不同施肥方式对杉木容器育苗基质真菌群落的影响
2023-12-25马东旭王佳琪蓝伟立陈块明叶义全范福金林开敏
马东旭,王佳琪,蓝伟立,陈块明,李 茂,叶义全,范福金,林开敏
(1. 福建农林大学 林学院,福建 福州 350000;2. 国家林业和草原局 杉木工程技术研究中心,福建 福州 350000;3. 福建省洋口国有林场,福建 南平 353000)
土壤真菌是土壤微生物群落中重要的分解者,能够分泌相应酶分解有机物为植物生长提供有效养分[1]。由于土壤真菌群落结构对环境因素变化较为敏感,因此将其作为评价土壤质量状况的重要指标[2]。相关研究表明土壤真菌群落结构与多样性主要受不同管理措施、土壤肥力、土壤类型、栽培植物类型、温度、水分等综合因素有关[3-4]。通过调控土壤真菌群落结构与多样性有利于改善土壤肥力、增加土壤养分有效性,提高土壤生产力[8]。
杉木Cunninghamialanceolata作为我国南方主要造林树种[5],兼具速生丰产与材质优良的特性。杉木幼苗时期生长迅速,对养分需求较高,而土壤真菌群落对土壤营养元素的周转有重要调节作用,有利于加快养分循环,可为杉木幼苗生长提供有利条件。
近年来,人们对木材需求以及质量要求日益增长,对优质良种苗木的需求也不断增加。施肥作为提高苗木质量的主要措施,在改善土壤肥力,促进苗木生长的同时,对土壤微生物的群落结构与多样性具有重要调控作用[6-7]。因此,合理施肥能维持土壤稳定的真菌群落结构与多样性,对改善土壤肥力和为杉木幼苗生长提供养分有重大意义。
目前通过高通量测序技术开展施肥对土壤真菌群落结构的研究已有许多报道,但这些研究多集中于农田作物栽培中,如茶树[8]、冬小麦[9]、番茄[10]等,并取得了较好研究成果。有关木本植物的研究则相对较少,有关杉木幼苗土壤真菌群落和功能多样性[11-12]已有相关报道,有关不同施肥对杉木容器育苗基质真菌群落结构影响的研究却鲜有报道。有鉴于此,以栽培杉木无性系幼苗的轻型基质为对象,基于高通量测序技术,对不同施肥方式下轻型基质真菌群落结构组成和多样性进行系统分析,比较不同施肥处理下的基质真菌群落结构差异,从微生物角度来研究不同施肥方式对杉木容器育苗基质真菌群落结构和多样性的影响及其与环境因子之间的关系,为施肥调控杉木容器育苗基质真菌群落提供理论参考。
1 材料与方法
1.1 试验材料
由福建洋口林场采穗圃提供生长势一致的“洋061”优良无性系杉木穗条,长度为9.0±0.5 cm。2018 年4 月初采用无纺布袋(直径5.5 cm×高9.5 cm),将轻型基质(泥炭土∶珍珠岩∶杉木皮=1∶2∶2(体积比))装入其中,并置于塑料托盘上。将装好轻型基质的托盘放置于平整苗床之上,在苗床上方铺一层黑色遮阳网,扦插前对基质进行浇水以保持湿润。轻型基质初始理化性质如下:容重0.17 g/cm3,总孔隙度58.8%,含水率29.2%,pH 值5.87,全氮9.93 g/kg,水解氮0.26 g/kg,有效磷0.64 g/kg,速效钾11.3 g/kg,有机质295 g/kg。
