雄激素代谢相关基因AKR1C3、SHBG、SRD5A2 单核苷酸多态性与指长比的相关性
2023-12-14马成凤马秀慧杨梦怡马占兵霍正浩
马成凤, 李 帆, 吴 曦, 马秀慧, 张 静, 杨梦怡, 马占兵,陆 宏, 霍正浩, 党 洁
(1.宁夏医科大学基础医学院,银川 750004; 2.宁夏医科大学生育力保持教育部重点实验室,银川 750004)
指长比是指各指长度的比值,本研究指人类同一只手食指指长(2D)和无名指指长(4D)的比值,女性大于男性,是个体出生前宫内性激素暴露水平的潜在生物学标记[1-2]。2D∶4D 与孕期母体宫内雄激素和雌激素的比例密切相关,雄激素水平与指长比呈负相关[3]。研究[4-6]显示,性激素代谢途径相关基因多态性,如雄激素受体(androgen receptor,AR)基因、LIN28B 基因、CYP19A1 基因等与指长比存在关联性。2018 年,全基因组关联研究(genome-wide association study,GWAS)发现11 个单核苷酸多态性(single nucleotide polymorphism,SNP)位点与指长比强相关,能够解释平均3.8%的指长比差异[7]。由于性激素代谢途径复杂,作用区域广泛,不仅可影响机体的许多表型,同时,也可能影响指长比的形成。因此,指长比成为个体发育和生理功能外在宏观生物学的标记,受到生物人类学和临床医学的广泛关注[8]。
2 型3α-羟基类固醇脱氢酶醛酮还原酶(AKR1C3)、性激素结合球蛋白(SHBG)和睾酮5-α 还原酶Ⅱ(SRD5A2)基因在雄激素合成代谢过程中发挥重要作用。研究[9]证实,AKR1C3 可以促进雄激素、雌激素和孕激素与它们的同源无活性代谢物相互转化,优先将雄烯二酮转化为睾丸激素,是细胞合成睾酮和二氢睾酮(dihydrotestosterone,DHT)的关键酶;SHBG 既可以调节体内雄激素水平,也可以介导性激素的信号传导并调节其生物利用度[10];SRD5A2 是雄激素合成过程中将睾酮转化为DHT 的关键酶,能够增强DHT 的生物活性以及与AR 的亲和力,在性别分化和雄激素代谢中起到核心作用[11]。目前,有关雄激素代谢相关基因参与指长比形成的研究鲜有报道。本研究旨在分析健康人群AKR1C3、SHBG、SRD5A2基因6 个SNP 位点与指长比的相关性,进一步探讨雄激素对指长比形成的调控机制,为人类指长比发育的生物学基础提供理论依据。
1 资料与方法
1.1 研究对象
选取宁夏医科大学2019 级在校大学生799名为研究对象,男性396 名,女性403 名,平均年龄(19.18±0.76)岁。所有参与者均为宁夏籍大学生,三代祖居宁夏,彼此无血缘关系,无既往慢性疾病史及手术外伤史、无手部残疾或手部精细结构缺失。本研究已通过宁夏医科大学伦理审查委员会批准(宁医大伦理第2022-G185 号),所有参与者均知情同意。
1.2 研究方法
1.2.1 指长比的测量及分析 通过数码相机(Canon EOS M3,日本)提取双手掌面照片。照片采集时,被测者双手手掌向上,手指并拢,用力伸直放于桌面,由专门培训的研究人员依据统一标准和要求[右侧放置标准直尺(30 cm)作为标准,保持拍摄距离为垂直于手部正上方50 cm 处]进行拍摄,将照片(剔除手指有创伤的样本)导入图像分析软件Image-Pro Plus(Media Cybernetics,Inc.),参考软件操作手册,对各研究对象左手和右手各指(拇指除外)长度,即从近体侧手指折痕中点到指尖中点的直线距离进行测量。所有测量均经过2 人分别重复3 次后完成,最终取测量均值完成双手指长比的计算。
