CaCl2、甜菜碱、5-氨基乙酰丙酸提升番茄耐低温弱光研究
2023-12-09苏常红寻雅雯宋子豪
苏常红,寻雅雯,宋子豪
(天津师范大学 地理与环境科学学院,天津 300387)
0 前言
低温弱光是设施农作物冬季和早春最具威胁性的灾害逆境,二者往往相伴发生。低温易破坏植物叶片叶绿素的合成,阻止卡尔文循环,光能转化速率下降,削弱植物光合作用;杨再强等研究表明,低温处理后,植物的光合速率、叶绿体色素含量、叶绿素荧光等参数均下降,且低温持续时间越长、温度越低,作物的光合作用受抑制越明显[1]。其次,低温条件下,细胞膜容易由液晶变为凝胶态,膜流动性变差,细胞质渗出,导致离子代谢紊乱;低温还导致酶活性下降,细胞内活性氧大量产生,与不饱和脂肪酸发生过氧化反应,生成丙二醛(MDA)[2-3]。光照是植物正常生产重要的环境条件,光通过多种信号途径调控植物生长代谢和形态的建成,导致植物的形态、生理、结构和生理代谢等发生一系列改变。光照不足会造成植物徒长,降低植物抗逆能力,弱光还导致植物体内类囊体膜和光合反应中心等光合作用机构受损,导致光系统I(Photosystem I,PSI)产生能量积聚,破坏叶绿体结构,影响植物光合作用[4]。朱雨晴等研究发现,遮阴虽然使番茄叶片叶绿素a、叶绿素b和类胡萝卜素含量上升,但导致光合速率下降[5]。低温弱光相伴发生情况下,对植物生长发育不利影响更为明显。潘东云等研究表明,低温弱光环境下,黄瓜幼苗叶片的光合速率、气孔导度和蒸腾速率均受不同程度抑制[6]。
番茄(Solanum lycopersicum L.)是茄科番茄属的一年或多年生植物,原产南美亚热带地区,是我国设施园艺的重要作物。番茄是一种喜温、喜光蔬菜,其最适宜生长温度为20 ℃~25 ℃;未经过低温锻炼的番茄幼苗,在低于5 ℃时,就会引起低温危害。番茄喜光,其最适宜的光照强度为30 000 LUX~50 000 LUX。光照不足的情况下,番茄易徒长,发育不良,后期坐果率降低,影响产量和品质。目前,对植物逆境胁迫几乎没有有效的措施,农艺措施改善植物生产环境虽有一定效果,但效果有限,费工费时,抗逆育种周期长,且植物遗传抗性特征很难和高产、优质等农艺性状同时出现。比较简单有效的抗逆栽培方式是指施用外源物质来提高作物逆境耐受力,改善作物逆境下的生长发育状况。研究表明,外源化学物质具有增强植物抗逆能力,与逆境下内源物质的特异性变化具有密切的联系[7]。目前针对番茄低温弱光研究主要集中于番茄逆境下材料的耐性筛选、鉴定及逆境伤害机理等方面,对外源物质对番茄耐低温弱光能力的研究及应用还比较少,外源物质影响番茄抗性机理方面报道得不多,影响了设施园艺作物生产技术的提升。
外源化学物质在植物抗低温方面影响的研究比较多,研究角度多为细胞膜稳定性、渗透调节物质、信号转导、抗逆基因表达等。Ca2+对植物抗低温具有增强效应,可以稳定细胞膜系统、增加细胞壁木质素和非纤维素的合成从而提高植物低温耐受。此外,Ca2+作为第二信使传导低温信号,诱导抗冻基因的表达等[8]。甜菜碱作为一种生物碱,在生物体内广泛分布。它含有一个由三个甲基组成的非极性的碳氢基团,既能与大分子亲水区结合,又能与疏水区结合,是一种高效的渗透调节物质,在植物受到低温胁迫时降低渗透势,减少渗透失水[9]。