川西高原地区猕猴栖息地选择的季节差异
2023-12-22斯煌凯金祖祥张克处周飞姚永芳肖洪涛李八斤普错汪甲徐怀亮
斯煌凯 金祖祥 张克处 周飞 姚永芳 肖洪涛李八斤 普错汪甲 徐怀亮*
(1 四川农业大学生命科学学院,雅安 625014)(2 雅江县林业和草原局,雅江县自然保护区服务中心,雅江 627450)
栖息地是动物生存的地理环境,包括动物利用的一切生态环境因子的总和,为动物个体或种群的生存、繁衍等一系列生命活动提供保障 (颜忠诚和陈永林,1998;钟明等,2016)。不同动物生存繁衍的环境条件各有差异,因此动物对栖息地的选择具有偏好性。动物对栖息地选择呈动态变化,与其所处的时空尺度息息相关 (Groffet al., 2017; Allegueet al., 2022),同一物种不同种群,同一种群在不同季节,同一个体在不同生理时期,都会因生存需求的差异而调整选择策略 (张明海和李言阔,2005;武正军等,2012; Fuet al., 2022)。栖息地质量显著影响区域内物种的生存状态 (高梅香等,2011),从而对物种栖息地选择产生显著影响,目前栖息地破碎化、环境污染、人类活动等一系列问题使适宜物种生存繁衍的栖息地大量减少,这也成为世界范围内众多物种面临灭绝风险的重要原因 (Fahrig, 2003;Nickelet al., 2020)。因此,揭示野生动物栖息地选择规律对物种保护具有重要意义,也能促进对生态系统的认识与保护 (Johnson, 1980)。
猕猴 (Macaca mulatta) 是灵长目猴科猕猴属最具代表性的物种 (蒋学龙等,1991),其分布范围广泛,栖息地环境多样化程度高,在复杂多样的自然环境中都表现出极强的适应性 (路纪琪等,2018)。目前,有关猕猴地理分布、种群动态、行为生态等的研究已有较多报道 (Cuiet al., 2019;丁振芳等,2020;付文等,2021;张茹帆等,2021),但对于猕猴栖息地选择策略的研究仍较少。食物因子是影响猕猴栖息地选择的主要因素,四川泸山猕猴冬季为获取更多食物资源而靠近人类活动频繁的区域,依赖人工投喂解决冬季食物短缺的困境 (亓东明,2014),太行山猕猴缺乏食物的冬季会主动靠近人类 (谢东明等,2009);在云南白马雪山,猕猴冬季尽可能回避人类活动,选择在常绿阔叶林、针阔混交林觅食 (黎大勇等,2012);尼泊尔境内的猕猴多分布在海拔3 000 m 以下的热带、亚热带、温带森林中,这些区域食物资源更为丰富 (Wada, 2005);孟加拉国的猕猴多在森林密度较低或森林边缘活动,游客的投喂为其提供了大量食物 (Naheret al., 2016)。