1.2.2 SNP 筛选 采用基因组SNP 在线数据库SNPinfo Web Server(http://snpinfo.niehs.nih.gov/)进行SNP 位点筛选。要求所选位点需满足:1)涵盖基因上下游1 000 kb 范围,包括外显子、内含子、5’-UTR、3’-UTR 等区域;2)最小等位基因频率(minor allele frequency,MAF)≥0.05;3)有过文献报道[12-15]的位点。最终选取rs12529、rs1937845、rs523349、rs727428、rs3760213 和rs6259共6 个SNP 位点进行研究,所有SNP 位点信息见表1。

表1 AKR1C3、SHBG、SRD5A2 基因6 个SNP 位点信息
1.2.3 DNA 提取及基因分型 使用一次性真空采血管(EDTAK2 抗凝管,5 mL)采集所有研究对象静脉血5 mL,低温(0~4 ℃)转运,随后快速置于-80 ℃冰箱冻存。所有DNA 样本采用血液基因组DNA 提取试剂盒(TIANGEN)[天根生化科技(北京)有限公司]提取全血细胞DNA,采用多重聚合酶链反应技术(北京诺禾致源科技股份有限公司)进行基因分型,各位点引物信息见表1。PCR 反应分两轮进行,第一轮:95°C 预变性15 min;94 °C 30 s,60 °C 10 min,72 °C 30 s,共4 个循环;94°C 30 s,60°C 1 min,72°C 30 s,共24 个循环。第二轮:95°C 预变性15 min;94°C 30 s,60°C 4 min,72°C 30 s,共5 个循环;94°C 30 s,65°C 1 min,72°C 30 s,共10 个循环。随后,通过CA2 吸附柱纯化PCR 产物后于Illumina X-10 测序平台上机测序,并采用Illumina RTA 软件进行质量控制和原始数据分析。
1.3 统计学方法
利用Haploview 4.2 软件完成Hardy-Weinberg 遗传平衡(HWE)检验,并进行单倍型的构建及连锁不平衡分析。利用SPSS 23.0 软件对数据进行统计学分析。不同性别2D∶4D 比较采用t 检验。基因型及等位基因频率采用Pearson 卡方检验。不同性别采用单因素方差分析方法比较目标基因不同基因型与人群间指长比分布的差异。P≤0.05 为差异有统计学意义。
2 结果
2.1 不同性别2D∶4D 分布比较
女性左手2D∶4D(t=-2.903,P=0.004)及右手2D∶4D(t=-2.339,P=0.02)均高于男性,见表2。

表2 不同性别2D∶4D 分布比较
2.2 AKR1C3、SHBG、SRD5A2 基因6 个SNP 基因型及等位基因频率分布
全部6 个SNP 位点基因型的分布符合HWE定律(P>0.05),提示样本具有较好的代表性。
AKR1C3 基因的rs12529 位点基因型频率分布在性别间存在差异(χ2=7.806,P=0.02),但其等位基因频率在性别间差异无统计学意义(P>0.05);SHBG 基因rs3760213(χ2=6.642,P=0.036;χ2=5.873,P=0.015)和rs6259(χ2=7.468,P=0.024;χ2=4.481,P=0.034)位点的基因型和等位基因型频率在性别间差异有统计学意义,见表3。

