植物功能性状对半干旱黄土区生态系统多功能性的影响
2023-12-09董红霞杨智姣杨磊卫伟张钦弟
董红霞,杨智姣,杨磊,卫伟,张钦弟*
(1.山西师范大学 生命科学学院,山西 太原 030031;2.中国科学院生态环境研究中心 城市与区域生态国家重点实验室,北京 100085)
0 引言
黄土高原为我国典型的半干旱黄土区,气候干燥,土地退化严重,并且由于人为活动的加速运行,导致生物多样性下降以及生态系统结构逐步地简化,最终造成生态系统功能的迅速衰减[1]。因此,在植被恢复背景下加大对生态系统功能的研究,有助于植被恢复和生态建设。生态系统功能是通过生物群落来实现的生态系统所体现的各种作用[2]。近年来,随着对半干旱区生态系统功能研究领域的延伸,越来越多的研究表明生态系统功能与植物功能性状之间有着密切的关系,植物功能性状很大程度上会影响生态系统功能的变化[3-4]。植物功能性状是指在植物个体水平上通过影响植物生长、繁殖和存活能力,从而间接地影响其适合度的形态、生理以及生活史特征[5]。zhu 等在对黄土高原的研究中发现植物功能性状(如:叶面积,叶片抗拉强度,叶片冲孔力,以及根长,外根面,根体积等根系性状)对恢复草地的水土保持功能具有一定的影响[6]。Suding 等基于植物功能性状-影响框架,探究特定生态系统功能的影响性状(Effect traits)[7]即能够反映植物影响生态系统功能性的植物功能性状[8-9]中发现,叶片性状可决定资源捕获与利用及凋落物分解的速率,进而对生态系统功能产生影响[10];叶面积指数可用来指示植被的覆盖度,可通过影响蒸腾作用间接影响土壤水分[11];比叶面积大则不利于养分的储存,会增加光合速率、生长速率及叶片对养分的利用和储藏,往往具有更高的养分和水分吸收率[12]。诸多研究均表明,植物功能性状是研究植物群落影响生态系统功能的可靠手段[13-14]。目前大多数的研究都是植物功能性状与单独的一种或几种生态系统功能的研究[15],但随着对生态系统功能的研究,发现生态系统能够同时提供多种功能与服务即生态系统的多功能性(Ecosystem multifunctionality,EMF)。生态系统多功能性能够考虑植物功能性状如何同时影响多个生态系统功能,更有利于科学制定生态系统恢复规划和进行长期有效的可持续性管理[16]。但植物功能性状对生态系统多功能性的影响机制尚不明确。因此,在植被恢复背景下,亟待加强植物功能性状与生态系统多功能性关系的研究。
植物功能性状与生态系统功能关系的研究表明,植物群落主要通过“选择效应”(selection effect)和“生态位互补效应”(niche complementarity effect)两种机制影响生态系统功能。“选择效应”,认为优势种的性状很大程度上决定了植物群落对生态系统功能的影响[17]。通常使用物种的相对丰度或相对生物量的加权平均值来计算得到的植物功能性状的群落加权平均值( weighted mean)表征[5];而“生态位互补效应”则认为功能多样性越高,则物种间的资源利用的互补性越强,从而提高生态系统功能[18]。一般通过表示某一特定群落中植物功能性状值的大小、范围和分布状况的植物功能性状多样性(Functional diversity)[19-20]表征。基于植物性状,揭示植物群落影响生态系统多功能性的生态学机制及定量识别影响性状(effect traits),同样是制定恢复计划的关键所在[21-22]。