此外,人为干扰、水源等也是影响猕猴密度和分布的重要因素 (张忠员等,2010; Kumar, 2017)。然而大多现有研究聚焦于猕猴单季内短期的栖息地选择策略,忽略了猕猴面对环境变化在不同季节对栖息地选择做出的适应性调整。长期的栖息地选择研究能更客观反映动物选择的事实 (Yamagiwaet al., 1998),但相关研究往往着眼于栖息在中低海拔地区的物种(余鑫等,2009),对高原地区的物种缺乏较多的关注,仅对雪豹 (Panthera uncia)、藏羚羊 (Pantholops hodgsonii) 等物种有较多报道 (Qiet al., 2015;Baiet al., 2018; Honget al., 2021)。
高原地区的猕猴种群相比其他区域的猕猴具有独特的生物学特征,在形态、种群遗传等方面都与低海拔种群有明显差异 (蒋学龙等,1991;苏倩,2019),这与猕猴适应高寒、低氧、强紫外线等恶劣高原自然条件相关。本研究以川西高原地区一猕猴群体为研究对象,通过一个年周期的跟踪观察,分析其在不同季节对栖息地的选择与利用,以探究高原地区猕猴栖息地选择随自然环境动态变化的规律。我们推测:不同季节猕猴的栖息地选择存在较大差异;食物资源变化是影响猕猴不同季节栖息地选择的主要因素;高原人类活动少,猕猴对人为干扰较为敏感,避免与人类接触;夏季猕猴对水源依赖降低,而在冬季,水源是栖息地选择的重要影响因素。
1 研究方法
1.1 研究地点
研究区域属于四川省甘孜藏族自治州雅江县,地处雅砻江中游、青藏高原东南缘横断山区,受青藏高原复杂地形影响,为典型大陆性季风高原气候,日照强、紫外线辐射强、昼夜温差大。雅江县境内有格西沟国家级自然保护区及神仙山、庆达沟、亿比措、那溪措等省级自然保护区。
研究地点位于雅江县西俄洛乡、红龙乡及柯拉乡三乡交界的嗄波沟,地理坐标北纬29°03′ ~30°30′,东经100°19′ ~ 101°26′,处于四川格西沟国家级自然保护区与四川神仙山省级自然保护区之间。研究区域海拔3 500 ~ 4 300 m,植被类型随海拔变化呈现明显垂直分布带,3 500 ~ 3 700 m 主要为针阔混交林及灌丛,3 700 ~ 4 100 m 为针叶林与高山栎林交错分布,4 100 ~ 4 300 m 主要为草地,4 300 m 以上多为流石滩。一条主干道路穿过整个研究区域,区域内人为干扰强度较大,主要为附近村庄居民放牧、砍伐、林下采集等。
1.2 研究对象
