土壤微生物群落结构与功能对咖啡果皮覆盖的响应
2023-12-29张尧榆卢志青李丽华董云萍林兴军赵青云龙宇宙
张 昂,张尧榆,卢志青,李丽华,董云萍,林兴军,赵青云,孙 燕,谭 军,龙宇宙
(1. 中国热带农业科学院香料饮料研究所/海南省热带香辛饮料作物遗传改良与品质调控重点实验室/农业农村部香辛饮料作物遗传资源利用重点实验室,海南 万宁 571533;2. 云南省董云萍专家工作站, 云南 普洱 665099;3. 云南省闫林专家工作站, 云南 普洱 665099;4. 云南农业大学热带作物学院,云南 普洱 665000)
【研究意义】咖啡是世界三大饮料之一,经济价值较大。咖啡在我国种植面积超过1.2×105hm2,生豆产量达1.7×105t,其产区主要集中于云南、海南、广东和广西等热带亚热带地区,也是我国热带地区重要的经济作物。因此,大力推广特色咖啡产业、提高咖啡种植收益,已成为推动农民致富、乡村振兴的主要途径之一[1]。咖啡属于浅根系作物,其根系分布在10~30 cm土层中[2],传统上以覆盖栽培为主要生产模式[3],一般使用椰糠、秸秆等农林废弃物进行覆盖栽培,用以保持水土、改善土壤微环境、维护根系健康[4]。化肥农药在土壤中的累积严重威胁咖啡植株健康,加剧咖啡园早衰减产现象。不仅如此,咖啡生果附加值低、咖啡副产物大量损失还造成环境污染与大量资源浪费。因此,采用咖啡凋落物与果皮代替传统覆盖物进行生态循环栽培,既减少咖啡生产过程中化肥和除草剂等投入品用量、降低咖啡栽培劳动强度与生产成本,又推动咖啡废弃物资源的循环利用,提高咖啡的综合利用率,成为提高咖啡种植效益和相关产业可持续发展的重要举措。【前人研究进展】土壤微生物作为土壤生态系统的重要组成部分,不仅调节土壤养分循环和物质交换等过程,还影响动植物的生长发育以及作物的生产力,进而改变农田生态系统多样性与稳定性[5-6],成为表征土壤健康的重要指标之一。研究表明,种植模式变化主要通过改变农田微环境或影响不同生物间互作关系等途径直接或间接调控土壤中微生物群落[7]。例如,覆盖栽培不仅能够通过提高土壤水分含量、促进土壤养分累积、增强土壤生物活性等途径促进土壤微生物多样性[8],还能够通过加速有益菌之间的协同演替,进而改善土壤微生物群落结构[9-10]。覆盖栽培同样可能对土壤微生物群落产生消极作用,研究表明农业废弃物覆盖通过加速土壤致病菌群落繁殖,导致土壤微生物群落失衡,加剧土传病害的发生[11]。因此,不同咖啡废弃物覆盖对咖啡园土壤微生物的影响仍需进一步探究。土壤微生物群落结构和功能的变化能够影响作物的生长,然而作物新陈代谢或死亡分解产生的有机物、蛋白质和多糖又对土壤的理化性质、微生物群落结构和功能产生反馈作用[12]。