1.2 试验设计
2018 年5 月23 日进行扦插,将穗条基部裹上黄泥,扦插于轻型基质中。扦插一个月后(2018年6 月22 日)待苗木扦插成活,选取长势一致的幼苗作为试验材料进行指数施肥试验,使用氮磷钾复合肥(N∶P2O5∶K2O=16∶16∶16)对杉木幼苗进行施肥处理,采用霍格兰营养液对其进行中、微量元素补充,除氮、磷、钾外其他元素每个处理施入量相同,施肥量以施氮量来表示[11-12],以便直观呈现,施肥方案与进度见表1。

表1 杉木“洋061”无性系苗施氮进程表Table 1 Nitrogen application progress for the superior clone of C. lanceolate “Yang 061”
试验共设置3个处理,分别为总施氮量0 mg/株(不施肥处理为对照,CK)、总施氮量40 mg/株(常规集中施肥处理,CF)、总施氮量40 mg/株(指数施肥处理,EF);每个处理设置3 个重复,每个重复40 株苗。其中常规集中施氮总共施两次(第一周和第六周),每次施氮20 mg/株;氮素指数施肥每7 天施肥1 次,共施肥10 次,施氮量参照指数施肥模型[13]设置。杉木“洋061”无性系穗条初始氮含量为5.764 mg/株,试验期间进行正常的田间管理和苗木病虫害防治,施肥至2018年8 月24 日。
1.3 基质样品采集与处理
待施肥结束后半个月即2018 年9 月8 日,对苗木轻型基质进行取样,剪开基质袋放入塑料自封袋中,隔袋除去基质袋和扦插苗,混匀塑料袋中基质[12],取3g 装入无菌袋中并放入装有足量液氮的保温盒中。采用ITS rDNA 对其真菌群落多样性进行高通量测序。
1.4 基质养分测定
自2018 年6 月8 日—2019 年6 月8 日,每90 天测定1 次,每次选择3 株平均株。取基质混匀,放在干净的牛皮纸上自然阴干,研磨成粉状,过0.149 nm 细筛,采用H2SO4-HClO4法消煮基质,电感耦合离子发射光谱仪(PE optima 8000)测定基质养分。水解氮、有效磷、速效钾参照国标分别采用碱解扩散吸收法(LY/T 1229—1999)、氟化-盐酸浸提法(LY/T 1233—1999)、火焰光度计法(LY/T 1236—1999)。C、N 元素使用全自动碳氮分析仪(Vario Macro Cube)测定[14]。
1.5 基质真菌测定
1)DNA 提取