脱落酸(ABA)作为植物生长调节激素,具有调控植物气孔关闭及提升植物保护酶能力,防止膜脂过氧化,提高植物抗冷性[10-11]。多胺是一种低分子量、脂肪族含氮物质,是植物体内细胞分裂和发育的重要参与者,它可以和细胞膜上磷脂物质结合改变膜系统在低温下的通透性,减缓低温冷害期间细胞液pH 值的变化,减少乙烯的合成等,提高植物抗冷性[12]。此外,研究表明氯化胆碱和水杨酸也具有增强植物抗低温冷害的效果。前者能够提高叶绿素含量,促进类囊体膜光合作用电子传递,提高净光合速率[13],而水杨酸则主要作为逆境的信号分子,诱导植物的防卫反应[14]。磷脂是植物细胞膜系统的骨架成分,逆境下磷脂酶D(PLD)活性增加,催化磷脂降减。研究发现正丁醇、异丁醇等可以有效抑制PLD 活性,减少冷害发生[15]。针对弱光的外源化学物质研究相对较少,主要有5-氨基乙酰丙酸(ALA),作为一种含氧和氮的非蛋白氨基酸,ALA 存在于植物体活细胞中,与光合作用和呼吸作用密切相关[16]。此外,研究表明水杨酸(SA)也可以一定程度上增强植物耐弱光能力[17]。
前期研究以盆栽4 叶期番茄为试材,参照相关文献,从信号转导、抗过氧化、渗透调节等角度,并综合考虑简便有效、易获取和保存等原则,筛选抗低温弱光的备用化学物质。需要注意的是,虽然某些物质能够传递逆境信号,但由于可能参与其他反应,抗逆性并不一定总能得到增强,如磷脂酶Dα 虽然证明能够传递低温信号,由于其水解膜磷脂进而改变膜磷脂的组成,导致膜透性加大,抗寒力反倒可能降低[18]。某些低温信使物质有可能在弱光下其信号介导作用弱化[19]。此外,某些物质主要以内源物质发挥抗冷性,如多胺等[12]。选择了CaCl2(0.1、0.5、1、2.5、5 mmol/L)、ALA(10、20、30、40、50 mg/L)、甜菜碱(GB)(1、2.5、5、7.5、10、15 mmol/L)、氯化胆碱(5、0、100、300、500、700、1000 mg/L),正丁醇(0.25、0.5、0.75、1、1.25 mmol/L)、异丁醇(0.25、0.5、0.75、1、1.25 mmol/L)等作为备选物质。经筛选后,结果表明1 mmol/L 的CaCl2、2.5 mmol/L 甜菜碱、30 mg/L 的ALA 具有增强番茄幼苗耐低温弱光的能力[20],有必要进一步对外源物质增强植物耐低温和弱光的机理进行分析。本研究基于前期研究结果,尝试对三种化学物质耐单一低温和单一弱光的能力进行甄别,挖掘植物抗逆机理,同时为设施园艺生产防灾减灾及光温环境调控提供科学方法及参考依据。
1 方法
1.1 材料
选取番茄品种“中杂105”,将番茄种子浸泡在50 ℃热水中并向同一方向搅拌,水温降到30 ℃停止搅拌,继续浸种6 h。浸泡完毕后,将番茄种子放入垫两层湿润滤纸的培养皿中,保持滤纸湿润,30 ℃催芽48 h。将露白的番茄种子每孔一粒播种在128 孔的穴盘中,尽量保证番茄种子发芽程度一致。将草碳、蛭石、园艺土按体积4∶2∶1 混合,添加少量有机肥作基肥。将混合好的基质放入口径12 cm 的塑料花盆,均匀喷水保持湿润待用。穴盘育苗番茄长至一片真叶出现时,移入塑料花盆。两片真叶展开时,每盆间隔7 d 施两次250 mL 的日本园试营养液,分别用“吡虫灵”和“斑潜净”杀灭蚜虫和美洲斑潜蝇。