2019年7月至2021年3月,选择长期生活于研究区域内的一个猕猴群体进行跟踪观察。依据实际跟踪观察,从猴群清晨离开夜宿地开始取样,跟踪猴群至天黑回到夜宿地为完整取样日;从发现猴群的任意时刻起进行观察取样,到天黑猴群回到夜宿地或从观察者视线消失超过30 min为非完整取样日 (黄中豪等,2010)。累积跟踪观察猕猴178 日,其中完整取样123 日,非完整取样55日。研究期间,猴群内有幼猴出生,亦有亚成体离群,个体数保持在40 只左右。猕猴是典型的杂食性动物,食物多样化程度高,以植物性食物为主,喜食植物的根、茎、芽、嫩叶、花、果实等,食物缺乏期也会啃食树皮 (吕九全等,2002)。区域内猕猴的主要天敌为普通鵟 (Buteo japonicus)、草原雕 (Aquila nipalensis) 等猛禽以及豹 (Panthera pardus)、猞猁 (Lynx lynx) 等大中型食肉目动物。
1.3 季节划分
根据研究期间记录的气温数据 (图1),并结合观察当地的物候情况,按照“高原物候季节划分法”(范思睿等,2011),对研究地区季节进行了划分:春季为4—6 月,平均气温 (10.37 ± 2.41) ℃,初春时期植物嫩芽膨大生长,逐渐开花,晚春大多植物绿叶封冠,开花植物较少;夏季为7—9月,平均气温 (12.33 ± 1.12) ℃,初夏为二次开花盛期,到了仲夏、晚夏开花的植物逐渐减少,此时叶片已达成叶面积,且夏季为全年光照最为充足时期,植物进入光合作用最旺盛阶段,植被茂密;秋季为10—11 月,平均气温 (7.33 ± 2.06) ℃,入秋后植物叶片迅速枯黄,果实和种子成熟至凋落,11 月下旬气温快速下降,因此秋季时间短,仅有2个月;冬季为12月至次年3月,平均气温 (0.68 ±2.26) ℃,天气寒冷干燥,大部分植物枯萎。相比平原地区,高原地区入冬早、入春晚,积雪难以融化,因此冬季更为漫长。
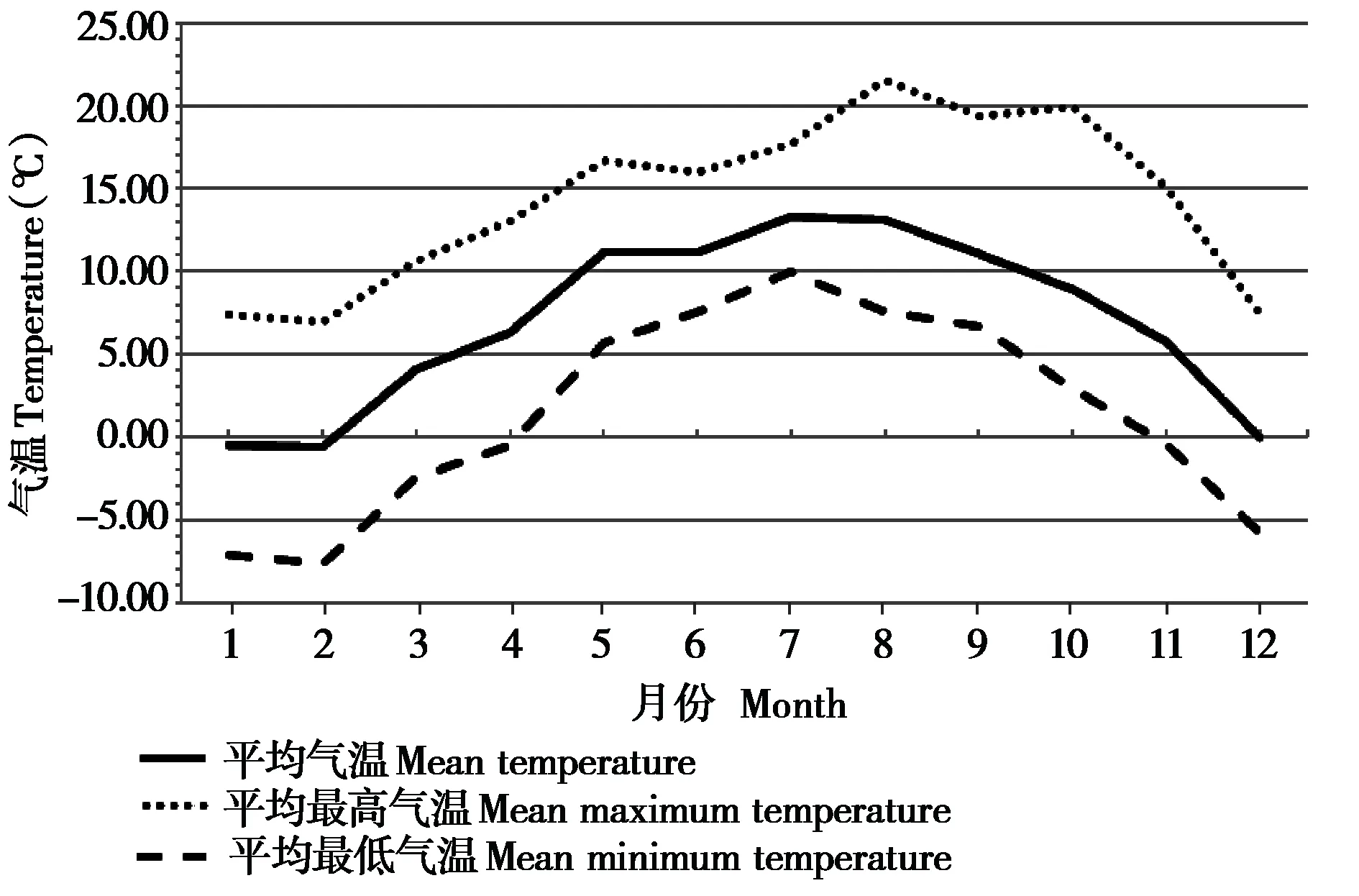
图1 研究区域的月平均最低气温、平均气温、平均最高气温Fig. 1 Monthly mean minimum temperature, mean temperature,mean maximum temperature in the study area
1.4 样方设置
通过长期跟踪观察,确定猴群的主要活动区域。在实际调查中,若直接观察到猕猴活动 (觅食、休憩、玩耍等) 或有猕猴新鲜湿软粪便样本(调查区域内无其他灵长类动物,且猕猴粪便与当地其他野生动物粪便易于区分),则利用GPS记录猕猴活动位置的中心位点,调查期间共记录到135个位点,并在每个位点设置20 m × 20 m的利用样方,同时在区域内无猕猴活动及痕迹的区域设置20 m × 20 m 的对照样方共60 个,对照样方距离最近的利用样方至少大于1 km,并使单季内的利用样方和对照样方覆盖研究区域内所有栖息地类型(刘振生等,2009)。共计设置样方195个 (表1)。