基质真菌总DNA 采用MOBIO 公司的PowerSoilKit 试剂盒提取;对提取后的DNA 产物经琼脂糖凝胶电泳进行检测,合格DNA 稀释至1 ng/μL 保存于-80 ℃用于PCR 扩增,采用带有Barcode 的特异引物扩增DNA 样本中特定区域,具体扩增信息见表2。扩增程序为:95 ℃预变性2 min,随后98 ℃变性10 s,62 ℃退火30 s,68 ℃延伸30 s,共27 个循环后68 ℃延伸10 min[15-21]。

表2 真菌多样性测序引物信息Table 2 Diversity sequencing primer information of fungal
2)PCR 产物的混样和纯化
PCR 产物使用浓度2%的琼脂糖凝胶进行电泳检测;根据PCR 产物浓度进行等量混样,充分混匀后使用1×TAE 浓度2%的琼脂糖胶电泳纯化PCR 产物,剪切回收目标条带。产物纯化试剂盒使用Thermo Scientific 公司GeneJET 胶回收试剂盒回收产物。
3)文库构建和测序
对扩增产物切胶回收,使用Quanti Fluor TM荧光计对回收产物进行定量。将纯化的扩增产物进行等量混合,连接测序接头,使用New England Biolabs 公司的NEB Next® Ultra ™ DNA Library Prep Kit for Illumina 建库试剂盒进行文库的构建,构建好的文库经过Qubit 定量和文库检测,合格后使用Hiseq 2500(Illumina, SanDiego)的PE250 模式上机测序。
1.6 数据处理
微生物群落多样性结果使用FLASH 软件对原始数据进行质量过滤和双端序列的拼接,得到有效数据。采用Uparse 软件,相似度≥97%的序列归为1 个操作分类单元即OTUs(operational taxonomic units)[22-24],利用Greengene 数据库进行物种注释;运用QIIME 软件对所得优质序列进物种分类分析,对OTUs 进行丰度、多样性指数等分析,将相对丰度大于0.1%的门与目定义为优势类群;使用Excel 2010 软件对数据进一步整理,SPSS 26.0 软件对数据进行单因素分析、Duncan 多重比较、person 相关性分析、主成分分析,采用Canoco 5.0 软件进行冗余分析(RDA),Orign 8.5软件进行图形绘制。
2 结果与分析
2.1 轻型基质理化性质对不同施肥方式的响应
不同施肥方式对基质养分含量产生了较大影响(表3)。与CK 相比,EF 处理后的基质AN、TP、TK、AP、AK、TN 含量增加幅度均大于CF处理,且CF 处理后的AN、AK 含量相较CK 有所降低;方差检验结果表明,EF、CF 处理较CK均显著提高了基质中TN、AP 含量,其中AP 含量变化最为显著,分别提高21.69%和12.44%;综上表明指数施肥较常规施肥更有利于基质养分积累。
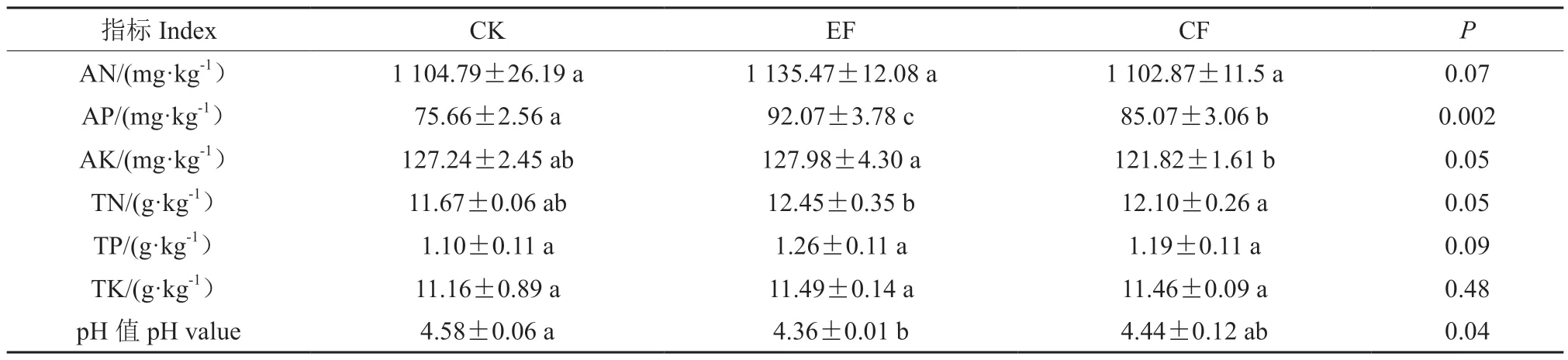