1.2 化学物质处理
番茄长至4 片真叶全展时,分组用配制好的1 mmol/L 的CaCl2、2.5 mmol/L 的甜菜碱(GB)、30 mg/L 的5-氨基乙酰丙酸(ALA)溶液对叶面正反两面进行喷施,每组5 个重复,对照组喷施清水。喷施时用移液管移取0.1 mL 洗洁精加入化学物质溶液,使溶液更好地附着于叶面。
1.3 单一低温处理
低温处理时要保证足够的光照,番茄苗白天(1 月10—12 日,上午9 时—下午4 时)放置室外自然温度和自然光照处理,晚上(下午4 时—第二天上午9 时)放置温室内,无光照。天气状况:晴天,白天最高气温6 ℃,夜间最低气温-3 ℃,中午最大光强1600 μmol·m-2·s-1。处理地址为北京市海淀区皂君庙路中国农业科学院东门实验站,东经 116.33°、北纬 39.97°。
1.4 单一弱光处理
弱光采取温室内遮阳网处理,中午遮阳网下最高光强为300 μmol·m-2·s-1,最高温度为30 ℃;夜间最低温度为18 ℃,无光照,处理时间为7 d。
1.5 指标测定
参照相关文献[1],分别于胁迫处理后2 h以及恢复5 d两个时间段测量生理指标和形态指标。
1.5.1 生理指标
测量叶片部位为第四片复叶顶端第一片小叶。测量的光合指标包括净光合速率、气孔导度、蒸腾速率。用Licor-6400 便携式光合仪进行测量,测量时采用人工光源,光强800 μmol·m-2·s-1。电解质外渗率(EC)采用外渗电导法,其原理为:质膜的选择透性因逆境伤害而改变或丧失,细胞液电导率与电解质外渗程度成正比,而与抗逆性成反比。过氧化物歧化酶(SOD)活性检测采用氮蓝四唑(NBT)光还原比色法测定。在氧气中,还原的核黄素与氧反应产生O2-将无色的氮蓝四唑还原为蓝色,SOD 通过催化O2-歧化,抑制蓝色形成,反应液蓝色愈浅,则SOD 酶活性愈高。丙二醛(MDA)测定采用硫代巴比妥酸(TBA)反应比色法,其原理为MDA 与TBA 结合,产生红色复合物三甲川(3,5,5-三甲基恶唑-2,4-二酮),通过532、450、600 nm 处吸光值差值测定MDA含量。脯氨酸的测定采用酸性水合茚三酮法,脯氨酸经磺基水杨酸提取后与酸性茚三酮反应,生成稳定红色产物,用比色法测定520 nm处吸收峰值测定脯氨酸含量,叶绿素的测定采用丙酮乙醇等量混合液比色法。
1.5.2 形态指标
结合Semeniuk 植株冷害形态标准和植物冷害分级法,设定叶片萎蔫指数,对植株受害形态进行分级。0 级,植物未受冷害,叶片正常。1 级,仅第一片真叶受冻,萎蔫区域大于50%。2 级,第一第二两片真叶有害,其中第一片真叶受害区域大于50%。3 级,第三片真叶受冻,其中第二片真叶受冻萎蔫区域大于50%。4 级,第四真叶呈现受冻状态,且第三片真叶的萎蔫区域大于50%。5 级,各叶片普遍受害,第四片叶受冻萎蔫区域大于50%。6 级,全株受冻死亡。冷害指数计算公式为:
式中S为每一级冷害的番茄植株数。
2 结果与分析
2.1 三种物质对番茄幼苗耐低温能力的影响