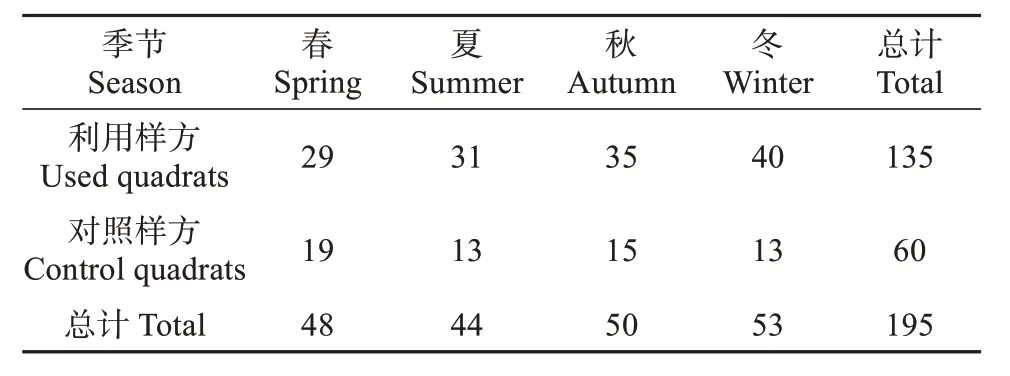
表1 研究区域不同季节样方设置Table 1 Setting of quadrats in different seasons in study area
1.5 生态因子指标
根据研究区域内的相关环境特征,选取了18 个相关的生态因子,这些因子对满足猕猴生存繁衍需求高度相关 (黎大勇等,2012; Xieet al.,2012;赵海涛等,2015)。调查并记录每个样方内18 个生态因子的具体指标,不同变量采取不同的衡量手段 (表2)。

表2 猕猴栖息地选择生态因子划分标准Table2 Details of ecological factor for habitat selection about Macaca mulatta
1.6 数据分析
使用Vanderloeg 选择系数Wi(Lechowicz, 1982)和Scavia 选择指数Ei(Chesson, 1978),评价猕猴对栖息地的偏好程度及其季节差异,公式如下:
在公式 (1) (2) 中,i为特征值,n为特征值总数。pi表示调查的所有样方中具有i特征值的样方总数,ri表示猕猴选择的具有i特征值的样方数。Ei值的范围为 [-1、1]。当Ei= -1 时表示回避 (None, N),当-1 ≤Ei≤ -0.1 时表示避免选择 (No preference,NP),当-0.1 <Ei< 0.1 时表示随机选择 (Almost random, AR),当0.1 ≤Ei≤ 1时表示偏好选择 (Preference, P),当Ei= 1时表示积极选择 (Strong preference, SP)。
使用SPSS 22.0,对影响猕猴栖息地选择的18 个生态因子数字化变量进行主成分分析 (Principal component analysis, PCA),得到相关的特征根、特征向量、主成分和贡献率。提取特征值大于1 的主成分进行分析,以确定不同季节影响猕猴栖息地选择的主要生态因子 (黎大勇等,2012;王伟等,2013;朱洪强等,2019)。
2 结果
2.1 猕猴栖息地选择的季节变化
研究获得猕猴4 个季节对18 个生态因子的选择指数,并确定了猕猴选择偏好 (附表A,表3)。春季猕猴偏好在海拔3 900 ~ 4 100 m活动,夏、秋季偏好海拔3 900 ~ 4 300 m的区域,而冬季则更偏好栖息在海拔3 500 ~ 3 900 m的区域。冬、春季猕猴仅选择山脚与溪谷活动,对其他地形均回避;夏季偏好山谷与山鞍部,秋季偏好山脊与山鞍部。猕猴在春、秋季偏好选择阳坡活动,夏季多在阳坡与半阴半阳坡活动,冬季对坡向表现为随机选择。猕猴夏、秋季仅在坡度小于40°的平缓坡活动,冬、春季在坡度小于70°的区域均能够见其踪迹。猕猴在夏、秋季偏好选择中坡位,在冬、春季偏好下坡位。