除此之外,研究过程中生态系统多功能性的量化方法的选择也是关键所在。目前,量化生态系统多功能性的方法有很多如:单功能法(single function approach)、平均值法(averaging approach)、功能物种替代法(turnover approach)和单阈值法(single threshold approach)。其中使用次数最多的是平均值法[23]。平均值法是将不同生态系统功能的测定值通过转化、平均从而得到一个能够解释生态系统功能平均水平的指数,该方法易于计算且容易解释,但缺陷在于默认一种功能的减少可以由另一功能来补偿。单阈值法主要是通过计算生态系统中达到选定阈值的生态系统功能数,最后得到一个指数,即为选定阈值下该生态系统多功能性的整体水平。此方法在功能之间存在权衡的情况下也不影响对多功能性的理解,可弥补平均值法的不足[24]。因此,根据已有研究,探究植被恢复背景下植物功能性状影响生态系统多功能性的机制,解析其核心影响性状,是制定半干旱区生态系统恢复计划和实现可持续管理的基础,也是目前急需解决的科学问题。所以,本研究采用平均值法和单阈值法两种方法量化生态系统多功能性,从植物功能性状出发,基于植物功能性状加权均值和功能多样性,探讨植物功能性状影响生态系统多功能性的选择效应和生态位互补效应,揭示在植被恢复背景下植物功能性状影响生态系统多功能性的主要机制,分析影响生态系统多功能性的主要植物性状,以选择合适的植被,因地制宜,为该区植被建设、生态系统可持续管理提供新思路,使该区域生态系统朝更高效、更可持续的方向发展。
1 研究区概况和研究方法
研究区位于甘肃省定西市巉口镇龙滩流域(104°45'—104°51' E,35°72'—35°75' N)(图1),2019 年正式加入中国生态系统研究网络(CERN)。海拔在1964 m~2212 m,流域总面积约16.1 km2,属于典型的黄土丘陵沟壑区[8]。气候为典型的半干旱气候,年平均降水量为386 mm,主要集中在7 月—9 月,且常以暴雨形式出现,蒸散量大,年平均气温为6.8 ℃,平均无霜期152 d[25]。该流域土质较均一,主要以黄绵土为主,有机质含量较低,土壤养分较为贫瘠。该流域为典型的草原地带,植被恢复比较典型,主要的人工恢复植被类型有:紫苜蓿草地(Medicago sativa)、柠条锦鸡儿灌丛(Caragana korshinskii)、油松林(Pinus tabuliformis)、山杏林(Armeniaca sibirica)[8]。天然植被中优势种主要以长芒草(Stipa bungeana)、赖草(Leymus secalinus)、大针茅(Stipa grandis)、百里香(Thymus mongolicus)等草本植物为主。
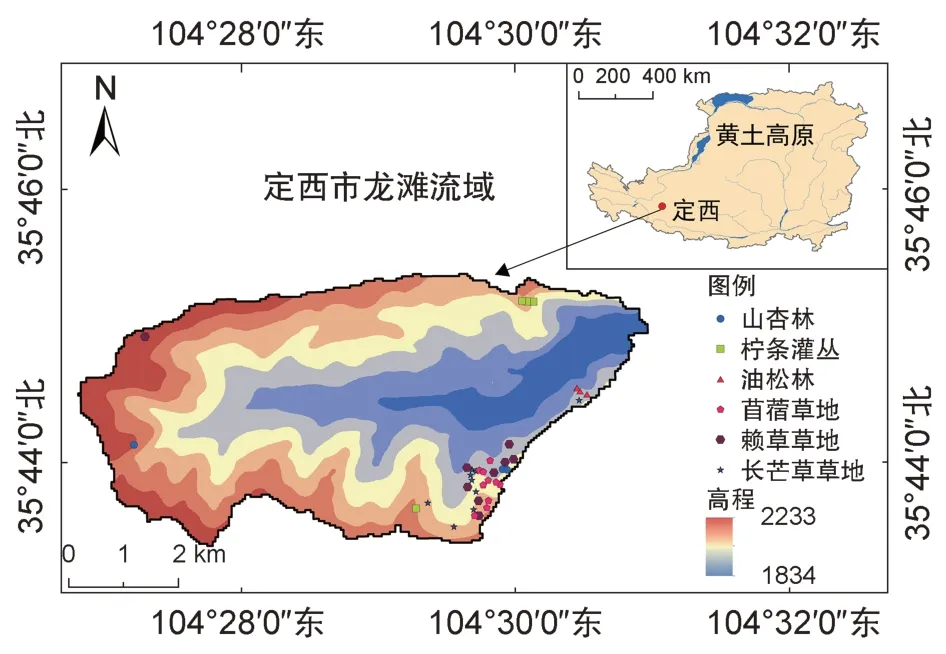
图1 研究区域及实验样点分布图Fig.1 Location of the study area and experimental samples
1.1 研究方法
1.1.1 样地设置与调查