表3 指数施肥对基质理化性质的影响Table 3 Effect of exponential fertilization on physical and chemical properties of substrate
2.2 轻型基质真菌群落测序结果及OTU 统计分析
CK、EF、CF 处理获得的平均序列数分别为14 598、22 022、17 782,测序量均已达到饱和状态(表4)。通过韦恩图将各处理间真菌群落OTU 的重叠关系呈现出来[17](图1),EF、CF、CK 处理中的OTU 数分别为665、473、397,表现为EF >CF >CK;特有OTU 数分别为288、194、165,表现为EF >CF >CK,表明指数施肥对基质真菌群落OTU 有明显促进作用。
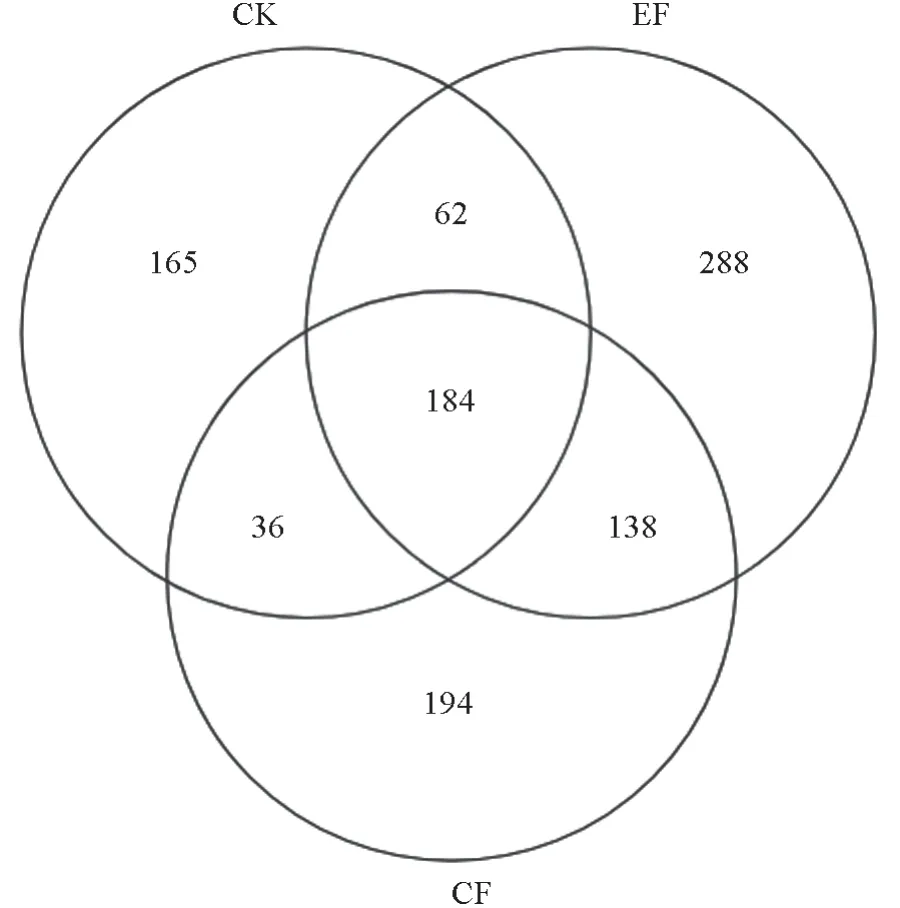
图1 不同施肥处理下真菌OTUs 韦恩图Fig. 1 Venn diagram of fungi OTUs under different fertilization treatments

表4 不同处理土壤真菌群落的α 多样性指数Table 4 α diversity indices of soil fungi community in different treatments
2.3 轻型基质真菌群落α 多样性与群落组成
基于整体OTUs 水平的基质真菌群落α多样性结果显示(表4),多样性指数(Shannon 指数和Simpson 指数)表现为EF >CK >CF,与CK处理相比,EF 处理下的Shannon 指数和Simpson指数分别增加15.21% 和7.41%,且Simpson 指数达到显著变化水平(P<0.05);CF 处理下的Shannon 指数和Simpson 指数则分别降低10.19%和3.85%,没有达到显著水平(P>0.05);EF、CF 处理下的丰富度指数(Chao1 和ACE 指数)较CK 均显著降低(P<0.05),表现为CK >CF >EF,EF、CF 处理下的Chao1 指数较CK分别减少47.21%、32.43%,ACE 指数分别减少43.16%、31.82%(P>0.05)。综上表明,对比CK,指数施肥会增加基质真菌多样性指数,降低丰富度指数。基于基质整体OTU 水平的相异系数bray-curtis 距离矩阵进行主坐标分析(图2),两轴共解释了群落结构差异的48.8%。Permanova 检验表明两种施肥下的真菌群落较CK 没有发生显著分离(P=0.37),综上表明短期两种施肥方式没有显著改变基质真菌群落组成。
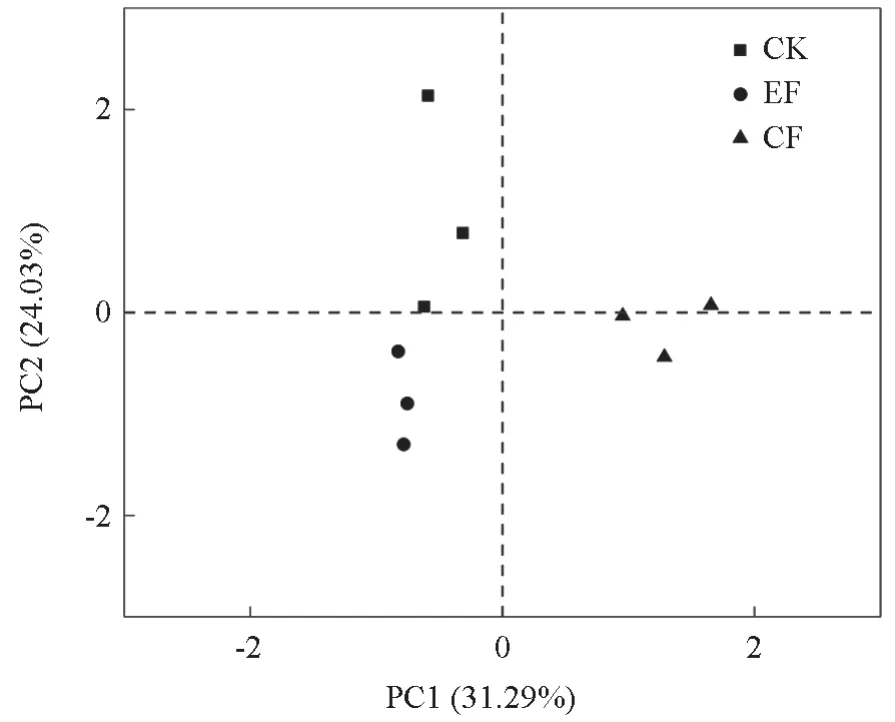