表3 AKR1C3、SHBG、SRD5A2 基因6 个SNP 位点基因型和等位基因频率[例(%)]
单倍型分析结果显示,AKR1C3 基因的rs12529-rs1937845 位点与 SHBG 基因的 rs727428 -rs3760213-rs6259 位点存在强连锁(图1),然而,只有后者构建得到的3 种单倍型在男女间差异有统计学意义(χ2=6.354,P=0.042),见表4。

图1 AKR1C3、SHBG 和SRD5A2 基因6 个SNP 位点连锁不平衡分析

表4 AKR1C3 和SHBG 基因SNP 单倍型性别间差异分析[例(%)]
2.3 AKR1C3、SHBG、SRD5A2 基因6 个SNP 基因型频率与不同2D∶4D 的关系
AKR1C3、SHBG、SRD5A2 基因的6 个SNP基因型频率均与不同性别2D∶4D 无关(P 均>0.05),见表5。
表5 AKR1C3、SHBG、SRD5A2 基因6 个SNP 位点基因型与不同性别2D∶4D 的关系(±s)
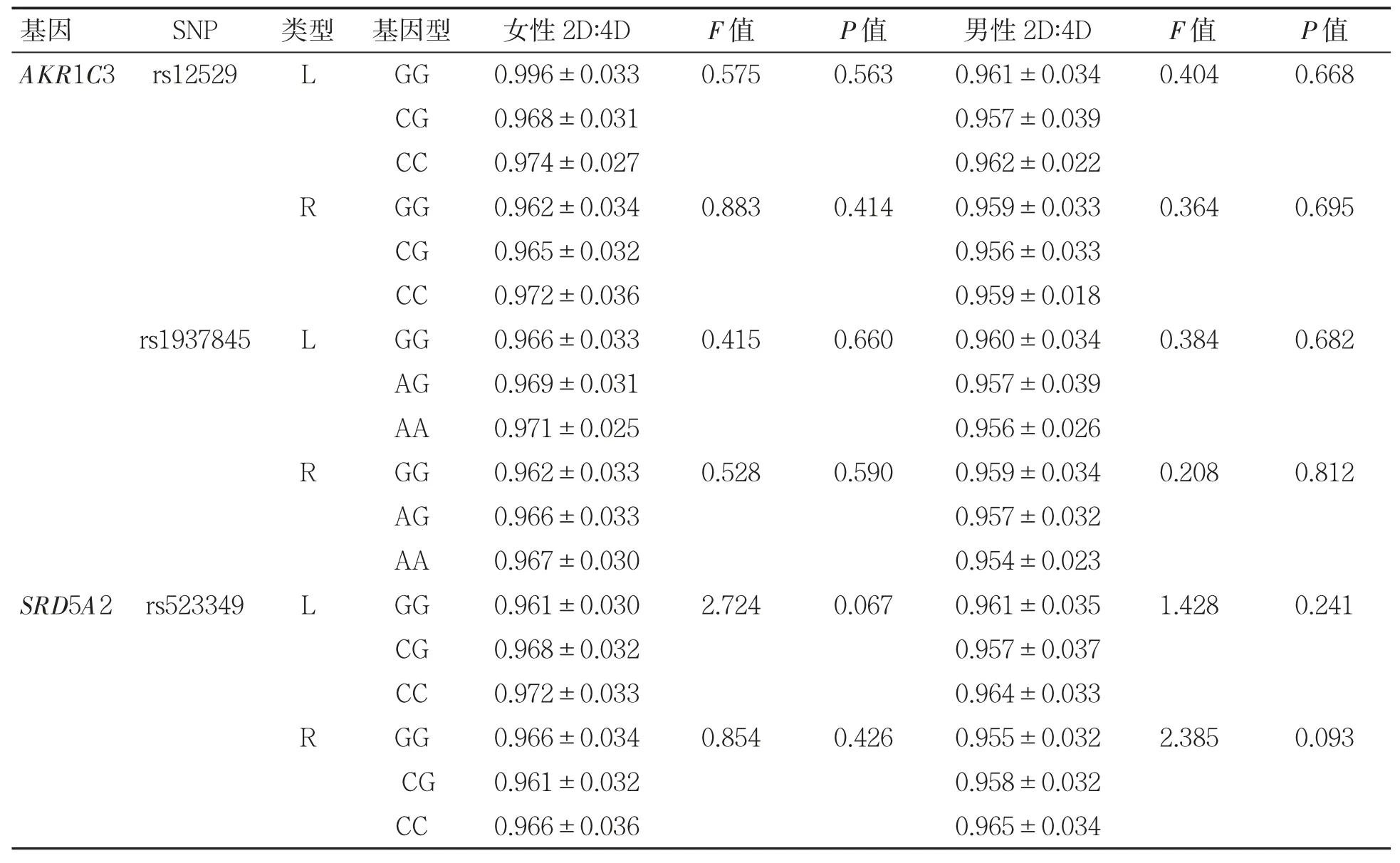
表5 AKR1C3、SHBG、SRD5A2 基因6 个SNP 位点基因型与不同性别2D∶4D 的关系(±s)
女性2D∶4DF 值P 值男性2D∶4DF 值P 值AKR1C3 rs12529LGG0.996±0.0330.5750.5630.961±0.0340.4040.668 CG0.968±0.0310.957±0.039 CC0.974±0.0270.962±0.022 RGG0.962±0.0340.8830.4140.959±0.0330.3640.695 CG0.965±0.0320.956±0.033 CC0.972±0.0360.959±0.018 rs1937845LGG0.966±0.0330.4150.6600.960±0.0340.3840.682 AG0.969±0.0310.957±0.039 AA0.971±0.0250.956±0.026 RGG0.962±0.0330.5280.5900.959±0.0340.2080.812 AG0.966±0.0330.957±0.032 AA0.967±0.0300.954±0.023 SRD5A2 rs523349LGG0.961±0.0302.7240.0670.961±0.0351.4280.241 CG0.968±0.0320.957±0.037 CC0.972±0.0330.964±0.033 RGG0.966±0.0340.8540.4260.955±0.0322.3850.093 CG0.961±0.0320.958±0.032 CC0.966±0.0360.965±0.034基因SNP类型 基因型
续表

表5 AKR1C3、SHBG、SRD5A2 基因6 个SNP 位点基因型与不同性别2D∶4D 的关系(±s)
L 为左手,R 为右手。
基因SNP类型基因型女性2D∶4DF 值P 值男性2D∶4DF 值P 值SHBGrs727428LTT0.970±0.0331.0940.3360.961±0.0340.2390.788 TC0.965±0.0330.960±0.034 CC0.969±0.0270.957±0.043 RTT0.962±0.0350.1560.8560.959±0.0340.4870.615 TC0.963±0.0330.959±0.033 CC0.965±0.0300.955±0.031 