晒田环境胁迫对克氏原螯虾卵巢发育相关生化指标和激素的影响
2023-12-29黄玉英何苹萍郑玉斯卢智发张圣杰李文红彭金霞王大鹏
袁 畅,李 蔚,黄玉英,何苹萍,郑玉斯,李 鑫,卢智发,张圣杰,李文红,彭金霞,王大鹏
(1.广西水产科学研究院/广西水产遗传育种与健康养殖重点实验室,南宁 530021;2.广西大学动物科学技术学院,南宁 530004)
【研究意义】克氏原螯虾(Procambarusclarkii)原产于北美洲,20世纪30年代引进中国,如今已成为重要养殖经济虾类之一,主要靠自然繁殖供给苗种[1]。自然繁殖亲本主要是捕捞残留的成虾,大多个体小,生长速度慢,繁殖速度慢且繁殖时间长[2]。当克氏原螯虾生活环境发生巨大变化(如干旱、高温酷暑)时,其种族延续与繁殖需求将变成生存的首要选择,这种情况下繁殖性能将产生一定变化,如在环境胁迫下克氏原螯虾会出现早繁、多繁现象[3]。晒田环境胁迫是一种人工诱导繁殖的常用方法,近年来在生产上广泛使用,但其技术尚未标准化,晒田起始时间和晒田时长主要依靠生产经验决定。因此,急需研究晒田环境胁迫下克氏原螯虾体内生理调节机制,为改进生产操作提供理论依据。【前人研究进展】在不同胁迫环境下,水产动物将从神经内分泌、生理生化、免疫调节等方面进行协同调控,以维持内环境稳定[4]。如青海湖裸鲤(Gymnocyprisprzewalskii)在碱度胁迫下,AKP/ALP和ACP活性随碱度升高或降低均呈先下降后上升的变化趋势;GSH-Px活性随碱度升高呈先上升后下降的变化趋势,随碱度降低则呈先下降后上升的变化趋势[5];太平洋白虾(Pacificwhiteshrimp)的PGF2α和PGE2浓度在3.0%盐度下显著高于1.5%盐度[6];随着褪黑激素浓度的增大,锯缘青蟹(Scyllaserrata)体内MF与Ecdysone水平显著增加[7];凡纳滨对虾(Litopenaeusvannamei)在急冷与空气暴露联合胁迫下,随着胁迫时间的延长,其Cortisol含量呈下降趋势[8];在使用双酚A胁迫福寿螺的研究中,性类固醇激素(Estrone、17β-E2、T和PROG)含量随着双酚A浓度持续升高而显著降低[9]。关于环境胁迫下水产动物高血糖家族激素(CHH、GIH、MIH和MOIH)含量变化的研究则鲜见报道。【本研究切入点】克氏原螯虾抵抗恶劣环境的能力较强,对晒田环境胁迫具有独特的适应能力,在此过程中可促进卵巢发育及繁殖,但目前鲜见关于晒田环境胁迫下克氏原螯虾体内生化与激素调节机制的报道。【拟解决的关键问题】本研究对比分析晒田环境胁迫下克氏原螯虾各项生化指标变化、激素调节机制及对卵巢发育的影响,为晒田环境胁迫促进克氏原螯虾卵巢成熟这一生产操作的改良和标准化提供理论依据。