本研究调查典型样地共38 个,包括长芒草草地、赖草草地、紫苜蓿草地、柠条锦鸡儿灌丛、油松林、山杏林6 种植被类型,其中,长芒草草地为区域代表性植被,未经人为干扰,属天然草地,赖草草地为自然恢复样地,紫苜蓿草地、柠条锦鸡儿灌丛、油松林、山杏林为人工恢复样地,各样地数量比为9∶9∶10∶4∶3∶3,并于2017 年8 月进行野外采样。其中,每个典型草地样地随机设置4 个1 m×1 m 样方;典型灌丛样地设置4 个5 m×5 m 灌木样方,且在每个样方内设置1 个1 m×1 m 草本小样方;乔木样地设置4个10 m×10 m 乔木样方,在每个乔木样方内设置1 个5 m×5 m 灌木样方和1 个1 m×1 m 草本小样方[22]。记录每个样方内出现的物种的名称及它们的高度、盖度、密度等基本信息,灌木和乔木样方测量灌木地径和乔木胸径等指标。坡向取正北方向为0°,按顺时针方向依次递增。
1.1.2 功能性状测定
根据样地的基本特征和之前在该区域对草地群落功能性状的研究[10]以及研究内容的需要,对每个样地选取叶长(LL)、叶宽(LW)、叶厚(LT)、单叶面积(ILA)、比叶面积(SLA)、叶干物质含量(LDMC)和高度(H)7 个指标来指示植物功能多样性。每种随机选取10 株左右用来测量它们的高度,根据植物叶片大小摘取每植株各方向的叶片15 片左右,草本叶片采集时,根据叶片特征采集的小叶片较多。Yaxin-1241 便携式叶面积扫描仪(北京雅欣理仪科技有限公司)测定各叶片的叶长、叶宽、叶厚、单叶面积,然后将叶片铺满叶面积仪,重复3 次测量植株总叶面积。随后将植株先放于105 ℃的恒温烘干箱中杀青30 min,最后在65 ℃的恒温下烘干至恒质量,记录烘干前的叶鲜质量和烘干后的叶干质量。
根据以下公式将物种水平的性状上推至群落水平,群落功能性状加权均值(Community weighted mean,CWM)表征“选择效应”,用于解释优势种的功能性状对生态系统多功能性的影响,功能多样性(Functional diversity,FD)反映“生态位互补效应”用于量化功能性状的变异性对多功能性的影响[26]。具体计算如下:
其中,Pij为物种i在样方j中的相对盖度,Tij为物种i在样方j中的测定性状均值,n为群落中物种数。
1.1.3 植物相关功能指标数据获取
各样方内均匀摘取来自各个方向的植物叶片,并取它们的根与茎装入信封,105 ℃杀青后65 ℃烘干研磨后测定植物全氮(PTN)、全碳(PTC)、全磷(PTP)、植物碳氮比(P C∶N)、植物氮磷比(P N∶P)。其中,植物全碳、全氮用Vario MAX cube 元素分析仪(德国Elementar 公司)进行测定,全磷采用钼锑抗比色法进行测定,最后取根茎叶均值来指示植物整体吸收状况。最后,草本层地上生物量采用齐地刈割法获取,灌木层则为先测定每丛高度、地径和分支数,选择3 株标准丛杀青、烘干至恒重称重,利用标准丛拟合相关关系进而推算单丛生物量;油松林通过测定树高和胸径,利用程堂仁的生物量模型进行估算[27];山杏林通过测定植株高度、胸径等指标,采用曾伟生的生物量模型进行估算[28]。
1.1.4 土壤功能指标数据获取
首先,在每个样方内用内径8 cm 的土钻取0~60 cm 土层的土壤,每20 cm 取1 次,分3 层,装密封袋后待测。其中,土壤全碳(STC)、全氮(STN)用Vario MAX cube 元素分析仪测定,速效氮(AN)用碱解扩散法测定,土壤全磷(STP)用NaOH 熔融—钼锑抗比色法测定,有效磷(AVP)用碳酸氢钠浸提后比色法测定,土壤有机质(OM)用重铬酸钾氧化-容量法测定。使用轻型人力钻在样地中进行0~5 m 深层取样,每0.2 m 进行一次取样,装袋密封后回实验室用来测定土壤水分。最后,用容积为100 cm3的环刀,在每个样方内取0~60 cm 的土壤,平均分成3 层,用来计算土壤总孔隙度(BP)。土壤pH 则使用FE20/EL20 型pH 计(中国 Mettler toledo 公司)进行测定。