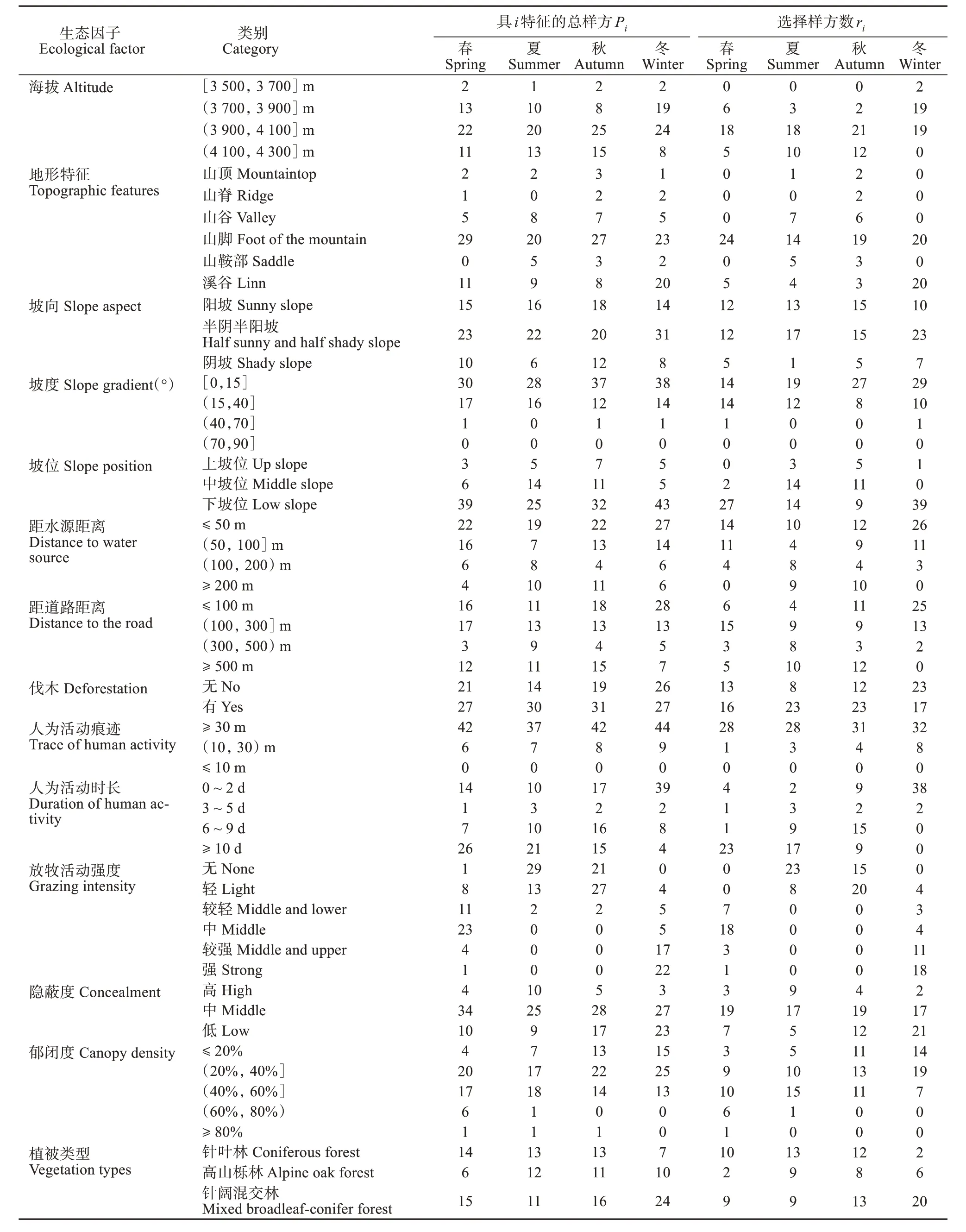
附表A 川西高原猕猴四季栖息地选择的生态因子特征样方Appendix A Ecological factor characteristic plots for habitat selection of macaques in the Western Sichuan Plateau during the four seasons

表3 川西高原猕猴四季栖息地选择偏好Table 3 Preference for habitat selection of macaques in the Western Sichuan Plateau during the four seasons
猕猴对高山流石滩与高山草地 (山顶) 表现出明显回避,对针叶林、高山栎林、针阔混交林、落叶阔叶灌丛、草地 (河谷) 的选择因季节而不同。夏季回避灌丛与低地草地 (河谷附近),秋季回避灌丛。夏季猕猴偏好选择乔木胸径大于20 cm 的栖息地,秋季则偏好乔木胸径大于40 cm 的环境,而冬季偏好在乔木胸径10 ~ 30 cm 的区域活动,春季对乔木胸径的选择无明显偏好。春、夏季猕猴偏好选择的栖息地乔木均高为10 ~ 20 m,秋季更偏好乔木均高大于10 m 的乔木,而冬季更多活动于乔木均高小于10 m 的区域。猕猴在夏、秋季偏好灌木均高小于2 m的栖息地,回避灌木均高大于3 m的栖息地,但在冬、春季偏好灌木均高大于3 m 的栖息地。对草本均高,猕猴偏好草本植物长势越好 (均高越高) 的栖息地。春季猕猴偏好选择在郁闭度大于60%的区域活动,夏季偏好郁闭度40% ~80%的栖息地,而秋、冬季多在郁闭度小于60%的区域活动。夏季猕猴偏好选择高隐蔽度栖息地,冬季则偏好低隐蔽度栖息地,而春、秋季对隐蔽度为随机选择。冬、春季猕猴多选择距水源小于100 m 的区域活动,而夏、秋季偏好选择距水源100 ~ 200 m的栖息地。
人为干扰因子中,夏季猕猴偏好选择在距道路300 m 以上的区域活动,冬季偏好选择距道路小于300 m 的区域,春季偏好在距道路100 ~ 500 m处活动,而秋季表现为随机选择。猕猴对已发生伐木活动的区域四季都表现为随机选择。猕猴偏好距人为活动痕迹大于30 m 的区域,但在冬季偏好选择距人为活动痕迹10 ~ 30 m 的栖息地,除冬季外不选择距人类活动时长小于2 d 的栖息地。夏、秋两季猕猴偏好无放牧活动或轻放牧的栖息地,而冬、春季选择放牧活动频繁的区域。