图2 不同施肥处理下的基质真菌群落NMDS 分布Fig. 2 NMDS distribution of matrix fungi community under different fertilization treatments
2.4 轻型基质真菌群落结构在门与属水平上的分布特征
高通量测序结果显示,3 个处理中共包含7门、23 纲、41 目、69 科、83 属。基于门水平,除去相对丰度小于0.1%和未能归类的其他类真菌,各个处理一共获得5 个类群(图3)分别为担子菌门Basidiomycota(40.61%~46.02%)、子囊菌门Ascomycota(25.97% ~35.94%)、被孢霉门Mortierellomycota(1.17% ~10.04%)、 壶菌门Chytridiomycota(0.43% ~4.01%)、 毛霉门Mucoromycota(0.32%~1.35%)。施肥后基质真菌门水平相对丰度发生明显变化,与CK 相比,EF 处理明显降低子囊菌门相对丰度约为25.72%(P>0.05),担子菌门呈相反趋势,增加了15%(P>0.05);被孢霉门相对丰度相较于CK 显著增加了2.5 倍(P<0.05),壶菌门减少了77%(P>0.05);CF 处理下的子囊菌门、担子菌门、被孢霉门相对丰度均呈下降趋势,壶菌门呈明显上升趋势,但均没有达到显著水平。为进一步探究指数施肥对基质真菌群落组成的影响,选择相对丰度大于0.1%的目水平(10 个)进行热图分析(图4),结果显示EF、CF 处理聚为一类,两种施肥后的真菌群落组成较CK 有明显分离,表明不同真菌目水平受施肥影响显著。与CK 处理相比,CF 处理下的粗糙孔目(Trechisporales)、粪壳菌目(Sordariales)相对丰度显著下降57.14%和48.13%, 而肉座菌目(Hypocreales) 相对丰度则显著增加3.6 倍;EF 处理下的粪壳菌目(Sordariales)相对丰度显著下降67.77%,而被孢霉目(Mortierellales)与肉座菌目(Hypocreales)的相对丰度较CK 处理显著增加2.5 和3.15 倍;EF 与CF 处理间仅粗糙孔目(Trechisporales)存在显著差异。
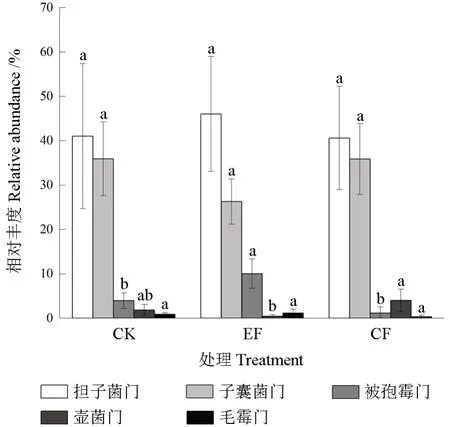
图3 基于门分类水平上的基质真菌群落结构组成Fig. 3 Community structure composition of matrix fungi based on phylum classification level
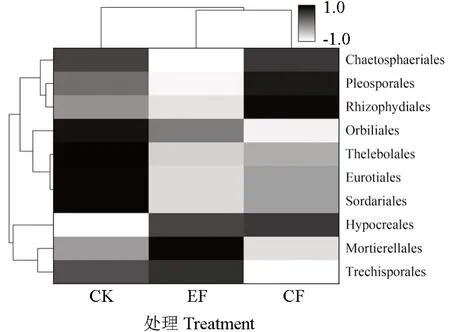
图4 各处理基质优势真菌在目水平的群落热图分析Fig. 4 Heat map analysis of the main fungi at the order level in each treatment substrate