1.2 生态系统多功能性
本研究共选取与地上初级生产力(地上生物量、叶面积指数)、植物生长策略(植物碳氮比、氮磷比、比叶面积)、植物养分吸收(植物全氮、全碳、全磷)、多样性指数(Gleason 丰富度指数、Shannon-Wiener 多样性指数、Pielou 均匀度指数)、土壤肥力(土壤全碳、全氮、速效氮、有机质、全磷、有效磷、土壤氮磷比)、水源涵养(土壤水分、毛管孔隙度)和营养物转化与循环(土壤总孔隙度、土壤碳氮比、土壤pH)相关的共23 个指标指示生态系统多功能性。基于上述生态系统功能,分别采用平均值法(Averaging approach,EMFA)和单阈值法(Single threshold approach,EMFT)两种互补的方法对生态系统多功能性进行评价。
平均值法首先对23 个功能指标用最小 —最大标准化方法进行标准化,然后计算标准化Z值,最后对Z值取平均,得到一个功能指数,具体计算过程如下:
其中,F为生态系统功能数,fi为生态系统功能i的测定值,ri是将fi转化为正值的数学函数,g是将所有生态系统功能转化为统一标准尺度。
单阈值法中最高的量化为5 个最高值取平均即取每个功能排名前5%的测定值的平均值作为该功能的最大值[23-24]。单阈值法中阈值的选择带有随机性,阈值的选择不同,得到的结果不同[29],阈值的具体确定方法见参考文献[23,29]。可根据研究目的的不同,选取不同的阈值来量化,本文选取20%、40%、60%、80%的阈值来量化不同水平生态系统多功能性[30]。以EMFT20%为例:
其中,EMFT20%为20%阈值水平下的生态系统多功能性,F为生态系统功能数,fi为生态系统功能i的参数,ri是将fi转化为正值的数学函数,ti是功能i的阈值(即功能i中的5 个最高值取平均值后与20%的乘积)。
1.3 统计分析
本研究主要运用Canoco 5.0 中的冗余分析对影响生态系统多功能性的植物功能性状加权均值与功能多样性进行分析。用筛选出的主要影响因子与生态系统多功能性(平均值法、单阈值法)进行回归分析,并用OriginPro 2016 进行具体数据分析及绘制图形。
2 结果和分析
2.1 功能性状加权均值与功能多样性对生态系统多功能性的方差贡献分析
功能性状加权均值与功能多样性对平均值法与不同阈值水平下的生态系统多功能性影响的方差分解结果如图2 所示。采用平均值法、20%、40%阈值法量化生态系统多功能性时,植物性状可以较好地解释生态系统多功能性,解释度分别为42.4%、38.2%、40.6%,而采用60% 和80% 阈值时,植物性状的解释度较低(<5%)。其中,功能多样性对单阈值法量化的生态系统多功能性中40%阈值的生态系统多功能性解释率最高,为14.9%;功能性状加权均值对平均值法量化的生态系统多功能性的解释率最高,为33.6%。以平均值法为例,功能性状加权均值能够解释多功能性的33.6%,功能多样性的解释率为8.8%。总体上,功能性状加权均值对生态系统多功能性的解释率总体高于功能多样性,植物性状主要通过选择效应影响生态系统多功能性,其次是生态位互补效应。
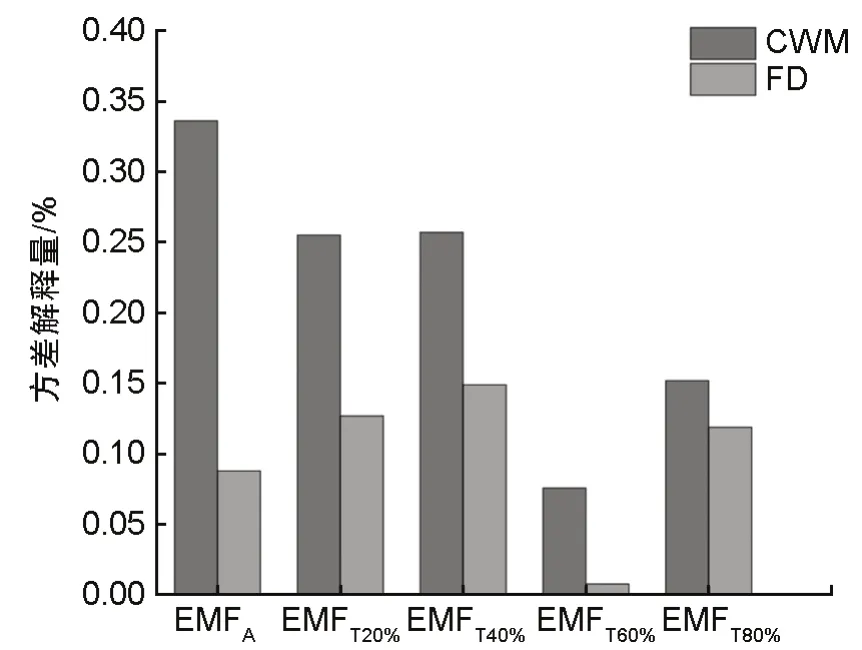