2.2 猕猴栖息地选择的主要影响因素
为确定猕猴不同季节栖息地选择的主要影响因素,分别对18 个生态因子进行主成分分析。春季栖息地选择的前八个主成分特征值大于1,累积贡献率高达84.095%,夏季、秋季和冬季则均为前六个主成分特征值大于1,累积贡献率分别为72.724%、77.096%、76.139%,表明这些主成分已包含了海拔、坡度、坡向、放牧活动强度、郁闭度、草本均高等18 个参数的所有信息,可代表猕猴不同季节栖息地选择特征 (附表B)。
对每个季节特征值大于1的主成分进一步计算其特征向量的得分矩阵。春季前八个主成分中,主成分1 的乔木均高 (0.814)、主成分2 的海拔(0.675)、主成分3 的坡向 (0.734)、主成分4 的植被类型 (-0.582)、主成分5 的地形特征 (0.725)、主成分6的放牧活动强度 (0.579)、主成分7的草本均高 (0.740) 以及主成分8 的放牧活动强度(0.528),是各个主成分中相关系数绝对值最大的变量;夏季前六个主成分中距道路距离 (-0.820)、乔木均高 (0.806)、 隐蔽度 ( - 0.742)、 坡度(0.615)、放牧活动强度 (0.649)、坡位 (-0.396) 为各个主成分中相关系数绝对值最大的变量;秋季前六个主成分中距道路距离 (0.902)、乔木均高(0.800)、放牧活动强度 (0.674)、坡度 (0.513)、人为活动痕迹 (0.609)、坡度 (0.664),为各个主成分中相关系数绝对值最大的变量;冬季前六个主成分中地形特征 (0.868)、坡度 (-0.798)、海拔(0.824)、 坡位 (0.743)、 坡位 (0.550)、 伐木(0.523) 是各个主成分中相关系数绝对值最大的变量。
根据表2中各个生态因子的分类及上述主成分相关系数,表明影响猕猴春、秋季栖息地选择的主要因子为地理特征、人为干扰和植被;影响猕猴夏季栖息地选择的主要因子为地理特征、人为干扰、隐蔽条件和植被;影响猕猴冬季栖息地选择的主要因子为地理特征和人为干扰。
3 讨论
猕猴对环境的适应能力强,自然分布广泛、生境多样,栖息环境可从近海平面至海拔4 000 m以上高原地区 (Fooden, 2000;路纪琪等,2018)。生存于高原环境下的猕猴相比低海拔猕猴面临更恶劣的自然条件,如食物匮乏、强紫外线辐射、寒冷干燥等,在长期的进化过程中,形成了相对特殊的栖息地选择策略。动物栖息地选择不仅取决于动物本身,还与栖息地的食物丰度、隐蔽度、种间或种内竞争等因素密切相关,是遗传与适应共同作用下形成的 (杨萃等,2011;张阳等,2022)。一般认为,食物、水源与隐蔽条件是动物栖息地选择的三大要素 (马建章等,2004)。一年中季节性的气候变化是引起这些生态因子变化的直接原因,动物的栖息地选择也会随季节而改变。