因此伴随咖啡废弃物的分解过程,土壤微生物群落结构和功能存在较大差异。首先,咖啡果皮与枯落物自身性质(养分含量、机械强度等指标)的互异,导致其主要的分解微生物类群存在显著差异[13]。其次,咖啡果皮和枯落物的分解产物可能具有一定的化感作用,不同土壤微生物群落对不同咖啡废弃物分解产物的响应机制可能存在显著差异[14]。再次,咖啡废弃物的分解过程主要有环境因子调控的前期快速失重阶段以及由土壤微生物驱动的后期长时间裂解阶段[15]。在不同的分解阶段,土壤微生物群落演替会受到咖啡果皮和枯落物残量的影响[16],导致土壤微生物群落结构与功能在咖啡废弃物的不同分解阶段,随着有机残体的演变而发生变化[17-18]。此外,咖啡废弃物的覆盖同样能够通过改变土壤微环境进而调控土壤微生物群落结构[17, 19]。前人研究表明,咖啡园中覆盖咖啡废弃物能够显著提高土壤碱解氮和速效钾含量,对提升土壤微生物群落多样性具有积极作用,但过量的果皮施用会导致土壤酸化,对土壤微生物群落多样性和结构稳定产生消极作用[19]。【本研究切入点】目前仍尚不清晰不同废弃物对土壤微生物群落的影响,咖啡土壤微生物对咖啡果皮和枯落物覆盖的响应机制仍需进一步探究。【拟解决的关键问题】本研究通过监测不同咖啡废弃物覆盖下微生物群落结构与功能差异,明确土壤理化性质及微环境与微生物群落结构及功能的内在关系,阐明咖啡废弃物分解驱动土壤微生物群落结构变化的关键调控因子,为咖啡园土壤健康保育技术开发提供理论基础。
1 材料与方法
1.1 实验地概况
实验样地位于海南省东部万宁市兴隆热带植物园(110°11′ E,18°44′ N,海拔 36 m)。该地位于南亚热带与热带气候过渡带,属热带季风气候,雨热资源充足,年降水量2100 mm,年日照时长大于1750 h。该地区土壤分类主要为砖红壤,土壤pH 6.15,有机质含量22.06 g/kg,土壤总氮含量1.49 g/kg,土壤总磷含量1.28 g/kg,土壤全钾含量6.29 g/kg。
1.2 实验设计
实验采取随机区组设计,共设置未覆盖咖啡废弃物(C)、覆盖咖啡枯落物(L)、覆盖咖啡果皮(P)和同时覆盖咖啡枯落物及果皮(PL)4种覆盖处理。前期通过随机调查海南咖啡种植园现存咖啡枯落物和果皮累积量,计算得到咖啡枯落物平均累积量为301.39 g/m2,果皮的平均累积量为239.05 g/m2,本试验按照平均累积量对咖啡废弃物进行等量添加,具体添加量根据树盘面积(咖啡植株垂直投影面积)进行换算。咖啡果皮覆盖土壤表面,不进行翻耕等处理。每个覆盖处理重复4次。所有咖啡植株采用相同的水肥处理,即统一按照传统咖啡植株生产模式进行浇水和施肥。