图2 功能加权均值与功能多样性对生态系统多功能性方差的贡献Fig.2 Contribution of functional traits weighted mean and functional diversity to variance of ecosystem versatility
2.2 功能性状加权均值与生态系统多功能性的相关分析
为筛选对生态系统多功能性影响显著的群落功能性状,对各功能性状加权均值与不同分析方法得到的生态系统多功能性,分别进行冗余分析(表1、图3)。第1 轴与第2 轴的解释度分别为24.31%、26.81%,冗余分析(Redundancy analysis,RDA)第1 排序轴与CWMLT、CWMLL、CWMH正相关性强,沿第1 轴从左往右CWMH、CWMLT、CWMLL、CWMLDMC逐渐增加,CWMLW、CWMILA、CWMSLA逐渐减小。第2 排序轴从下到上CWMH、CWMLT逐渐增加,CWMLDMC、CWMSLA变化不明显,CWMLL逐渐减小。最后,根据分析结果最终选取CWMH、CWMLDMC、CWMLT、CWMLL4个解释度高的指标与生态系统多功能性做回归分析。线性回归分析结果如图4 所示,CWMLT和CWMLL与EMFA呈极显著负相关关系(P<0.01),CWMLDMC与EMFT40%呈显著负相关关系(P<0.05),CWMH分别与EMFT20%、EMFT40%、EMFT60%呈极显著负相关关系(P<0.01)。

表1 各功能性状加权均值与生态系统多功能性关系解释的累计百分比Table 1 Cumulative percentage explained by functional traits weighted mean and ecosystem multifunctionality relationship
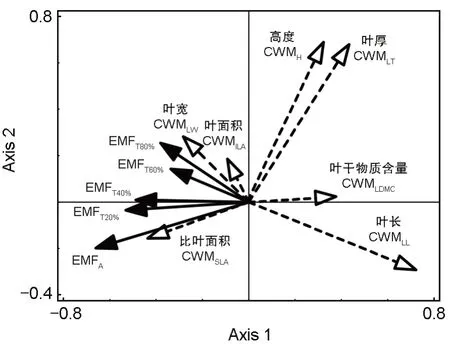
图3 各性状的功能性状加权均值与生态系统多功能性RDA排序图Fig.3 Redundancy analysis (RDA) ordination on the relationship between ecosystem multifunctionality and functional traits weighted mean of each trait
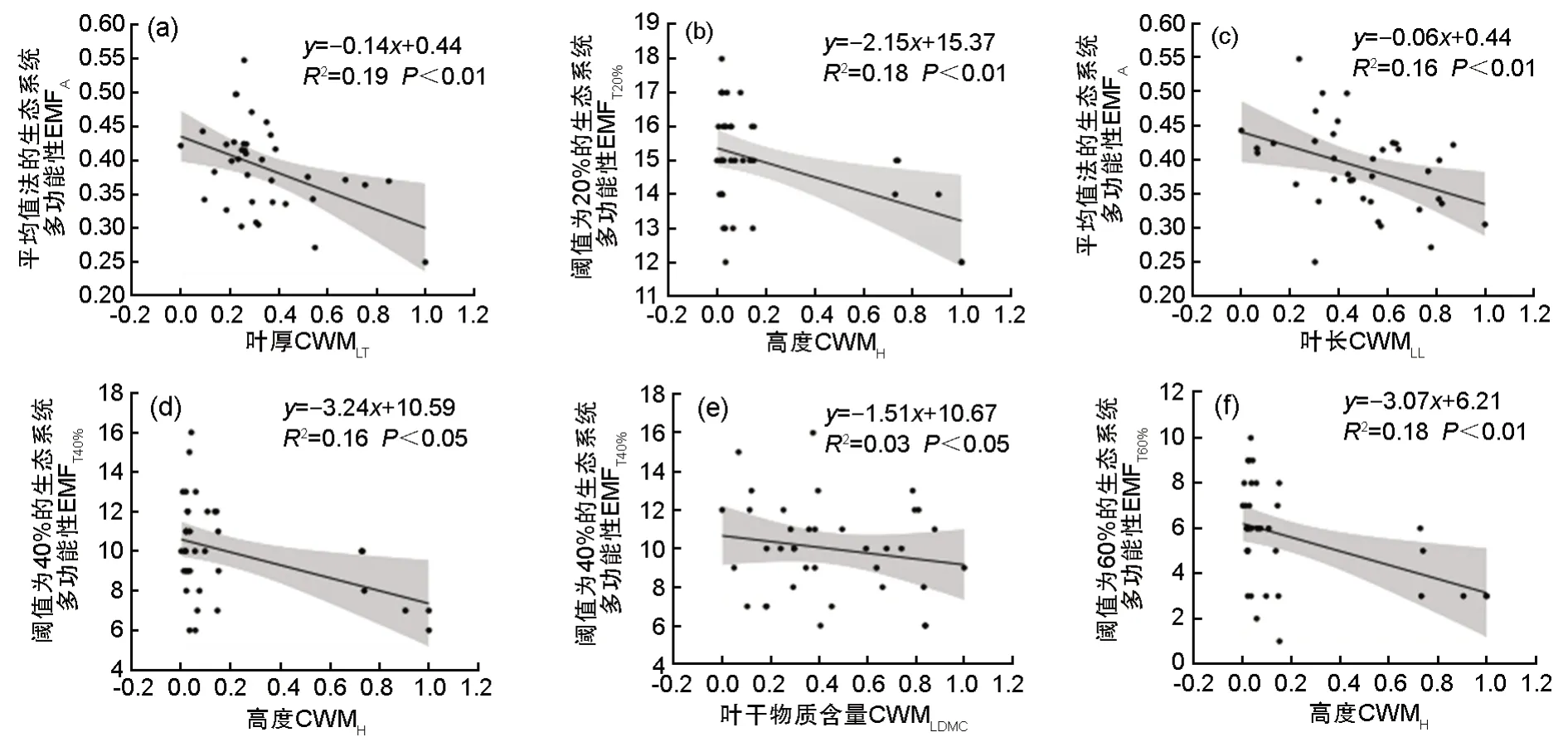
图4 群落功能性状加权均值与生态系统多功能性的关系(a)、(c) 图表示CWMLT、CWMLL与平均值法量化的生态系统多功能性的关系;(b)、(d)、(f) 图表示CWMH分别与阈值为20%、40%、60% 时生态系统多功能性的关系;(e) 图表示CWMLDMC与阈值为40% 的生态系统多功能性的关系。** 表示极显著相关(P< 0.01); * 表示显著相关(P< 0.05)Fig.4 Relationship between weighted mean of community functional traits weighted mean and ecosystem multifunctionality.(a) and (c) show the relationship between, CWMLT、CWMLL and ecosystem multifunctionality quantified by average method; (b),(d) and (f) show the relationship between CWMH and ecosystem multifunctionality at thresholds of 20%, 40% and 60%, respectively; (e) represents the relationship between CWMLDMC and ecosystem multifunctionality with a threshold of 40%