1 材料与方法
1.1 试验材料
试验所用克氏原螯虾取自广西来宾市的一个商业养殖场。2022年9月,分别选取50 kg克氏原螯虾(雌雄比1∶1,雌雄各25 kg)放至2个池塘中,1周后开始正式试验。其中1个池塘改造后作为晒田池塘,池内所有试验虾为晒田组,另一池塘为不晒田的对照池塘,池内所有试验虾为对照组。2个池塘投放数量均大于2000尾。晒田池塘和对照池塘面积均约为1300 m2,底质均为稻田壤土,土壤保水力较强。
1.2 试验方法
1.2.1 试验设计 试验期间天气晴朗,无降雨。将晒田池塘的田面(300 m2)用塑料围子和铁管围牢,防止逃逸,对照池塘不需改造。2个池塘水深30~40 cm,提前1周放虾入池至田面。正式试验开始后将晒田池塘田面水排干,使田面自然曝晒,对照池塘正常养殖。田面在白天日照时间约12.5 h、平均温度34.1 ℃的条件下自然暴晒,其底泥在排水时仍为软泥底,但不足以使虾躲避,经1~3 d自然暴晒,达到田块中间不陷脚,田边表土无裂缝和发白的板结状态,期间可发现虾挖掘洞穴进行躲避。
1.2.2 样品采集 在晒田胁迫0、3、7、10 d时分别从晒田组和对照组中采集30只雌性克氏原螯虾。晒田组中虾平均24.61 g,对照组中虾平均23.56 g,二者规格接近。晒田组从田面挖掘虾洞进行采集(晒田胁迫0 d采集初始样本,无需挖洞采集),对照组在环沟中放置虾笼进行采集。记录样品体重、卵巢重量、肝胰腺重量和卵巢颜色,根据Zhong等[10]对克氏原螯虾卵巢发育分期的鉴定方法,卵巢发育至IV期鉴定为卵巢成熟。每个采样时间段随机取5只雌性克氏原螯虾的组织作为一个混样(每个时间段3个混样),在冰上取出肝胰腺和卵巢一并放入冻存管,所有组织均置于液氮中保存备用。从每只虾的围心腔抽取1 mL血淋巴放入抗凝管,同一混样血淋巴进行混合,于4 ℃冰箱静置20 min,4 ℃下2500 r/min离心10 min,取其上清液分装至离心管,得到血清。检测卵巢、肝胰腺和血清的生化指标,并检测血清激素。
性腺指数(GSI)=卵巢重量/体重×100%
肝体指标(HSI)=肝胰腺重量/体重×100%