rs3760213LGG0.968±0.0320.7060.4940.961±0.0361.9220.148 AG0.964±0.0320.954±0.036 AA0.972±0.0300.965±0.029 RGG0.964±0.0340.2070.8130.959±0.0320.3650.695 AG0.962±0.0330.956±0.035 AA0.962±0.0280.962±0.034 rs6259LGG0.968±0.0320.8980.4080.961±0.0361.4890.227 AG0.964±0.0320.955±0.036 AA0.976±0.0310.965±0.029 RGG0.964±0.0340.2900.7480.959±0.0320.2060.814 AG0.961±0.0330.957±0.034 AA0.965±0.0310.962±0.034
3 讨论
指长比常是判断胚胎起源成人性疾病(fetal origins of adult disease,FOAD)的宏观生物学标记[1,8]。近年来,国内外许多文献报道,指长比与精神分裂症[16]、不育症[17]、冠心病[18]及多种肿瘤[19]的发生存在相关性,这也使得指长比的研究成为探讨环境、遗传和行为等因素相互作用对FOAD 影响的热点之一。
2002 年,Manning 等[20]推测AR 基因可能会影响人类指长比的形成。AR 基因CAG 重复次数与男性右手2D∶4D 呈正相关[21]。之后,对基因多态性与指长比的关联性进行了一系列的研究,包括3 个不同人群的GWAS,分别发现LIN28B、SMOC1、FLI1、HOXD11/12 等基因/位点与指长比存在关联性,能解释3.8%的指长比形成差异[7]。这些基因主要集中影响指骨发育和性激素代谢途径。性激素代谢是非常复杂的过程,涉及许多关键酶,编码这些酶的基因多态性往往与其酶活性有关,由此影响指骨的发育与分化,最终影响2D∶4D 的形成[22]。因此,以性激素代谢相关基因为核心探讨指长比的形成成为目前的关注要点。
AKR1C3、SHBG 和SRD5A2 基因是雄激素合成代谢途径的关键基因,其可能通过影响雄激素合成代谢及雌-雄激素的相互转换,干扰胚胎期雌雄激素的比例,参与指长比的形成。本研究结果显示,AKR1C3 基因的rs12529 位点基因型、SHBG 基因的rs3760213 和rs6259 位点基因型和等位基因型频率虽然在不同性别间存在差异,但全部6 个SNP 位点基因型频率与指长比(2D∶4D)无关。
AKR1C3 是醛酮还原酶(aldo-keto reductase,AKR)超家族成员中催化效率最高的酶,位于雄激素合成代谢通路的终末端,能够促进雄烯二酮与睾酮的相互转化,促使雌激素转化为17β 雌二醇等[9]。既往研究发现,乳腺癌[23]、前列腺癌[9]、结肠癌[24]、胶质瘤[25]等多种激素相关性肿瘤均与AKR1C3 的异常高表达密切相关。同时,有研究[26]发现,前列腺癌、胃癌和脑瘤患者的2D∶4D 降低,而乳腺癌患者的2D∶4D 增高,因此,探讨AKR1C3表达与指长比的相关性,对于寻找肿瘤发生外在宏观生物学标记具有重要临床意义。rs12529 位于AKR1C3 基因第一外显子,为C-G 的错义突变。本研究发现,rs12529 各基因型频率存在性别差异,携带CG 基因型的男性多于女性。