2.3 功能多样性指数与生态系统多功能性的相关分析
为筛选对生态系统多功能性影响最强的植被功能多样性,对各功能多样性指数与不同分析方法得到的生态系统多功能性进行冗余分析(表2、图5),第1 轴从左到右FDLW逐渐减小,其余各性状的功能多样性均逐渐增加。第2 轴的解释度较高,从下到上各性状的功能多样性指数均逐渐增加,FDLL、FDSLA的值最大。最后,根据分析结果最终选取FDLL与FDSLA2 个指标与生态系统多功能性做线性回归分析。回归分析结果如图6 所示,FDSLA与EMFA呈现显著正相关关系(P<0.05),FDLL分别与EMFT20%、EMFT40%呈极显著正相关关系(P<0.01)。

表2 各功能多样性与生态系统多功能性关系解释的累计百分比Table 2 Cumulative percentage explained by functional diversity and ecosystem multifunctionality relationship
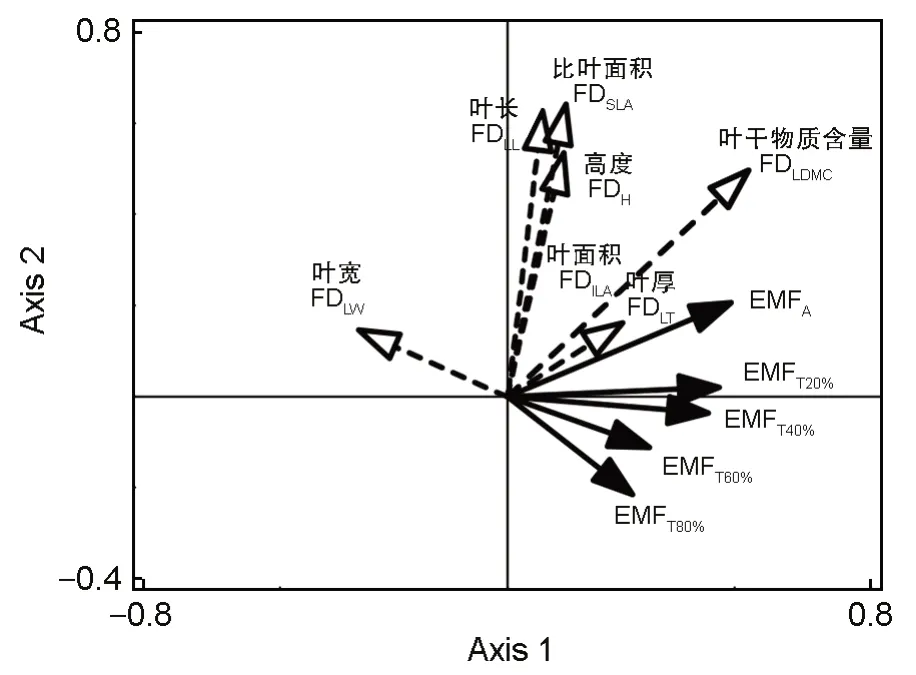
图5 各性状的功能多样性与生态系统多功能性的RDA排序图Fig.5 Redundancy analysis (RDA) ordination on the relationship between ecosystem multifunctionality and functional diversity of each trait
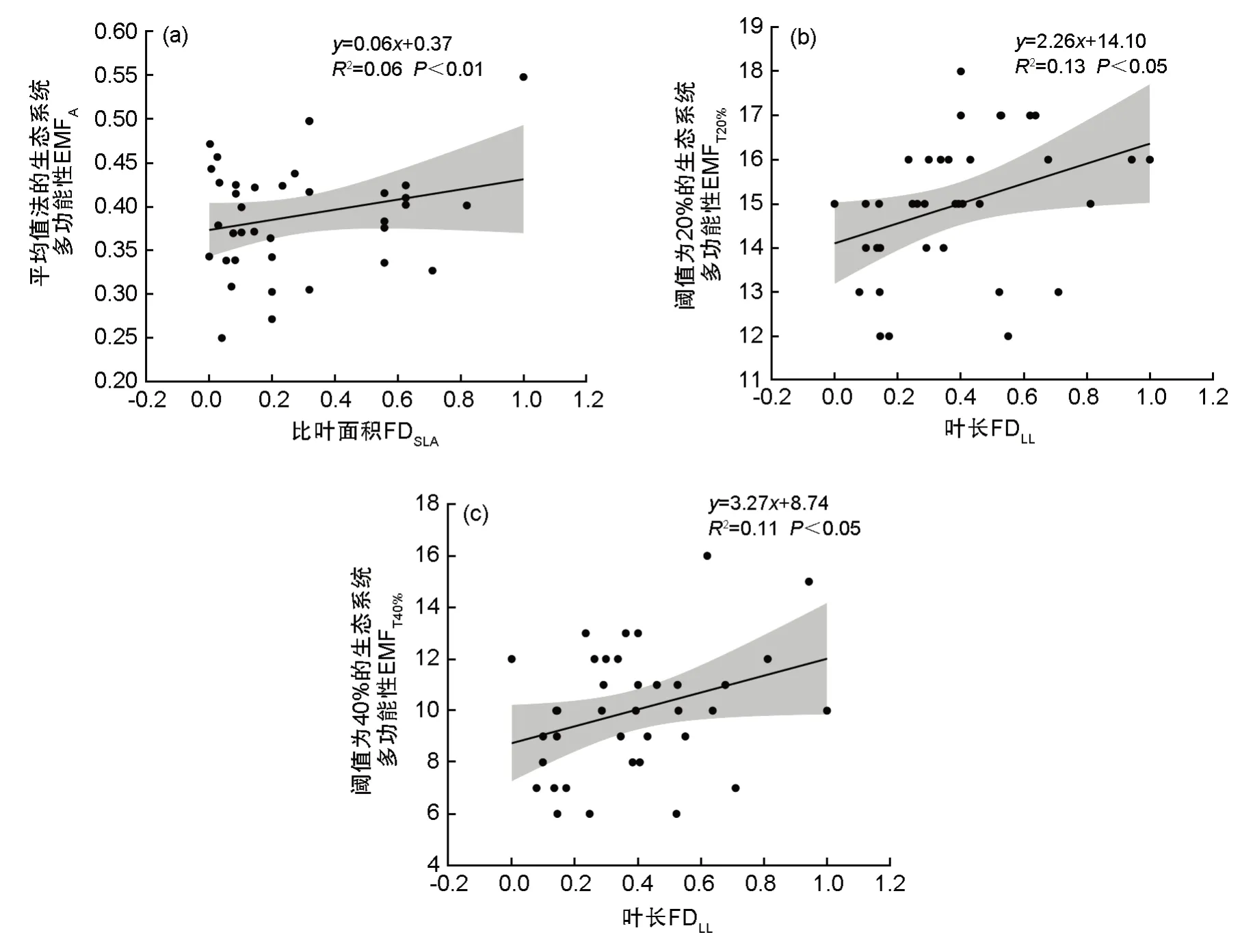
图6 群落功能多样性指标与生态系统多功能性的关系(a) 图表示FDSLA与平均值法量化的生态系统多功能性的关系;(b)、(c) 图表示FDLL分别与阈值为20%、40% 的生态系统多功能性的关系Fig.6 Relationship between community functional diversity indices and ecosystem multifunctionality(a) represents the relationship between FDSLA and ecosystem multifunctionality quantified by mean value method; (b) and (c)show the relationship between FDLL and ecosystem multifunctionality with thresholds of 20% and 40%, respectively
3 讨论
3.1 植物功能性状对生态系统多功能性的影响