2.5 轻型基质真菌多样性、群落组成与土壤理化性状的冗余分析
图5a 结果显示前两个轴特征值一共解释了基质真菌多样性变异程度的96.65%。其中,基质真菌多样性指数(Shannon 指数和Simpson 指数)与全氮、全钾、速效钾、pH 值呈正相关,与水解氮、全磷、速效磷呈负相关;丰富度指数(Chao1和ACE 指数)则与速效磷、全磷、水解氮、全钾呈正相关,与全氮、速效钾、pH 值呈负相关。Monte Carlo 检验结果表明,速效磷与全钾对基质真菌多样性贡献率最大,分别为39.2%、31.4%,且速效磷影响显著(P=0.02)。可见,本试验条件下,速效磷是驱动基质真菌多样性变化的主要环境因子。
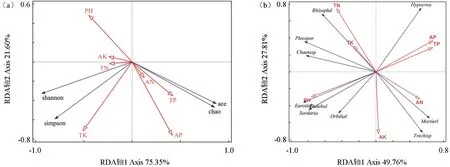
图5 基质真菌多样性指数、目水平类群分布与土壤理化性状间的多元分析Fig. 5 Multivariate analysis of diversity index, order level distribution of matrix fungi and soil physical and chemical properties
基于目分类水平(图5b),两个排序轴累计变量能在77.57%上解释基质真菌群落结构组成的差异性。壶菌目、格孢菌目、刺球壳目相对丰度均与全氮、全钾和pH 值呈正相关,与速效钾、水解氮、有效磷、全磷呈负相关,散囊菌目、圆盘菌目、Thelebolales、粪壳菌目相对丰度与pH 值和速效钾呈正相关,与全钾、全氮、水解氮、全磷和速效磷呈负相关,糙孢孔目、肉座菌目、被孢霉目相对丰度与速效磷、全磷、全氮呈正相关,与全钾、全氮、水解氮、pH值呈负相关。Monte Carlo检验结果表明,pH 值与全氮对基质真菌目水平群落分布的贡献率最大,分别为35.4%、25%,且二者影响均达到显著水平(P=0.014,P=0.008)。因此,pH 值与全氮是影响基质真菌群落分布的主要环境因子。
2.6 轻型基质真菌优势属与杉木苗生长特性的相关性分析
由表5 可知,通过EF 与CF 处理,杉木苗高、地径、总生物量相较CK 处理均有显著增加,均表现为EF >CF >CK。综上,在同等施肥量的情况下,指数施肥效果最佳,比传统常规施肥能更有效促进苗木生长。为进一步分析杉木生长特性与基质真菌群落的关系,将真菌群落差异物种(目水平)与杉木生长指标进行相关性分析发现(图6),苗高、地径、生物量粪壳菌相关性较小,与被孢霉目呈显著正相关(P<0.05),地径、生物量分别与糙孢孔目和肉座菌目呈显著正相关(P<0.05)。总体上说,基质真菌差异物种相对丰度与杉木幼苗生长有较为紧密的联系。其次,只有糙孢孔目与被孢霉目之间存在显著正相关,其余差异物种之间相关性相对较弱,存在较少的竞争关系。