1.2.3 生化指标与激素检测 碱性磷酸酶(AKP/ALP)、酸性磷酸酶(ACP)和谷胱甘肽过氧化物酶(GSH-Px)采用苏州科铭生物技术有限公司试剂盒检测;前列腺素 F2α(PGF2α)、前列腺素 E2(PGE2)、雌酮(Estrone)、17β-雌二醇(17β-E2)、睾酮(T)、孕酮(PROG)、高血糖激素(CHH)、性腺抑制激素(GIH)、蜕皮抑制激素(MIH)、大颚器抑制激素(MOIH)、蜕皮激素(Ecdysone)、甲基法尼酯(MF)和皮质醇(Cortisol)则采用江苏晶美生物科技有限公司ELISA 试剂盒检测。以上每个生化指标或激素检测均进行3次重复。
1.3 分析方法
试验数据采用Excel 2019和SPSS 26.0软件进行统计分析。
2 结果与分析
2.1 晒田环境胁迫前后克氏原螯虾卵巢发育情况
如表1所示,晒田胁迫0 d时,晒田组与对照组雌性克氏原螯虾卵巢发育各分期分布数量差异不明显,晒田胁迫10 d时,晒田组雌性克氏原螯虾主要分布在IV期,对照组雌性克氏原螯虾主要分布在III期和IV期。晒田胁迫0 d时,晒田组与对照组的雌性克氏原螯虾卵巢成熟率相近,晒田胁迫10 d时,晒田组雌性克氏原螯虾卵巢成熟率升至96.67%,但对照组雌性克氏原螯虾卵巢成熟率仅升至60.00%。
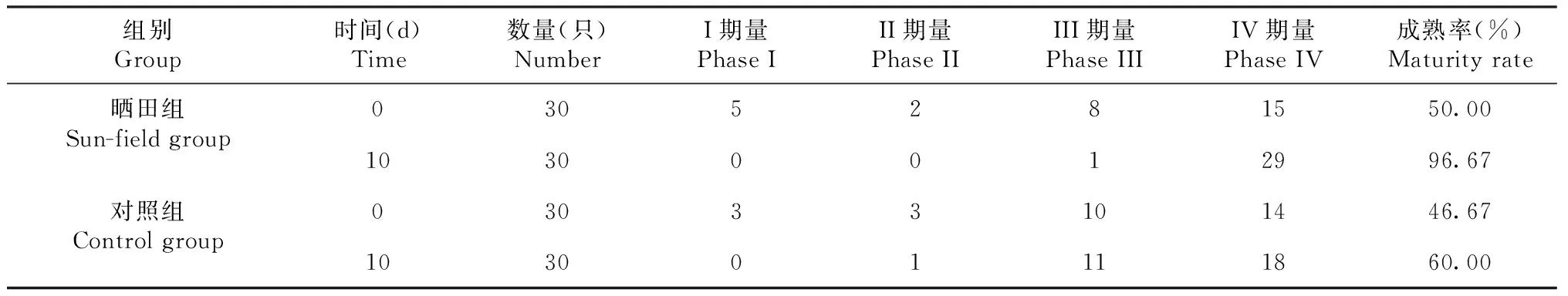
表1 晒田前后克氏原螯虾卵巢发育阶段分布及成熟率变化Table 1 Changes in ovarian development stage and maturity rate of P.clarkii before and after sun-field
2.2 晒田环境胁迫对克氏原螯虾生化指标的影响
如图1所示,晒田胁迫0 d时,晒田组雌性克氏原螯虾卵巢中AKP/ALP、ACP、GSH-Px、PGF2α和PGE2含量与对照组雌性克氏原螯虾无显著差异(P>0.05,下同);晒田组雌性克氏原螯虾肝胰腺中PGE2含量均显著低于对照组(P<0.05,下同),其他生化指标与对照组雌性克氏原螯虾无显著差异;晒田组雌性克氏原螯虾血清中PGF2α和PGE2含量显著低于对照组雌性克氏原螯虾,其他生化指标与对照组雌性克氏原螯虾无显著差异。晒田胁迫3 d时,晒田组雌性克氏原螯虾卵巢中GSH-Px和PGF2α含量显著高于对照组雌性克氏原螯虾,其他生化指标与对照组雌性克氏原螯虾均无显著差异;晒田组雌性克氏原螯虾肝胰腺中ACP、GSH-Px、PGF2α和PGE2含量显著高于对照组雌性克氏原螯虾,AKP/ALP含量与对照组雌性克氏原螯虾无显著差异;晒田组雌性克氏原螯虾血清中AKP/ALP、ACP、GSH-Px、PGF2α和PGE2含量与对照组雌性克氏原螯虾均无显著差异。