多项研究已证实,该位点CG 基因型及G 等位基因型可能作为保护性因素参与前列腺癌[27]、结肠癌[28]等的发生发展,这可能与其能够降低AKR1C3 表达水平进而抑制血清中前列腺特异性抗原(prostate-specific antigen,PSA)水平有关[29]。AKR1C3基因rs12529 与位于启动子区的rs1937845 位点处于强连锁不平衡状态,二者可能通过协同作用影响机体雄激素的代谢水平。然而,本研究并未发现rs1937845 位点基因型在不同性别间存在差异,这可能是由于AKR1C3 主要影响成人睾丸外雄激素产生水平的差异,对指长比的形成并无影响。
SHBG 作为一种运输类固醇激素的糖蛋白,对雄激素(如DHT)和睾酮的亲和力比对雌激素高5 倍,是雄激素转运的关键运输载体,通过与血液循环中的性激素特异性结合,发挥动态调节血液中性激素水平的作用[10,30]。rs3760213 和rs6259位点基因多态性被发现与血清SHBG 及TT 水平相关,携带rs3760213 A 等位基因及rs6259 A 等位基因的个体,血清SHBG 及TT 浓度上升[31]。rs6259 是SHBG 外显子8 中的错义SNP 位点,能导致第356 位密码子中的天冬酰胺被天冬氨酸取代(D356N),从而增加了一个额外的N-连接糖基化位点,降低SHBG 的血浆清除率,延长SHBG 的半衰期[32]。rs3760213 位于SHBG 基因第一内含子,可能通过影响SHBG 基因选择性剪切调节SHBG 表达水平。虽然在本研究中并未发现rs727428 各基因型分布存在性别差异,但通过单倍型分析发现,SHBG rs727428-rs3760213-rs6259(T-A-A)单倍型在男性中的比例高于女性,提示三者可能通过协同作用促进SHBG 的表达。由于SHBG 能显著调节机体雄激素代谢水平,而与SHBG 表达相关的多囊卵巢综合征和前列腺癌患者也被发现存在2D∶4D 下降的趋势[33],或许二者有一定关联。然而,本研究并未发现SHBG rs3760213 和rs6259 位点与指长比存在相关性。因此,本研究尚不足以认定SHBG 基因多态性与指长比无相关性,尚需在更大样本中,通过增加检测位点进一步验证。
SRD5A2 基因在雄激素合成过程中能够将睾酮转换为更具有活力的DHT,增强DHT 与AR的亲和力[34],是前列腺癌、尿道下裂、先天性肾上腺增生、小阴茎等男性化疾病的主要易感基因[35]。SRD5A2 基因rs523349(V89L)位点多态性可以导致第89 位密码子发生错义突变,由缬氨酸(Val)转变为亮氨酸(Leu),以此影响SRD5A2的表达活性。体外研究表明,rs523349 位点C 等位基因能够降低约40%的SRD5A2活性,这可能是其与晚期流产、转移性前列腺癌存在显著关联的原因[36-37]。然而,本研究并未发现该位点各基因型分布存在性别差异,同时与指长比无关。
综上,AKR1C3、SHBG 和SRD5A2 基因是雄激素合成代谢途径的关键基因,可能通过影响雄激素合成代谢及雌-雄激素的相互转换,影响指长比的形成。但是,本研究并未发现AKR1C3、SHBG 和SRD5A2 基因多态性与指长比(2D∶4D)有关,提示这些多态性位点可能主要影响个体出生后的性激素水平,对胚胎发育期形成的指长比没有贡献。由于本研究样本量较少且限于健康大学生群体,检测位点仅限于3 个基因的6 个多态性位点,对于探究雄激素代谢相关基因与指长比的关联性缺乏充足证据,尚需扩大样本及多态性位点,在不同人群体中完成进一步研究,以期为进一步揭示人类指长比形成的遗传学基础提供更多的理论依据。