1.3 实验材料
本研究采用中粒种咖啡热研1号(CoffeacanephoraPierre ex A. Froehner cv. Reyan No.1)。咖啡枯落物收集于中国热带农业科学院香料饮料研究所咖啡种质资源圃,咖啡果皮收集于海南省兴隆华侨农场太阳河咖啡加工厂,枯落物与果皮来源于与本研究相同的品种。
1.4 实验方法
1.4.1 取样方法 于2021年9月对土样进行收集,采用3点法进行采样,以土钻法将同一样点的3个土钻样品混合获得0~20 cm耕层土壤样品,其中一部分混合样品在实验室内风干后过1 mm筛,用于测定土壤理化性质,另一部分于混合取样后立即保存于-80 ℃超低温冰箱,用于土壤微生物丰度和多样性测定。
1.4.2 土壤理化性质测定方法 土壤含水量(Soil moisture content,SM)和容重(Bulk density,BD)测定固定体积下土壤鲜重和干重,土壤含水量=(土壤干重/土壤鲜重)×100%,土壤容重=土壤干重/土壤体积。土壤pH使用FE28型pH计测定,有机质(Soil organic matter,SOM)采用总有机碳分析仪(Multi N/C 3100)测定,碱解氮(Alkali hydrolyzed nitrogen, SAN)采用碱解扩散法测定,速效磷(Soil available phosphorus, SAP)采用紫外分光光度计法测定, 速效钾(Soil available potassium, SAK)采用火焰光度计法测定。具体测定方法参照鲍士旦[20]主编的《土壤农化分析》(第三版)。
1.4.3 土壤微生物测定方法 土壤总DNA采用DNA提取试剂盒提取和纯化[20]。具体方法是利用标记有barcode的引物序列(细菌:967F/1046R;真菌:ITS1F12/ITS2)扩增相应土壤细菌 16SrRNAV3-4区片段和真菌 ITS-1 序列片段,以2%琼脂糖凝胶电泳检测产物片段长度,根据定量检测结果,将扩增产物混合为1个样本,构建克隆文库;利用Illumina MiSeq高通量平台测序,数据经Flash软件双端序列拼接、质控、去接头后获得优化序列,基于优化序列进行 OTU聚类,获得 OTU丰度表,用于后续细菌多样性指数及细菌群落构成分析。FAPROTAX(v1.2.1)和FUNGuild(v1.0)软件分别用于预测和分析具有不同功能的土壤细菌和真菌群落的相对丰度[21]。
1.5 数据分析