晒田胁迫7 d时,晒田组雌性克氏原螯虾卵巢和肝胰腺中AKP/ALP、ACP、GSH-Px、PGF2α和PGE2含量均显著高于对照组雌性克氏原螯虾;晒田组雌性克氏原螯虾血清中GSH-Px、PGF2α和PGE2含量显著高于对照组雌性克氏原螯虾,AKP/ALP和ACP含量与对照组雌性克氏原螯虾无显著差异。晒田胁迫10 d时,晒田组雌性克氏原螯虾卵巢、肝胰腺和血清中AKP/ALP、ACP、GSH-Px、PGF2α和PGE2含量均显著高于对照组雌性克氏原螯虾。
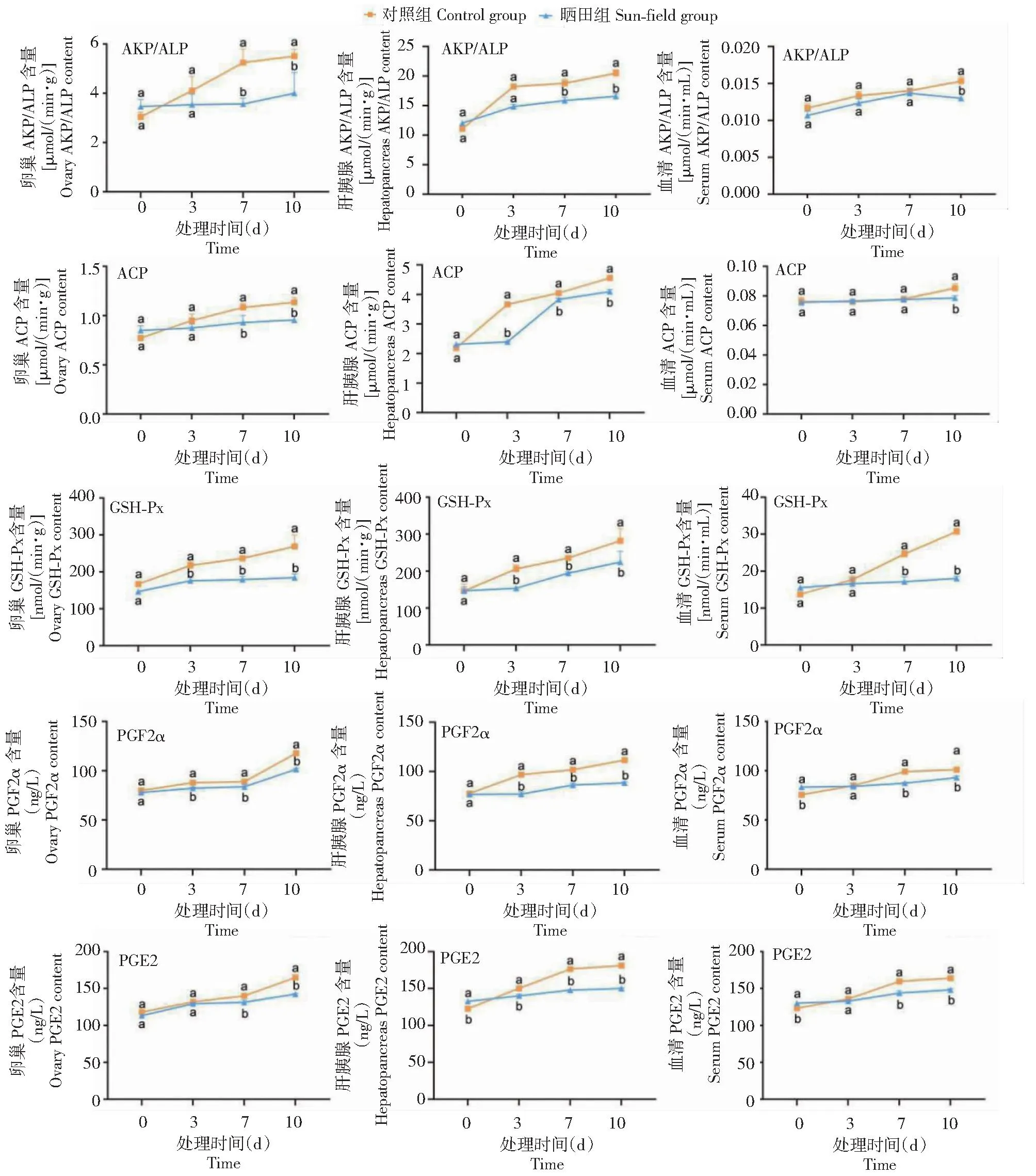
不同小写字母表示同一时间下不同处理组间差异显著(P<0.05)。下同。Different lowercase letters indicated significant difference between different treatment groups at the same time (P<0.05).The same as below.图1 晒田环境胁迫对克氏原螯虾生化酶活性的影响Fig.1 Effect of sun-field environmental stress on biochemical enzyme activity of P.clarkii
2.3 晒田环境胁迫对克氏原螯虾性类固醇激素的影响
如图2所示,晒田胁迫0 d时,晒田组雌性克氏原螯虾血清中T和PROG含量均显著高于对照组雌性克氏原螯虾,Estrone和17β-E2含量与对照组雌性克氏原螯虾无显著差异。晒田胁迫3 d时,晒田组雌性克氏原螯虾血清中17β-E2、T和PROG含量均显著高于对照组雌性克氏原螯虾,Estrone含量与对照组雌性克氏原螯虾无显著差异。晒田胁迫7和10 d时,晒田组雌性克氏原螯虾血清中Estrone、17β-E2、T和PROG含量均显著高于对照组雌性克氏原螯虾。
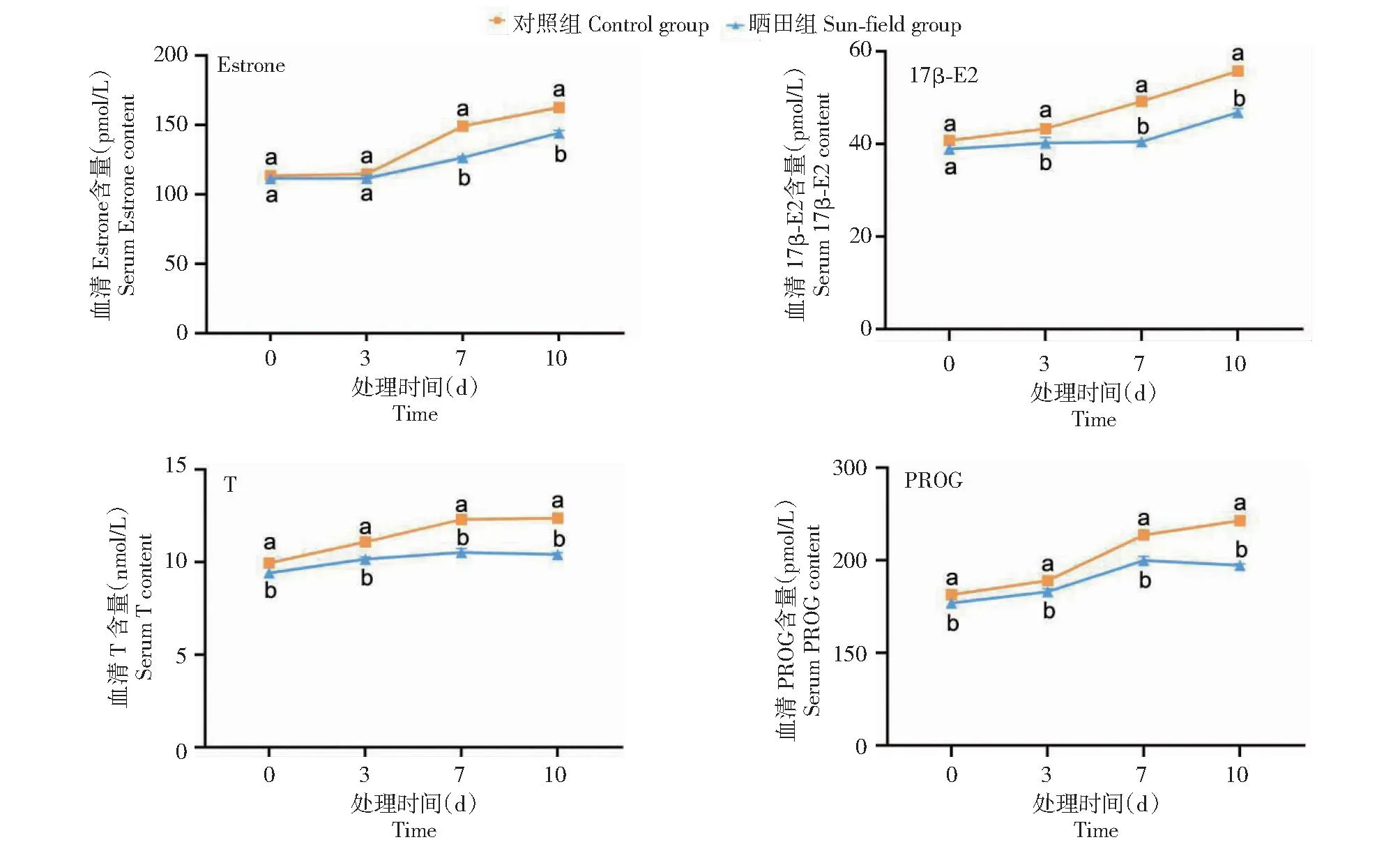
图2 晒田环境胁迫对克氏原螯虾性腺类固醇激素的影响Fig.2 Effect of sun-field environmental stress on gonadal steroid hormone of P.clarkii
2.4 晒田环境胁迫对克氏原螯虾高血糖家族激素的影响