本研究发现,在川西高原地区,尽管猕猴的栖息地总体上已属于高海拔地区 (3 500 m 以上),其活动范围在海拔上仍表现出明显的季节性分化。在春、夏、秋三季,川西高原地区的猕猴主要活动于3 900 ~ 4 300 m的较高海拔地带,而冬季主要活动于3 500 ~ 3 900 m的相对较低海拔地带。究其原因,可能是因为春、夏、秋三季较高海拔地带的气温较高,食物资源丰富,且人为干扰少,而较低海拔地带靠近居民点、农田、公路等人类活动区,人为干扰较大,因而猕猴在春、夏、秋季尽可能回避人类活动的干扰,多活动于较高海拔地区;冬季整个高原地区气候寒冷、食物贫乏,较高海拔地带更为显著,因而猕猴在冬季不强烈回避人类活动的干扰,更多地选择较低海拔地带活动,这与泸山猕猴冬季常靠近寺庙等人类活动区域取食的行为类似 (亓东明,2014)。对于地理特征因子的选择,猕猴倾向选择地形平缓、地势起伏小的栖息地,这有助于减少活动过程中的能量损耗;冬、春两季猕猴对山脚、溪谷等区域的偏好以及对山脊的回避也是一种对高原极端环境适应的觅食和能量分配策略,因为高原地区山脊、陡坡处的植被生长条件差、避风性弱,而山脚、溪谷区域地势平缓、水热环境适宜植物生长,食物资源丰富。但对较低海拔的太行山及王屋山的猕猴行为研究发现,这些地区的猕猴在冬季也常出没于山脊地形 (谢东明等,2009; Xieet al.,2012)。这些结果表明,在气候恶劣、自然生产力低下的高原环境中,猕猴的觅食行为策略更为保守,这与众多灵长类动物在应对食物短缺时采取“低成本-低收益”的能量平衡策略一致 (Agetsuma, 1995;赵海涛等,2015; Liet al., 2019)。不同季节猕猴对坡向选择存在差异,春、秋季猕猴倾向在植被繁茂的阳坡活动,而夏季对坡向的喜好扩散至半阴半阳坡,冬季对坡向则表现出随机选择,该结果与低海拔地区猕猴大多偏向选择阳坡活动不同,这可能与高原地区冬季食物更为匮乏,猕猴需进一步扩大活动范围寻找食物有关。总之,地理特征因子的选择偏好与食物资源、能量消耗高度相关,相比低海拔地区,高原地区的猕猴在食物缺乏季节更注重尽可能减少能量损耗而获取食物资源。