采用SAS v8软件进行双因素方差分析,以枯落物覆盖和果皮覆盖为2个固定因素,区组为随机因素,比较土壤理化性质指标和细菌丰度、细菌丰富度、细菌多样性指数、真菌丰度、真菌丰富度、真菌多样性指数等土壤微生物群落指标在不同处理下是否存在显著差异[22]。采用SPSS 23.0软件进行线性相关分析,用于计算不同土壤指标与土壤微生物群落之间的相关关系[23];使用CANOCO 5.0软件进行典型相关分析(CCA),用于明确影响土壤微生物群落结构的主要环境因子,分析过程中选择 manual forward selection 程序使用具有499个排列的蒙特卡罗测试,以确定环境变量参数的显著性[23-24]。Spearman的等级相关性分析用于测试土壤微生物优势群落与土壤理化性质之间的关联度[23-24]。使用Origin 9.1和R 4.0.3进行数据绘图。
2 结果与分析
2.1 不同咖啡废弃物覆盖对土壤理化性质及微环境的影响
双因素方差分析结果(表1)表明咖啡果皮与枯落物覆盖处理不影响土壤pH、温度、水分含量、有机质含量和速效磷含量,且两种处理对上述指标的影响不存在交互效应。咖啡果皮覆盖显著提高土壤速效钾含量和土壤碱解氮含量(P<0.05,下同),但咖啡枯落物覆盖不影响上述2个指标,且两种处理对上述指标的影响不存在交互效应(表1~2)。

表1 不同覆盖模式对土壤微环境及理化性质的双因素方差分析结果(F值)Table 1 Results of two-way ANOVA on soil micro-climate, physical and chemical properties under different mulching patterns (F value)
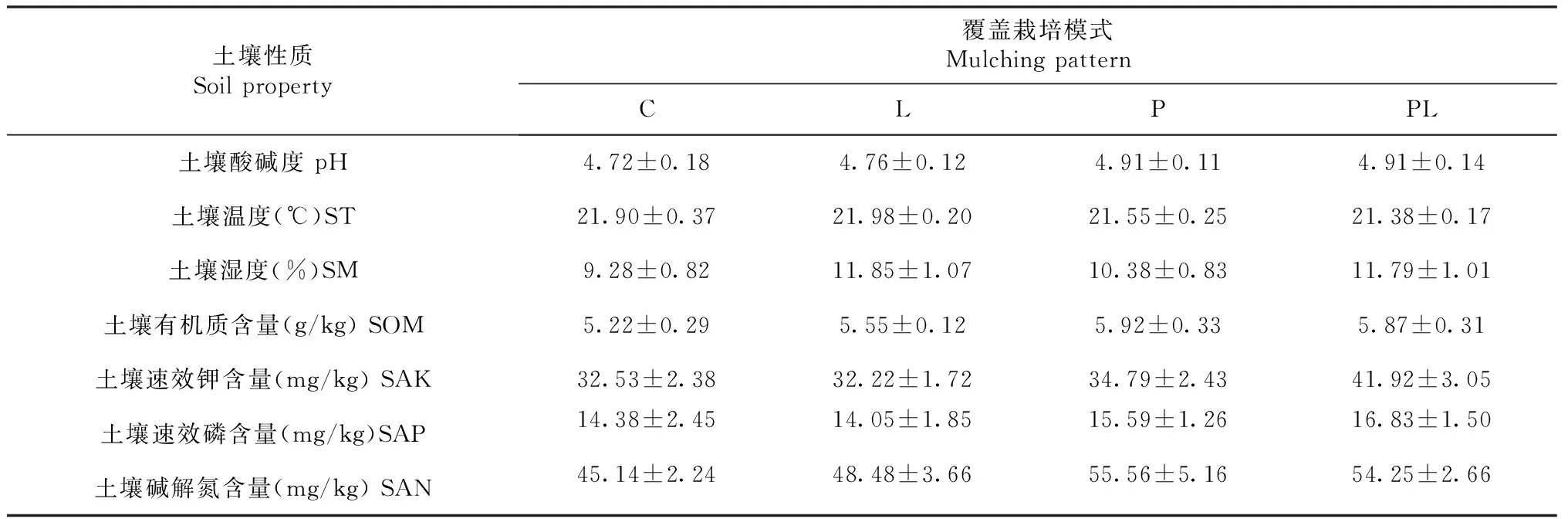
表2 不同咖啡废弃物覆盖模式下土壤理化性质Table 2 Soil physicochemical properties under different mulching patterns
2.2 不同咖啡废弃物覆盖对土壤微生物群落多样性的影响
咖啡果皮与枯落物覆盖处理不影响细菌群落丰度、真菌群落丰度、真菌丰富度和多样性,且两种处理对上述指标的影响不存在交互效应(表3)。咖啡果皮显著提高细菌群落丰富度和细菌群落多样性,但咖啡枯落物盖不影响上述2个指标,且两种覆盖处理对上述指标的影响不存在交互效应(表3,图1)。表明,相比于真菌群落,细菌群落对咖啡废弃物覆盖的响应更加敏感,且果皮覆盖对细菌群落丰富度和多样性的促进作用更加显著。
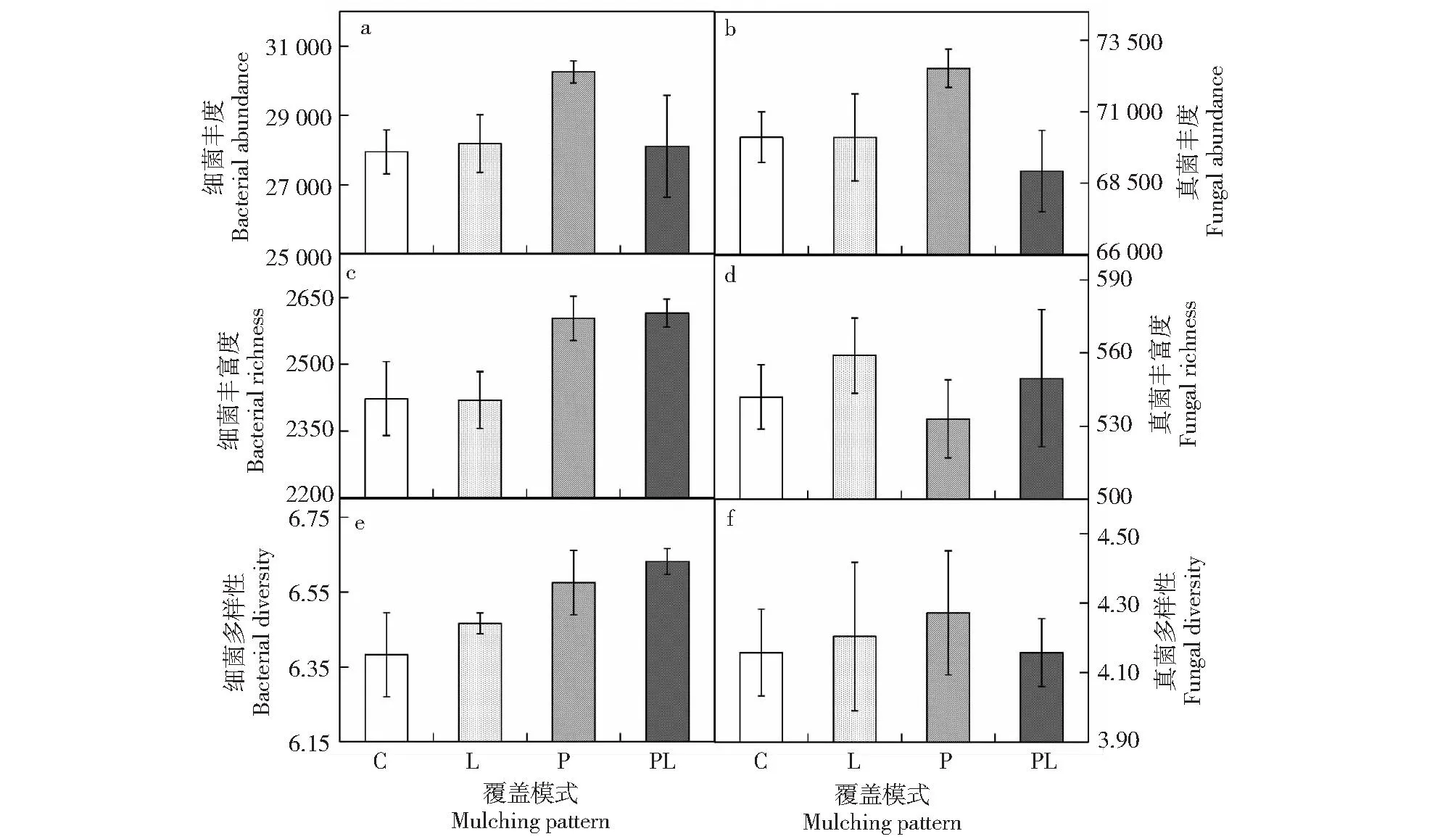
误差棒表示标准误(SE)。Error bar represents SE.图1 不同咖啡废弃物覆盖模式下土壤微生物群落丰度、丰富度和多样性指数Fig.1 Soil microbial community abundance, richness and diversity under different mulching patterns