如图3所示,晒田胁迫0 d时,晒田组雌性克氏原螯虾血清中CHH、GIH、MIH和MOIH含量均显著高于对照组雌性克氏原螯虾。晒田胁迫3 d时,晒田组雌性克氏原螯虾血清中CHH含量显著低于对照组雌性克氏原螯虾,MIH和MOIH含量显著高于对照组雌性克氏原螯虾,GIH含量与对照组雌性克氏原螯虾无显著差异。晒田胁迫7 d时,晒田组雌性克氏原螯虾血清中CHH和GIH含量显著低于对照组雌性克氏原螯虾,MOIH含量显著高于对照组雌性克氏原螯虾,MIH含量与对照组雌性克氏原螯虾无显著差异。晒田胁迫10 d时,晒田组雌性克氏原螯虾血清中CHH、MIH和MOIH含量均显著低于对照组雌性克氏原螯虾,GIH含量与对照组雌性克氏原螯虾无显著差异。
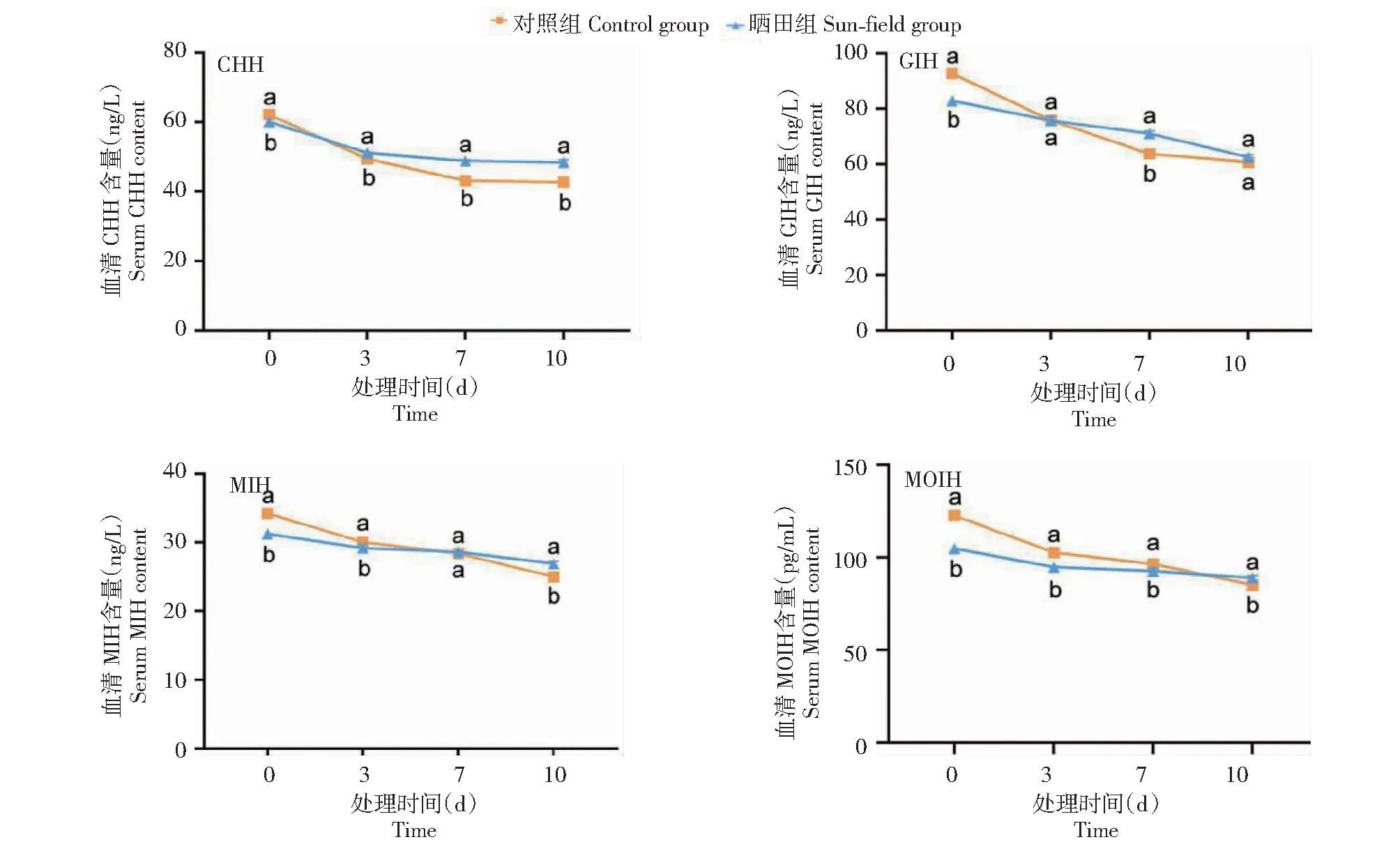
图3 晒田环境胁迫对克氏原螯虾高血糖家族激素的影响Fig.3 Effect of sun-field environmental stress on hyperglycemic family hormones of P.clarkii
2.5 晒田环境胁迫对克氏原螯虾其他卵巢发育相关激素的影响
如图4所示,晒田胁迫0 d时,晒田组雌性克氏原螯虾血清中Ecdysone含量显著高于对照组雌性克氏原螯虾,MF含量显著低于对照组雌性克氏原螯虾,Cortisol含量与对照组雌性克氏原螯虾无显著差异。晒田胁迫3 d时,晒田组雌性克氏原螯虾血清中Ecdysone和Cortisol含量显著高于对照组雌性克氏原螯虾,MF含量与对照组雌性克氏原螯虾无显著差异。晒田胁迫7和10 d时,晒田组雌性克氏原螯虾血清中Ecdysone、MF和Cortisol含量均显著高于对照组雌性克氏原螯虾。
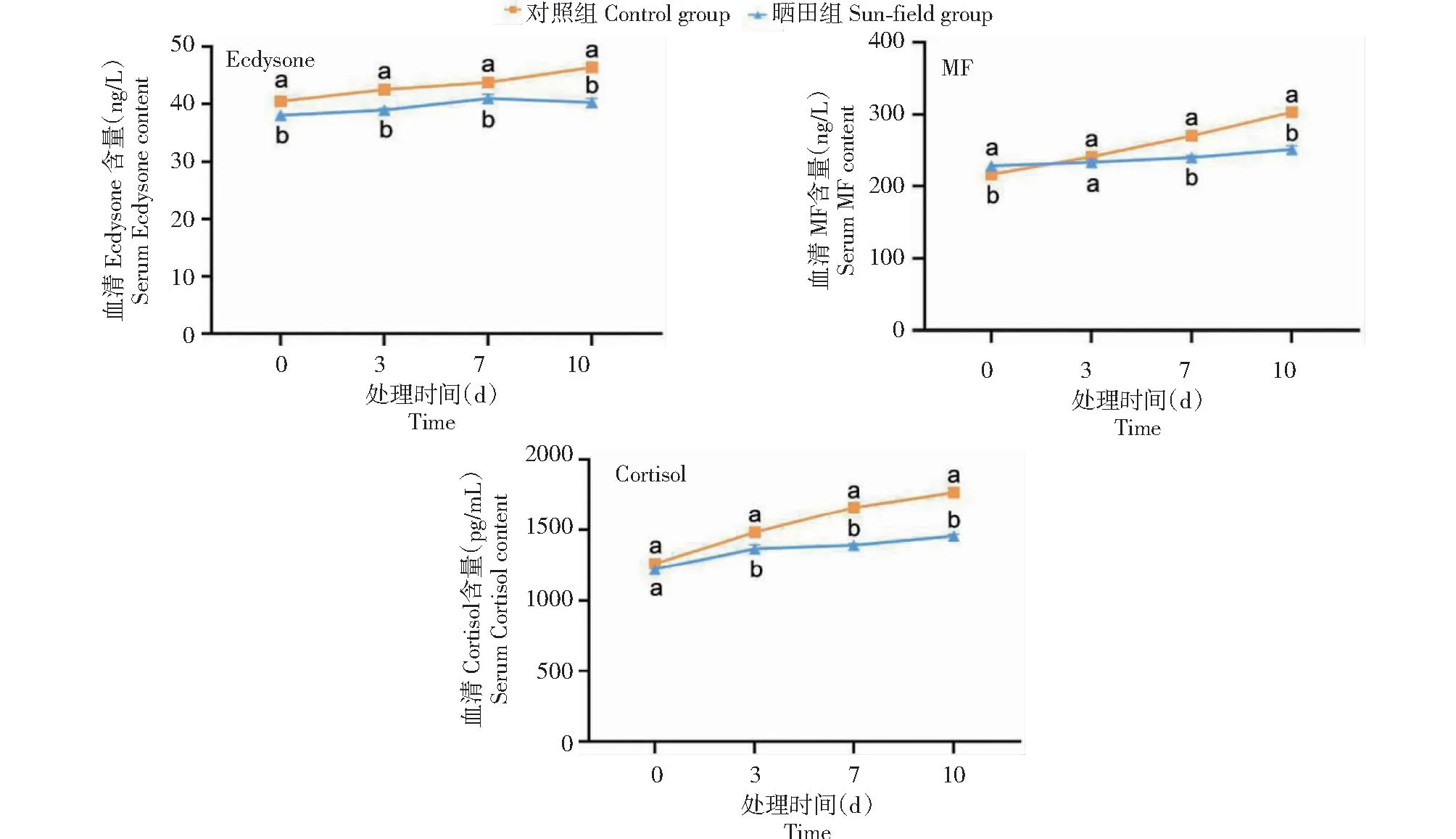
图4 晒田环境胁迫对克氏原螯虾其他卵巢发育相关激素的影响Fig.4 Effect of sun-field environmental stress on other ovarian development-related hormones of P.clarkii
3 讨 论
3.1 晒田环境胁迫对克氏原螯虾卵巢发育的影响
环境胁迫能影响生长、繁殖、储存等活动的能量最佳分配[11]。研究表明,晒田环境胁迫可诱导克氏原螯虾进行掘洞、交配和产卵,随胁迫时间增加能促进卵巢成熟并提高繁殖性能[12]。本研究中,晒田胁迫10 d时,晒田组雌性克氏原螯虾卵巢发育分期主要分布在IV期,卵巢成熟率为96.67%,对照组雌性克氏原螯虾的卵巢则主要分布在III期和IV期,卵巢成熟率为60.00%,表明晒田环境胁迫可在10 d内大幅度提高卵巢发育成熟速度,从而影响繁殖能力。推测晒田环境胁迫促进卵巢成熟与李玮等[3]研究报道的恶劣环境下克氏原螯虾体内营养能量将优先供给繁殖需求相关活动的特性有关。