图1 看出,无论是低温处理后2 h,还是5 d 后,经3 种化学物质处理的番茄幼苗的净光合速率均显著高于空白对照组(CK),其效果依ALA、GB、CaCl2递增。恢复5 d 后差异进一步加大,尤其是1 mmol/L 的CaCl2处理的净光合速率最高,达7.255 μmol CO2·m-2·s-1,气孔导度也表现出了与净光合速率相似,低温处理后2 h 的气孔导度依ALA、CaCl2、GB 递增,恢复5 d 后,依ALA、GB、CaCl2递增,总体上以CaCl2处理效果最佳,为0.226 mol H2O·m-2·s-1(图2)。从冷害指数(表1)来看,无论是低温处理后以及恢复5 d 后,三种物质处理后的冷害指数均显著小于对照,30 mg/L 的ALA、2.5 mmol/L 的GB、1 mmol/L 的CaCl2处理的番茄幼苗冷害指数逐渐减小,都低于对照。纵向比较来看,恢复5 d 后,植株冷害指数加大,其原因是水渍状的叶片进一步萎蔫干化;但靠近叶柄的叶片变得更厚实(图3)。
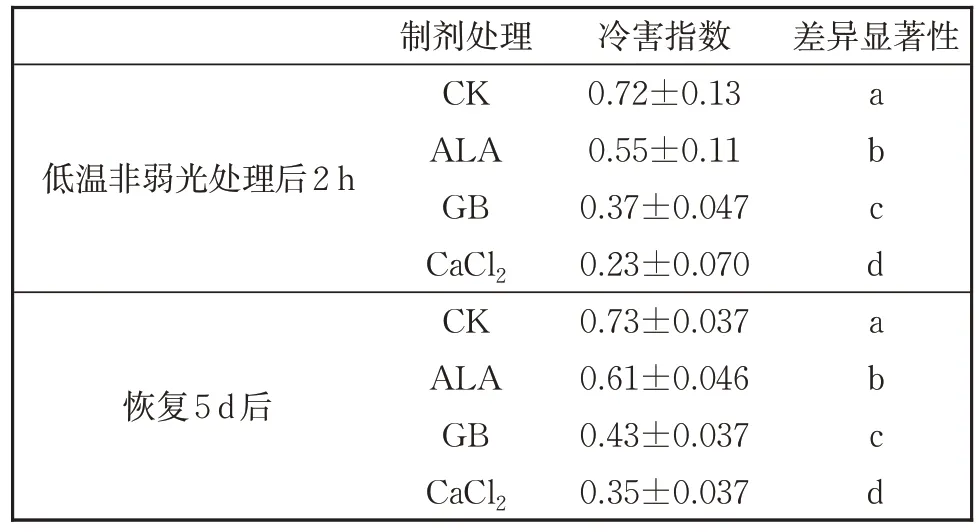
表1 三种化学物质处理并经单一低温处理后的番茄幼苗两次测量冷害指数Table 1 Wilting index of tomato seedling after treating with 3 chemicals and subsequent chilling treament
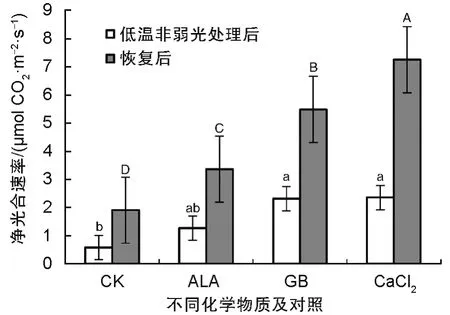
图1 不同化学物质处理低温非弱光番茄叶片净光合速率(Pn:Photosynthesis Rate)Fig.1 Pn (Photosynthesis Rate) of tomato seedlings after treating with different chemicals and subsequent chilling without low light

图2 不同化学物质处理经低温非弱光番茄叶片气孔导度(Cs:Stomatal Conductance)Fig.2 Cs (Stomatal Conductance) of tomato seedlings after treating with dirrerent chemicals and subsequent chilling without low light

图3 三种化学物质喷施后经低温处理恢复5 d后的番茄幼苗Fig.3 Tomato seedlings after 5 days recovery after treatment of 3chemicals and subsequent chilling
2.2 三种化学物质处理对番茄幼苗耐弱光能力的影响
图4 表明,无论是弱光处理后2 h,还是恢复5 d 后,三种化学物质处理的番茄幼苗净光合速率显著高于对照,尤其是ALA 处理的幼苗净光合速率表现最好,为0.15 μmol CO2·m-2·s-1。从图5 可以看出,弱光处理2h,GB 处理的番茄叶片气孔导度最高,恢复5 d 后,ALA处理的番茄叶片气孔导度显著高于其他处理,达到0.147 mol H2O‧m-2‧s-1,其次是GB 处理幼苗叶片气孔导度,为0.118 mol H2O‧m-2‧s-1。CaCl2处理的幼苗叶片气孔导度与对照无差别。从表2 中SPAD 值(相对叶绿素含量)可以看出,不同化学物质处理后的番茄幼苗的SPAD值均高于CK,其中ALA 的SPAD 值最高,其次是CaCl2和GB;恢复5 d 后,各处理的SPAD 值与弱光处理后的情况基本相同,只是ALA 与CaCl2处理的差异程度减少。