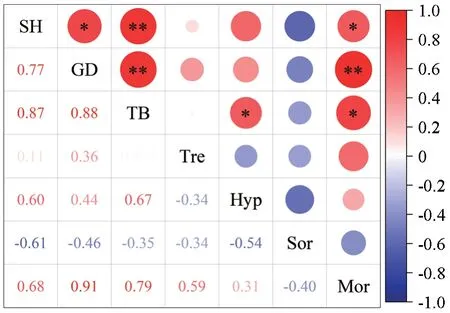
图6 基质真菌差异物种与杉木苗生长指标的相关关系Fig. 6 Correlation between different species of matrix fungi and growth index of C. lanceolata
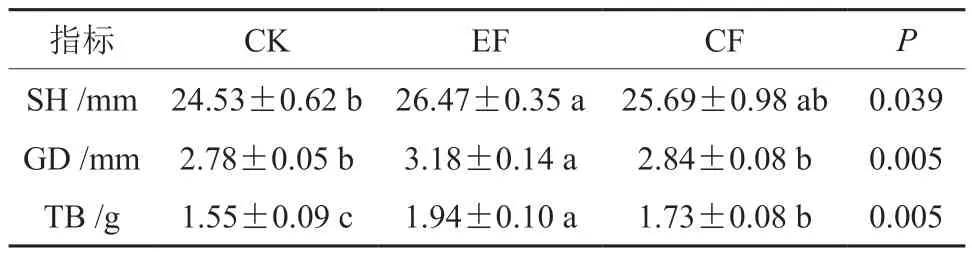
表5 不同施肥方式对杉木苗生长的影响Table 5 Effects of different fertilization methods on the growth of C. lanceolata seedlings
3 讨 论
3.1 指数施肥处理对杉木苗轻型基质真菌α 多样性的影响
目前有关施肥对真菌α多样性的影响结论并不一致,Yang 等[18]研究结果表明施肥会引起土壤微生物种间发生竞争,对营养元素敏感的微生物类群吸收营养元素以后迅速生长,进而会抑制其他对营养元素不敏感的微生物类群的生长与繁殖,最终导致微生物多样性降低[19];谢欢等[20]等通过选取杉木幼苗进行短期施肥发现,土壤真菌多样性指数明显下降,但没达到显著变化的水平;陈祖静[21]以桉树幼林为研究对象,进行短期不同氮水平添加后发现土壤真菌多样性指数与丰富度指数均有明显降低。在本研究中,两种施肥处理下的基质真菌丰富度指数较CK 呈显著下降趋势,分析认为,施肥改变了基质的微环境,促进了一些对环境变化敏感的微生物种群的生长,进而会抑制其他不敏感的微生物种群生长,导致整体丰富度下降;另外表示群落内优势种地位和作用的Simpson 指数在施肥后明显提高,在EF 处理下达到最大,这与陈祖静等[21]对桉树人工林土壤真菌多样性的研究结果大致相同。土壤理化性质对真菌多样性指数的影响已被证实,何敏红等[22]、王小玲等[3]、Zhou 等[23]发现,土壤pH 值、土壤有效养分含量、土壤有机碳含量是影响土壤真菌多样性变化的主要驱动因子。本研究中,速效磷是影响基质真菌丰富度与多样性指数的主要环境因子,这与张海芳等[19]研究结果一致,施肥能够有效改变土壤养分有效性进而直接影响土壤真菌多样性。此外,真菌周转时间较长,短期能在酸性环境中稳定生长,Sun 等[24]研究发现进行长期施肥较短期施肥能够显著改变土壤理化性质,影响基质真菌多样性。因此,在以后的实验中应考虑长周期施肥对真菌群落多样性的影响。
3.2 指数施肥处理对杉木苗基质真菌群落组成的影响