3.2 克氏原螯虾生化指标与卵巢成熟度相关性及其作用分析
AKP/ALP、ACP、GSH-Px、PGF2α和PGE2在甲壳类动物卵巢发育过程中发挥重要作用。AKP/ALP和ACP为生物体内2种重要代谢调控酶,可催化磷酸单酯水解与磷酸基团代谢,对免疫性能具有重要作用[13-14];AKP/ALP和ACP参与甲壳动物钙、磷、蛋白质、脂质、DNA和RNA等代谢,对水产动物生长、能量供给、营养吸收和利用等意义重大[14-15]。GSH-Px是甲壳动物抗氧化防御系统中的关键酶,在克氏原螯虾免疫防御和卵子发生过程中发挥重要作用[16]。刀额新对虾(Metapenaeusensis)卵巢成熟过程中,GSH-Px可在防止卵母细胞氧化损伤及平衡活性氧生成方面发挥关键作用[17]。前列腺素类化合物的突出作用是调节甲壳动物雌性生殖系统成熟[18]。研究表明,PGF2α和PGE2可显著提高淡水蟹(Oziotelphusasenexsenex)和锯缘青蟹(S.serrata)卵巢指数、卵母细胞直径和数量、卵黄蛋白原水平、Ecdysone和MF水平[19-20]。斑节对虾PGF2α浓度含量与卵巢成熟呈负相关,PGE2则相反[18];日本囊对虾PGF2α和PGE2浓度含量与卵巢成熟呈负相关[21],究其原因可能是物种特异性差异所致。本研究中,晒田胁迫0 d时,晒田组雌性克氏原螯虾不同组织中AKP/ALP、ACP、GSH-Px、PGF2α和PGE2含量均显著低于对照组或与对照组雌性克氏原螯虾无显著差异;晒田胁迫3 和7 d时,晒田组雌性克氏原螯虾不同组织中这些生化指标含量均显著高于对照组或与对照组雌性克氏原螯虾无显著差异,而晒田胁迫10 d时,晒田组雌性克氏原螯虾不同组织中所有生化指标含量均显著高于对照组雌性克氏原螯虾。这一结果表明随时间增加,晒田组雌性克氏原螯虾不同组织的生化指标含量均呈上升趋势,且这些生化指标含量与晒田胁迫时间呈正相关,结合上述晒田胁迫促进克氏原螯虾卵巢成熟的结论,可证实5个生化指标浓度含量与卵巢成熟呈正相关。推测5个生化指标对晒田环境胁迫下克氏原螯虾卵巢成熟过程中的能量供给、营养吸收利用、免疫防御、卵母细胞生长及部分卵巢发育相关激素调节起重要作用。然而,本研究中晒田组与对照组雌性克氏原螯虾的部分生化指标与激素在晒田胁迫0 d时存在显著差异,推测为晒田前1周,试验雌虾经捕捞搬运后进入2个试验池塘的新环境中,在此过程中产生一定应激,直至晒田开始时体内的生化指标与激素还受到影响。