表3 不同覆盖模式对土壤微生物群落多样性与结构的双因素方差分析结果(F值)Table 3 Results of two-way ANOVA on soil microbial community diversity and structure under different mulching patterns (F value)
2.3 不同咖啡废弃物覆盖对土壤微生物群落组成与结构的影响
一般将相对丰度大于1%的菌门视为优势菌门。本研究中变形菌门、放线菌门、酸杆菌门、绿弯菌门、厚壁菌门、芽单胞菌门、粘球菌门、Methylomirabilota、拟杆菌门、蓝细菌门和疣微菌门是细菌群落中的优势菌门。不同咖啡废弃物覆盖下细菌群落放线菌门、酸杆菌门、绿弯菌门、芽单胞菌门、厚壁菌门、粘球菌门、Methylomirabilota、拟杆菌门和疣微菌门丰度并未存在显著差异。果皮覆盖显著提高变形菌门丰度以及厚壁菌门丰度,却显著降低蓝细菌门丰度。咖啡枯落物覆盖显著提高厚壁菌门丰度,却显著降低蓝细菌门丰度,并且两种废弃物覆盖仅对放线菌门、厚壁菌门、芽单胞菌门和蓝细菌门的影响存在交互作用(图2)。

图2 不同咖啡废弃物覆盖模式下土壤微生物优势菌门相对丰度Fig.2 Relative abundance of dominant microbe in soil under different mulching patterns
本研究中子囊菌门、担子菌门、壶菌门、罗兹菌门、毛霉菌门和球囊菌门是真菌群落中的优势菌门。咖啡果皮覆盖对所有真菌优势菌门均无影响,但咖啡枯落物覆盖却显著提高子囊菌门丰度,并且显著降低放线菌门丰度。两种废弃物覆盖仅对子囊菌门的影响存在交互作用(图2)。
2.4 不同咖啡废弃物覆盖对土壤微生物群落功能的影响
由表4可知,对细菌功能而言,咖啡枯落物覆盖显著促进化学自养氮循环相关功能细菌多度增加,却显著减少光自养细菌相对丰度和有机质降解相关细菌相对丰度。咖啡果皮覆盖同样分别显著提高化能自养和氮循环细菌的相对丰度,却显著减少光自养相关功能细菌的相对丰度。对真菌功能而言,咖啡枯落物覆盖显著降低菌根真菌的相对丰度。咖啡果皮覆盖处理在显著提高菌根真菌的相对丰度的同时却显著降低病原体真菌的相对丰度。

表4 不同咖啡废弃物覆盖模式下土壤微生物群落功能Table 4 Soil microbial functions under different mulching patterns