不同施肥处理下,土壤养分不同,微生物群落结构的整体组成也会存在差异,但优势种群基本一致。本研究发现各处理下的基质真菌优势菌门均为担子菌门、子囊菌门、被孢霉门,担子菌门与被孢霉门相对丰度在EF 处理下达到最大;子囊菌门相对丰度在施肥后均明显降低,这与Guo等[25]研究结果不一致,分析认为,施肥增加了水解氮含量,子囊菌门对N 有效性极为敏感,其相对丰度较不施肥前会有明显增加。这可能因为本研究的施肥水平较低,加之施肥时间较短,因此没有显著改变基质水解氮含量。另外,陈祖静等[21]对尾叶桉幼苗进行长达9 个月的施肥研究,发现短期施肥仅能改变对养分有效性较为敏感的真菌类群,Guo 等[25]通过对日本落叶松幼苗进行为期3 月的施肥实验,同样得到相似结果。在目水平上,粪壳菌目被李鑫等[26]报道不仅参与凋落植物营养器官的重要分解者,和肉座菌目一样在有机质分解过程中扮演重要作用,本研究发现两种施肥方式较CK 显著降低了粪壳菌目的相对丰度,RDA分析结果显示与水解氮呈负相关,表明粪壳菌目在短期两种施肥条件下,容易受N 有效性抑制。而肉座菌目相对与速效磷呈显著正相关,表明肉座菌目对P 有效性较为敏感。另外本研究还发现EF 较CK 处理显著增加了被孢霉目的相对丰度。Lee 等[27]证实被孢霉目拥有转化有机化合物的能力,当外界环境发生变化时,能通过利用植物分泌物来溶解土壤P。乔志伟等[28]也发现被孢霉目与速效磷呈显著正相关关系,这与本研究RDA 分析结果基本相同,说明被孢霉目在短期施肥后的土壤P 转化过程中有重要作用。其次,相关性分析结果显示被孢霉目与苗高、地径、生物量均有显著正相关关系,表明被孢霉目在杉木幼苗P 供应与基质P 转化过程中的扮演重要的微生物角色。谢欢等[20]研究结果同样证实,短期施肥能够通过调控与P 转化有关的真菌功能类群,分解土壤养分提高养分有效性,促进杉木幼苗生长。
因此推测,两种施肥方式可能在一定程度上促进了难分解有机质与P 转化相关的真菌类群生长,进而影响基质真菌群落结构,真菌类群加快了基质中速效养分的转化与积累,
为杉木幼苗生长提供了条件,其深层原因还需要结合其相应的真菌功能类群来揭示。有报道称杉木是丛枝菌根树种,菌根能通过增加宿主根表面积与地下菌根网络增加宿主对养分的利用率,因而后期实验还需要对植株根系上定殖的重要菌根类群进行鉴别。
4 结 论
两种施肥明显增加了基质中养分含量,与常规施肥相比,指数施肥显著增加了速效磷、速效钾、全氮的含量,提高了基质真菌Simpson多样性指数,降低了丰富度指数,促进了基质真菌优势种群增长,全氮、pH 值是影响基质真菌群落结构的主要环境因子。两种施肥方式能够通过调控杉木容器育苗基质的真菌群落组成来影响植物幼苗生长。指数施肥较常规施肥能显著促进磷转化和有机质分解真菌类群的增长,更有利于基质后期有效养分积累,为杉木幼苗生长提供良好条件。目前关于杉木容器育苗基质真菌群落结构对短期指数施肥方式的响应的研究相对较少,具体的影响机制还需要结合具体真菌功能类群与土壤环境因子之间的关系来进一步揭示。
