个亚族,这与张鸿韬等[19]在巴西橡胶树和赵乐等[20]在红花中的分类一致。通过对水稻DBB基因家族成员的基因结构和motif分析发现,同一亚族DBB基因结构和motif极其相似,motif 1出现在水稻的13个DBB基因中,且主要位于蛋白质序列的N端,而对水稻DBB蛋白的保守结构域进行可视化分析发现,该结构主要位于蛋白质序列N端,且含有2个,因此推测motif 1很可能是水稻DBB蛋白的保守结构域。
基因家族成员的外显子/内含子多样化通过3种主要机制在多个基因家族的进化中发挥重要作用:外显子/内含子获得/丢失、外显子/假外显子化和插入/缺失。本研究发现,具有高度相似的外显子/内含子结构属于同一系统发育进化分枝,这与在小拟南芥和辣椒中的研究结果一致[21-22]。在几个水稻DBB基因的编码序列中观察到外显子/内含子长度存在明显差异,这可能导致产生功能不同的旁系同源基因。
植物在其生命周期中会暴露于一系列非生物和生物胁迫中,同时植物也进化出多种保护自身免受生物和非生物胁迫的途径,主要反应在胁迫反应基因和其表达上,并且基因的表达受启动子或转录因子的调控。本研究对水稻DBB基因CDS区上游2000 bp进行启动子区顺式作用元件分析,水稻DBB基因启动子区域中的部分元件数量不尽相同,其中激素类响应顺式作用元件最多,这些激素在植物对生物和非生物胁迫的响应以及植物的信号转导中发挥关键作用,如脱落酸(ABA)、茉莉酸(MeJA)和水杨酸(SA)等小信号分子介导植物响应以抵御压力[23-24],对于植物在自然界中的生存至关重要。其中茉莉酸甲酯及其生物活性衍生物是植物对生物胁迫(如伤害、病原体感染和昆虫攻击)以及非生物胁迫(如干旱和臭氧暴露)反应的关键调节剂[25]。在健康、未受伤的植物组织中,茉莉酸盐在控制多种重要发育过程中发挥广泛作用,包括根系生长、种子萌发、卷须卷曲、花发育和衰老[26]。而在水稻DBB基因启动子区域中脱落酸、茉莉酸甲酯响应元件数量最多(ABRE、CGTCA-motif)[27]。此外,水稻DBB基因除CAAT-box、TATA-box等启动子基本核心元件外,还包含大量参与光响应的元件(AE-box、ATCT-motif、Box 4、GT1-motif、LAMP-element、TCT-motifand chs-CMA1a),表明OsDBB基因在水稻生长发育和响应非生物胁迫中具有重要作用。
4 结 论
本研究首次对水稻DBB基因家族进行基因组学分析,共鉴定出13个OsDBB基因。水稻OsDBB基因的启动子序列含有与生长发育、激素、应激反应以及光响应有关的调控元件。不同水稻OsDBB基因在不同组织中的表达各不相同,具有一定的特异性。推测水稻OsDBB基因在植物生长发育和响应非生物胁迫中具有重要作用。