3.3 克氏原螯虾激素水平变化机理及其作用分析
性类固醇激素可调控甲壳动物卵黄发生和卵巢发育[22-23],对繁殖起关键作用,包括Estrone、17β-E2、T和PROG等。Estrone可促进鳗鲡卵巢发育,使其产生外观和颜色等第二性征变化[24]。17β-E2可刺激甲壳动物合成卵黄蛋白原,影响卵母细胞生长及卵黄发生[25]。T可诱导卵巢发育,在卵巢生长过程中基于睾酮芳构化成为雌二醇[26]。PROG作为一种前体,在甲壳动物卵巢细胞发育至特定阶段转化为其他类固醇激素,促进卵黄积累及卵巢发育[27]。在鱼类中,17β-HSD(17β羟类固醇脱氢酶)可将Estrone分解为17β-E2,P450芳化酶可将T分解为17β-E2[24],但在甲壳动物尚未发现相关报道。本研究中,晒田胁迫0和3 d时,晒田组雌性克氏原螯虾Estrone、17β-E2、T和PROG含量显著高于对照组或与对照组雌性克氏原螯虾无显著差异,晒田胁迫7和10 d时,晒田组雌性克氏原螯虾血清中的性类固醇激素均显著高于对照组雌性克氏原螯虾,表明晒田组雌性克氏原螯虾血清Estrone、17β-E2、T和PROG含量随时间增加均呈上升趋势。上述4个激素对卵巢发育均有促进作用,随时间增加,晒田组雌性克氏原螯虾血清中激素对卵巢发育的积极作用随之增强。
高血糖家族激素可调控甲壳类动物蜕皮、代谢、繁殖、渗透平衡等活动,主要成员为CHH、GIH、MIH和MOIH[28]。CHH可调节甲壳动物糖代谢和脂代谢,抑制MF分泌及卵黄蛋白原合成,从而抑制卵巢发育[29-30]。但王晓伟等[31]认为,CHH能刺激卵黄发生,促进生殖。GIH浓度随着美洲龙虾生殖周期成熟而降低,可抑制卵黄发生,从而抑制卵巢发育[32]。MIH可诱导蓝蟹(Callinectessapidus)和刀额新对虾(Metapenaeusensis)卵巢成熟,增加卵黄蛋白原表达[31,33]。 赵真慧[34]研究认为MIH抑制脊尾白虾卵巢发育和Ecdysone合成,降低MIH生成与释放,可促进卵巢成熟。MOIH可抑制甲壳类动物大颚器官分泌MF,降低MF分泌速度,抑制卵黄发生及卵巢发育[35]。本研究中,晒田胁迫0 d时,晒田组雌性克氏原螯虾CHH、GIH、MIH和MOIH含量均显著高于对照组雌性克氏原螯虾,晒田胁迫3和7 d时,晒田组雌性克氏原螯虾中高血糖家族激素含量显著高于、低于对照组或与对照组雌性克氏原螯虾无显著差异,晒田胁迫10 d时,晒田组雌性克氏原螯虾中高血糖家族激素含量显著低于对照组或与对照组雌性克氏原螯虾无显著差异,表明晒田组雌性克氏原螯虾CHH、GIH、MIH和MOIH含量随时间增加均呈下降趋势,高血糖家族激素含量与晒田胁迫时间均呈负相关,上述4个激素均能抑制卵巢发育。因此,随着胁迫时间增加,激素含量降低,晒田组雌性克氏原螯虾卵巢发育加快。