植物功能性状决定植物的生存、生长、繁殖,能够对生态系统过程产生强烈影响。基于上述植物功能性状与生态系统多功能性关系的研究,对影响生态系统多功能性的选择效应与生态位互补效应进行方差分解。本研究得出在植被恢复背景下,植物性状主要通过选择效应影响生态系统多功能性,其次是生态位互补效应。值得注意的是,植物功能性状加权均值的解释率高于功能多样性指数,其主要是由于在逆境胁迫的环境下共生物种往往表现出相似的生存策略,表达相近的功能性状特征值以适应环境,此时应该支持选择效应。而在环境条件较为优越的情况下,物种竞争作用更强,生态位分化更为明显,生态位补偿效应更强[31]。其次,可能因为在自然生态系统中,功能多样性对生态系统多功能性的影响更依赖于生态位互补效应,但在人工恢复生态系统中,功能多样性对生态系统多功能性的影响更加依赖于选择效应[32]。有研究证实功能性状各种机制并不是单独对生态系统多功能性起作用,而是通过各个机制及相互作用影响多功能性[33]。一方面考虑到优势物种对生态系统功能的影响强大,可以证明优势种可用于创建功能修复系统,但更重要的是要考虑到不同功能之间具有权衡作用,没有单一物种可以维持多种生态系统功能,且生态系统功能会随着多样性的增加而增加,因此,可以结合选择效应和生态位互补效应两种机制,采用多物种组合维持多种生态系统功能[34]。
功能多样性对平均值法量化的生态系统多功能性解释度较高,主要由于平均值法最后是将各个功能值的均值作为多功能性指数,而且平均值法量化各个功能的权重,可以比较明确地评估多样性改变对生态系统多功能的平均影响[28]。功能多样性对单阈值法量化的生态系统多功能性的解释率中阈值为20%、40%的解释率高于阈值为60%、80%,可能是受阈值选取的影响[29]。随着阈值的升高解释率呈下降趋势,可能是由于功能多样性对生态系统多功性的影响受功能之间的交互作用的影响而导致[35]。功能之间的权衡(如生态系统的生产力和抗逆性不可能同时处于高水平状态)使得任何一个群落类型都不可能同时提供多种高水平的生态系统功能[35]。
3.2 植物功能性状加权均值对生态系统多功能性的影响
本研究从植物群落功能性状加权均值和功能多样性两个方面,研究植物功能性状对生态系统多功能的影响[36]。发现植物群落功能性状加权均值与基于平均值法、单阈值法的生态系统多功能性均呈极显著负相关关系[37-38],主要表现为植被高度、叶干物质含量、叶长与叶厚等指标。在对单一生态系统功能的研究中曾发现,在干旱地区对生态系统功能具有显著影响的因素为植物大小和叶片的性状[39]。本研究区样地包括长芒草草地、赖草草地、紫苜蓿草地、柠条锦鸡儿灌丛、山杏林和油松林共6 种植被类型,植被高度与多功能性呈负相关,这也与该区域中不同恢复方式对生态系统多功能影响的研究中林地多功能性较低相符合,其主要是由于较高的植株根系深且分布范围广,且维持较高的生产力也消耗较高的土壤水分,不利于维持较高的水源涵养功能。此外,有研究表明,高度较高的植被,其冠层汇集的雨滴能量较大,对地表的溅蚀作用增强[40],易引起该区域的水土流失,从而直接影响土壤肥力储存与水源涵养等生态系统功能。另外,有研究发现较高植物的氮矿化和硝化速率较低,导致土壤氮有效性降低[14]。
以往的研究结果表明具有较高叶长、叶宽、叶面积等叶形态指标的植株,可有效增加植被盖度,进而影响土壤溅蚀强度,有利于维持较高的土壤养分与水分储存能力,且枯枝落叶层的覆盖也可有效减少土壤侵蚀[41],增加土壤肥力。然而本研究结果表明只有叶长、叶厚与生态系统多功能性呈极显著负相关,可识别为样地所选功能性状加权均值中对生态系统多功能性的影响性状,叶宽与叶面积的相关性则不显著,可能是由于植株在特殊环境中所产生的适应环境的特征。研究区属半干旱区,土壤有效水分短缺,大部分通过自然降雨补充,因此,植物需通过调整叶片大小、叶长、叶厚、株高等植物性状来适应生存环境。