2.5 土壤非生物因子与土壤微生物多样性和结构的相关性分析
土壤理化性质与微生物丰度、丰富度和多样性的相关性分析(图3)表明,细菌丰度与土壤湿度(R=0.54,P<0.05)呈显著负相关。细菌丰富度(R=0.55,P<0.05)、多样性(R=0.50,P<0.05)与土壤速效钾含量呈显著正相关。真菌群落丰度、丰富度和多样性对土壤非生物因子变化的响应不敏感。
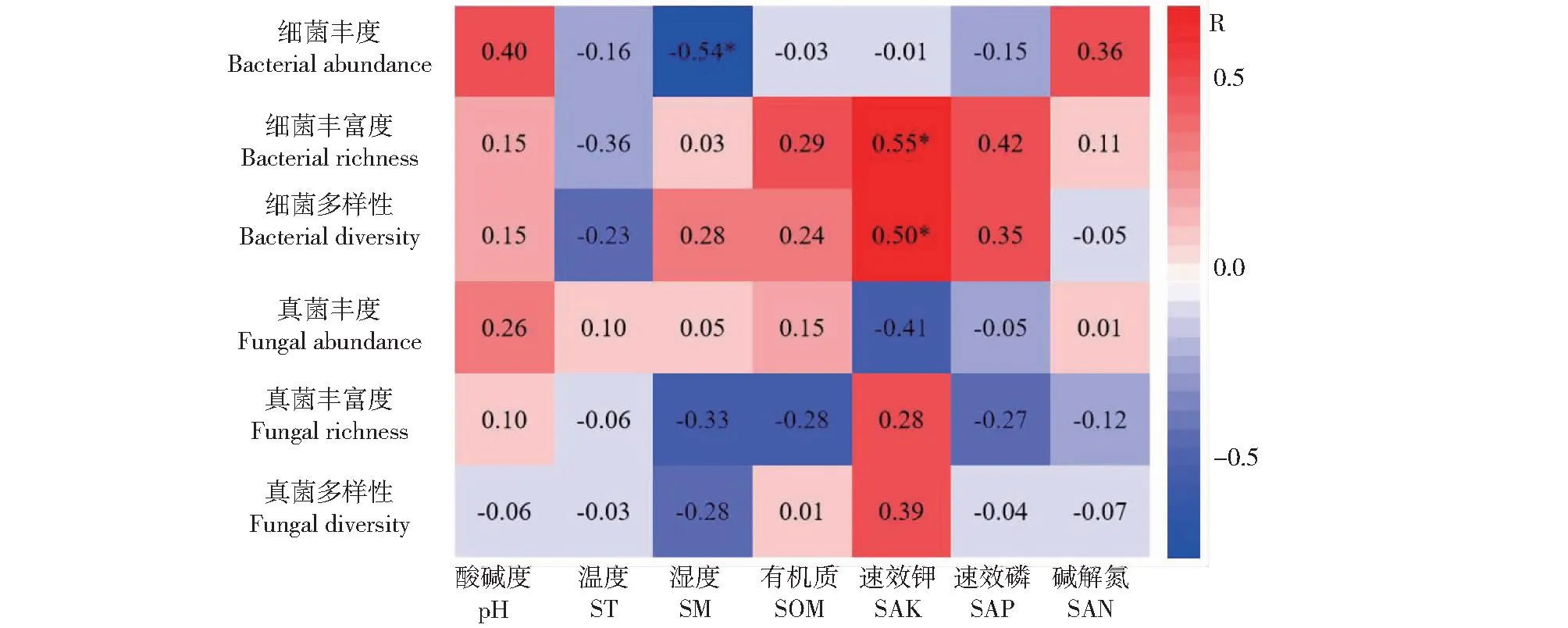
“*”表示P<0.05, “**”表示P<0.01。‘*’ represents P<0.05, and ‘**’ represents P<0.01.图3 不同咖啡废弃物覆盖下咖啡微生物群落指标与环境因子相关性分析Fig.3 Correlation analysis of soil microbial community index and environmental factor under different mulching patterns
采用典型相关分析(CCA)进一步分析各土壤非生物因子与微生物群落结构之间的关系。影响细菌群落优势菌门最大的土壤理化指标为SAK(F=3.20,P=0.03)和SOM(F=3.00,P=0.03,图4-a)。所有的环境变量共同解释了样本间细菌群落变异的53.41%,影响顺序为 SAK>SOM>SM>pH>SAN>SAP>ST。CCA的前2个排序轴分别解释总方差的44.73%和8.68%。影响真菌群落优势菌门的环境变量顺序为SOM>SM>SAK>SAN>pH>ST>SAP(图4-b)。所有环境变量共同解释样本间真菌群落变异的36.62%,CCA的前2个排序轴分别解释总方差的25.08%和11.54%。

图中a为细菌,b为真菌。“Prote”代表变形杆菌门,“Acti”代表放线菌门,“Acid”代表酸杆菌门,“Chlo”代表绿弯菌门,“Firm”代表厚壁菌门,“Gemm”代表芽单胞菌门,“ Myxo”代表粘球菌门,“Meth”代表Methylomirabilota,“Bact”代表拟杆菌门,“Cyan”代表蓝细菌门,“Verr”代表疣微菌门,“Asco”代表子囊菌门,“Basi”代表担子菌门,“Chyt”代表壶菌门,“Roze”代表罗兹菌门,“Mort”代表毛霉菌门,“Glom”代表球囊菌门。