植被因子往往与食物因素直接相关,同时也关系到隐蔽条件的优劣。夏季猕猴多在食物资源丰富的林区活动,几乎不到低地草地活动,其余季节林区和低地草地均有活动,可能是夏季林区不仅具有丰富的食物资源,林荫还能够减少夏季强光照对动物体的伤害。另外,已有一些研究表明,灵长类喜欢在有高大树木的环境中活动 (谢东明等,2009;韩宗先等,2011;黎大勇,2015;余吉等,2017),高大树木树枝粗壮、林冠茂密,方便猕猴在林上层移动从而减少能量消耗,若遭遇捕食风险,也能为猕猴提供良好的隐蔽条件,降低捕食风险。春、夏季是猕猴的繁殖季,为避免猛禽捕食幼猴,多在郁闭度大于60%的栖息地中活动,但秋、冬季更多在郁闭度小于60%的栖息地活动,可能是由于低郁闭度环境太阳辐射相对较强,有助于猕猴在寒冷季节抵御低温。对于水源的需求,冬、春季气候干燥,食物中水分少,猕猴偏好在靠近水源的区域活动;而夏、秋季,猕猴可从食物中摄取较多水分,且该区域夏季阵雨频繁,在低洼处易形成临时性小水塘,猕猴对河流水源的需求降低,这与神农架川金丝猴 (Rhinopithecus roxellana) 在不同季节对水源的选择类似 (铁军,2009)。
本研究发现,相比低海拔地区的猕猴,高海拔环境下的猕猴对人为干扰有更为灵活的选择偏好,可能与高海拔环境下食物资源缺乏相关。夏、秋季猕猴对人类活动有明显回避,偏好在高隐蔽度的区域活动,且对放牧活动高度戒备,因为夏、秋季自然环境中的食物资源能够满足其生存需要,无需增加风险靠近人类。但冬、春季不一样,尤其在冬季,猕猴活动区域与人类活动区域高度重叠,调查过程中发现冬季猕猴常与牦牛在同一草地觅食。冬季食物资源缺乏使猕猴对草本植物的依赖增加,为提高生存机会,其不得不权衡人类活动带来的风险。而当地牧民在遭遇猕猴时也存在投喂现象,使猕猴在冬、春季与人类接触显著增加。在调查分析前,我们认为猕猴会回避伐木区,因为伐木区缺少高大乔木、郁闭度差、人为干扰严重,但结果显示猕猴在伐木区仍有活动。伐木区有大量的倒木,腐烂后是昆虫繁衍栖息的优良场所,鞘翅目昆虫的幼虫、白蚁等常在其中筑巢。在调查中发现,猕猴会剥开这些腐木的表皮取食藏匿于其中的昆虫。昆虫是高质量的食物,能够为猕猴提供优质蛋白质,相关食性研究也证明猕猴存在食虫性 (Luet al., 2007; Tomar and Sikarwar, 2014),因此高质量的食物可能是吸引猕猴在伐木区活动的主要原因。
综上所述,川西高原的猕猴根据不同季节的自然资源差异以及人为干扰而产生不同的栖息地选择偏好,地理特征因子、人为干扰因子、植被因子与隐蔽条件因子发挥着重要的作用。与其他低海拔地区的猕猴相比,在四季中随自然环境的变化而最大限度获取高质量食物资源及减少能量消耗,并灵活权衡食物资源与人类干扰的利弊,是该地区猕猴在自然生产力低下的高原环境中生存和繁衍的关键。保护好区域内的自然环境,合理安排放牧、采集、耕种等人类干扰活动,能够减少人兽冲突,从而有效促进该区域人类与猕猴的和谐相处。
致谢:本次调查得到四川省林业和草原局、雅江县西俄洛镇人民政府的大力支持及汪堆村村民提供的帮助,在此一并致谢!