根据之前我们在该区域的研究得出的植物功能性状对土壤有效水分的梯度响应规律[8],本研究结果可能是由于植物的生存环境对其的影响。除此以外,叶性状相关指标与地上初级生产力呈负相关关系,可解释为该区域植被为适应该地区干旱、强风等环境产生的性状趋同效应[42]。叶厚、叶干物质含量等均属于植物抗旱指标,在缺水状态下,植被通过减小叶面积,增加叶厚,增加叶片栅栏组织密度进而抵抗干旱,在这种环境下,植物往往具有较高的植物生长策略[43]。另外,叶厚、叶干物质含量也可影响凋落物质量,预测植被的分解速率[44],这是碳和营养物质生物地球化学循环中的一个重要环节,其与植被相对生长速率[45]和凋落物分解[46]呈反比例关系[47],不利于生态系统功能的发挥。
3.3 功能多样性指标对生态系统多功能性的影响
目前对功能多样性的研究,大多集中在与单一生态系统功能关系的探讨,发现功能多样性对生态系统功能的解释度较高[48-49]。进一步研究表明当同时考虑多个功能时,较高的植物功能多样对生态系统多功能性的提高更加重要[50]。在生态系统中,较高的功能多样性可能有助于获取更高的资源和具有更高的利用效率[51],并且也可以促进植物与土壤间的相互作用[52]。本研究中功能多样性与生态系统多功能性呈显著正相关关系,且主要表现为叶长和比叶面积,可识别为研究样地所选功能多样性指标中对生态系统多功能性的影响性状。叶长和比叶面积功能多样性值较高可能由于研究区属半干旱区,水土流失,土地退化,荒漠化有关,植物适应环境的特征。除此之外,可能由于人工恢复植被有较高的生产力和养分吸收能力。
本研究结果表明:功能多样性可能通过增加叶长和比叶面积增加旱地的多功能性;比叶面积多样性高表明该生态系统中存在不同的植物生长策略。前人研究表明,增加叶长的功能多样性可以导致植株规律的空间分布[41],从而维持较高的物种多样性[53],提高植物生长和生态系统功能[54];生长策略的差异可能对生态系统功能有强烈的正向影响,如生产力[55]、碳循环[56]和凋落物分解速率[57]。有研究表明,增加比叶面积可能会产生分解速率较高的凋落物[58],与本研究结果一致。除此之外,增加凋落物的功能多样性还可能会对凋落物分解速率[59]产生积极影响,这是维持和改善旱地多功能的潜在重要因子。总而言之,植被功能多样性的增加可能导致生态位互补,从而提高资源的利用和凋落物的分解,增加土壤生态系统功能[60]。此外,一些研究发现,较高的植物功能多样性还可以对微生物群落产生影响,这对维持生态系统多功能性也有重要作用[61]。
4 结论
本文主要基于植物功能性状加权平均值和功能多样性,探讨植物性状影响生态系统多功能性的选择效应和生态位互补效应。通过分析各植物功能性状与功能多样性对生态系统多功能性的影响得出:植物功能性状和功能多样性对不同的方法量化生态系统多功能性的解释度不同。总体上植物功能性状的解释度高于功能多样性,植物性状主要是受优势种性状的影响较大,通过选择效应来影响生态系统多功能性。通过对植物功能性状加权均值和功能多样性与生态系统多功能性的相关性分析,得出叶长、叶厚、叶高、比叶面积、叶干物质含量均与生态系统多功能性有显著性相关,可以认为是该研究区内生态系统多功能性的影响性状。因此,在该研究区,可根据影响性状选择合适的植物种类,筛选一些植被高度较低,叶子较长较厚的植被来构建植物群落,结合植物功能性状以及两种影响机制,从而提高生态系统多功能性,改善旱地生态环境。