In the figure, a is bacteria, and b is fungi. ‘Prote’ represents Proteobacteria,‘Acti’ represents Actinobacteriota,‘Acid’ represents Acidobacteriota,‘Chlo’ represents Chloroflexi,‘Firm’ represents Firmicutes,‘Gemm’ represents Gemmatimonadota, ‘Myxo’ represents Myxococcota,‘Meth’ represents Methylomirabilota,‘Bact’ represents Bacteroidota,‘Cyan’ represents Cyanobacteria,‘Verr’ represents Verrucomicrobiota,‘Asco’ represents Ascomycota,‘Basi’ represents Basidiomycota,‘Chyt’ represents Chytridiomycota,‘Roze’ represents Rozellomycota,‘Mort’ represents Mortierellomycota and ‘Glom’ represents Glomeromycota.图4 土壤微生物群落优势菌门与环境因子的典型分析(CCA)Fig.4 Canonical correlation analysis (CCA) of dominant soil microbial community phyla and environmental factors
3 讨 论
3.1 土壤理化性质对不同咖啡废弃物覆盖的响应
在不同的农田生态系统中,覆盖栽培模式对土壤理化性质的影响存在较大差异。其差异主要与生态系统微环境变化和覆盖物的分解特性密切相关[25]。而在同一农田生态系统中,不同的覆盖材料对土壤理化性质的影响主要归因于分解特性的差异和对土壤微环境的调控[26]。本研究中,咖啡果皮覆盖显著提高土壤速效钾和碱解氮含量,主要归因于咖啡果皮中氮和钾含量较高[1]。而枯落物对土壤理化性质的中性效应可能与咖啡叶片较低的碳氮比有关。前期研究表明,具有较高氮含量的叶片在分解初期具有较快的分解速率[27]。在本试验采样期间,咖啡枯落物已进入分解后期,其纤维素和木质素分解较慢,因此对土壤养分含量的影响较小。一般认为农林废弃物覆盖能够通过调控土壤水分和降低土壤温度进而影响土壤养分含量及理化性质,但在本研究中,土壤温湿度和pH并非调控不同咖啡废弃物覆盖下土壤性质的主要因子。
3.2 不同咖啡废弃物覆盖对土壤微生物群落多样性的影响