甲壳类动物卵巢发育涉及多种激素。Cortisol为应激激素,具有调控糖、脂肪生物合成的作用,当动物受环境胁迫可使机体产生自我保护,体内能量失衡将刺激性腺分泌Cortisol[8,36]。本研究中,晒田胁迫0 d时,晒田组雌性克氏原螯虾Cortisol含量与对照组雌性克氏原螯虾不存在显著差异,而晒田胁迫3、7和10 d时,晒田组雌性克氏原螯虾Cortisol含量显著高于对照组雌性克氏原螯虾,推测为晒田组克氏原螯虾因水量骤变以及高温暴晒产生的应激反应,体内能量失衡促使卵巢分泌Cortisol进行自我保护。Ecdysone主要调控甲壳和节肢动物生长发育、性腺发育和形态发生,对虾蟹类卵黄生成、卵巢成熟和蛋白合成具有重要作用[31,37]。本研究中,晒田胁迫0、3、7和10 d时,晒田组雌性克氏原螯虾Ecdysone含量均显著高于对照组雌性克氏原螯虾,表明晒田组克氏原螯虾在试验期间全程分泌Ecdysone,以促进卵巢发育。MF可调控甲壳类动物卵巢发育,刺激Ecdysone分泌,对卵母细胞具有促进作用,已在克氏原螯虾、锐脊单肢虾(Sicyoniaingentis)和拟穴青蟹(S.paramamosain)等甲壳类动物中得到证实[38-39]。本研究中,晒田胁迫0 d时,晒田组雌性克氏原螯虾MF含量显著低于对照组雌性克氏原螯虾,晒田胁迫3 d时,则与对照组雌性克氏原螯虾无显著差异,晒田胁迫7和10 d时,晒田组雌性克氏原螯虾MF含量显著高于对照组雌性克氏原螯虾,表明晒田组雌性克氏原螯虾MF含量随时间增加呈上升趋势,雌虾体内大量分泌MF,同时刺激Ecdysone分泌,二者共同调控卵巢发育。
4 结 论
晒田环境胁迫能促进克氏原螯虾卵巢成熟,加快卵巢发育相关生化指标和激素的变化。晒田胁迫7~10 d时,促进卵巢发育成熟的效果达到峰值,表现为卵巢成熟率接近100.00%。在此过程中,AKP/ALP、ACP、GSH-Px、PGF2α、PGE2含量与卵巢成熟度呈正相关;性类固醇激素(Estrone、17β-E2、T、PROG)和卵巢发育相关激素(Ecdyone、MF、Cortisol)含量升高,高血糖家族激素(CHH、GIH、MIH、MOIH)含量降低,上述激素联合调控对卵巢发育具有积极作用。