土壤微生物群落是农田生态系统重要的组成部分[28],主要参与土壤中有机质的分解、腐殖质的形成和养分循环等生态过程[29]。土壤养分含量在微生物群落结构装配过程中同样具有重要作用,是主导微生物群落演替以及维持土壤微生物多样性的重要因子[30]。一般认为,覆盖作物能够通过增加分解底物显著促进部分功能微生物基因丰度,提高细菌群落多样性[8, 31]。本试验中咖啡果皮覆盖通过加速钾和氮在土壤中的累积,显著提高细菌丰度以及多样性,支持上述论点。尽管土壤温度、容重、含水量、孔隙度同样对土壤细菌的多样性具有较大影响[32],但在本研究中细菌多样性并不响应于土壤温湿度以及pH的变化,表明在不同咖啡废弃物覆盖下,上述土壤性质及微环境并非是驱动细菌演替的主要因子。此外,一般认为细菌和真菌群落对土壤性质等环境因子的响应存在显著差异[33]。本研究中土壤真菌群落多样性对咖啡废弃物覆盖不敏感,与前期研究结果保持一致。
3.3 不同咖啡废弃物覆盖对土壤微生物群落结构与功能的影响
微生物组成与结构变化是废弃物覆盖条件下生态系统特性和环境因素综合作用的结果,其中凋落物质量是重要影响因素之一[34]。此外,土壤性质与水热条件差异也是引起微生物组成与结构的重要影响因素之一[35]。前人研究表明,细菌中最庞大的菌门是变形菌门细菌,具有广泛的生理代谢功能,能够适应多种复杂的环境[36-37]。变形菌门细菌作为嗜养菌,富含碳的环境中能够刺激其快速增长[38],因此咖啡果皮覆盖通过为变形菌提供充足的代谢底物,促进其生长和繁殖。厚壁菌门细菌多为化能异养型细菌,能够促进土壤有机质分解,其丰度与土壤速效磷、速效钾和碱解氮呈正相关[23],因此本研究中两种覆盖栽培通过增加土壤速效养分含量进而提高了厚壁菌门细菌丰度。蓝细菌门细菌是一类通过产氧光合作用获取能量的细菌,但其中也有部分是异养型细菌咖啡废弃物的覆盖通过增加土壤异养代谢底物显著增强异养型微生物竞争优势,引起蓝细菌门的相对比例显著下降[39]。
本研究中土壤真菌群落除子囊菌门、担子菌门外对咖啡废弃物覆盖的响应不敏感。子囊菌门真菌的主要功能是加速纤维素或木质素的分解[40],因此,在咖啡枯落物分解后期剩余纤维素含量较高的叶柄和枯枝等残体,能够引起子囊菌门相对比例的提高。担子菌门的功能较复杂,其中部分是作物致病菌[41]。因此,咖啡枯落物覆盖后降低担子菌门相对丰度可能对维持咖啡能作物健康有潜在的积极作用。
4 结 论
本研究中咖啡果皮与枯落物覆盖均不影响土壤真菌群落,并且细菌群落丰富度与多样性对咖啡枯落物覆盖的响应不敏感。咖啡果皮覆盖显著提高土壤速效钾和碱解氮含量以及促进土壤细菌群落丰富度与多样性,并提高变形杆菌门和厚壁菌门细菌以及降低蓝细菌门细菌相对丰度,但土壤微生物主要功能未发生较大改变。咖啡果皮的养分分解与迁移直接驱动土壤微生物群落多样性的提高以及维持结构与功能的相对稳定。咖啡果皮覆盖栽培有助于咖啡园土壤健康保育以及咖啡种植产业的可持续发展。