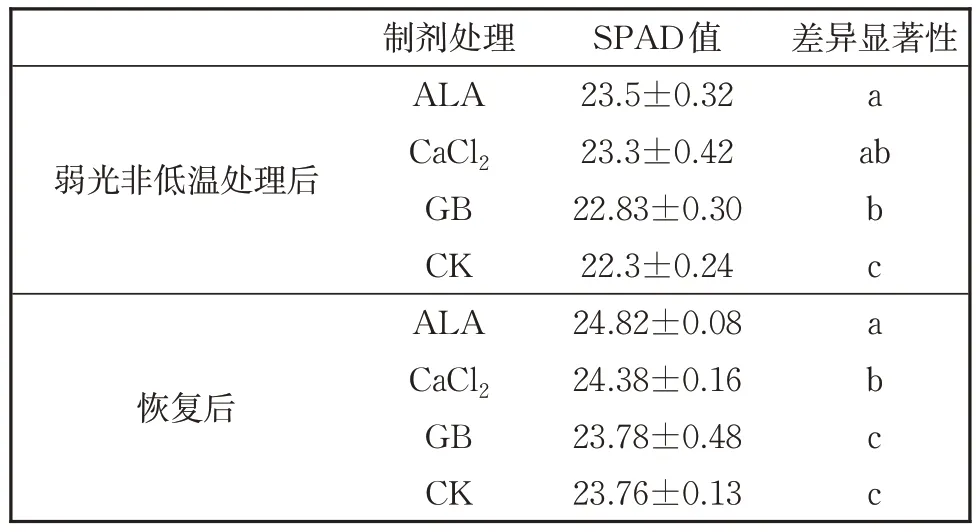
表2 几种化学物质处理后经单一弱光处理番茄幼苗SPAD值Table 2 SPAD value of tomato seedling after treating with 3 chemicals and subsequent low light treatment
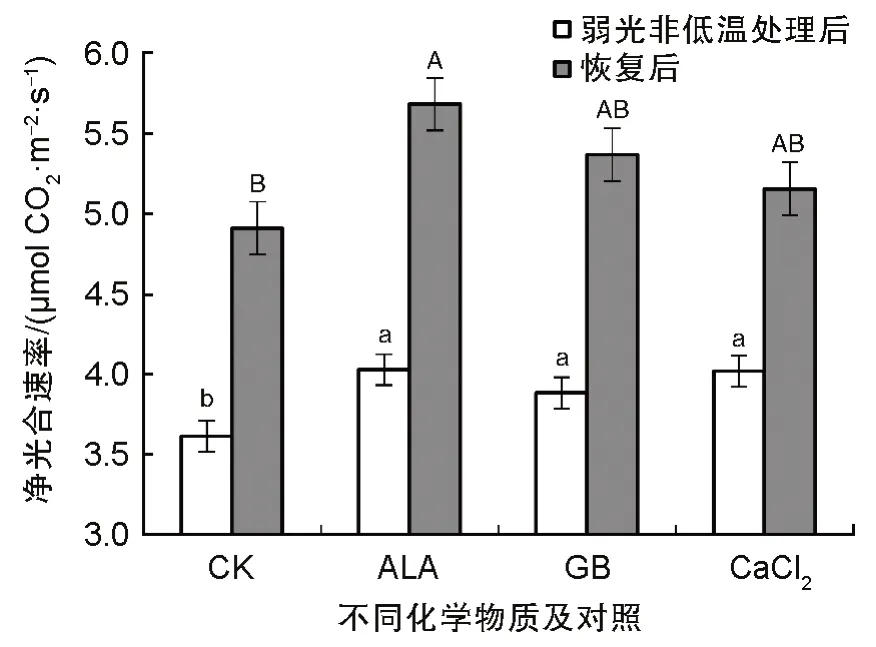
图4 化学物质处理经弱光非低温后番茄叶片净光合速率(Pn:Photosynthesis Rate)Fig.4 Pn (Photosynthesis Rate) of tomato seedlings after treating with different chemical and subsequent low light without chilling

图5 化学物质处理并弱光非低温处理番茄叶片气孔导度(Cs:Stomatal Conductance)Fig.5 Cs (Stomatal Conductance) of tomato seedlings after treating with dirrerent chemical and subsequent low light without chilling

图6 三种物质处理后并经弱光后番茄幼苗Fig.6 Tomato seedlings after treatment of 3chemicals and subsequent low light
2.3 三种化学物质处理对番茄幼苗增强耐弱光的机理
表3 表明,经30 mg/L ALA,2.5 mmol/L GB、1 mmol/L CaCl2处理的植株叶片电导率显著低于对照,说明这三种物质能够增强番茄叶片细胞膜在低温弱光下的稳定性,其中以1 mmol/L CaCl2效果最好。上述三种物质处理后番茄叶片的SOD活性明显高于对照株叶片,说明三种化学物质能有效促进植物在低温弱光逆境下消除活性氧成分,其中CaCl2和GB 的效果更优。MDA 是膜脂过氧化产物,反映细胞膜受损伤的程度,经三种物质处理后,番茄叶片MDA 含量均显著低于对照,说明喷施三种物质能够减少植物细胞膜的受损。经三种化学物质处理后,经胁迫的幼苗脯氨酸含量显著高于对照;脯氨酸是植物体内重要的渗透调节物质,可以保持原生质与环境的渗透平衡。从叶绿素含量来看,表3 中三种化学物质处理的番茄幼苗叶绿素含量高于对照,说明低温弱光下三种物质有利于植株光合能力的提升,其中尤以30 mg/L ALA 处理最为显著。此外表3 还表明三种化学物质处理会导致叶绿素a/b 值减小,即叶绿素b 的增长比叶绿素a 的增加更加明显。
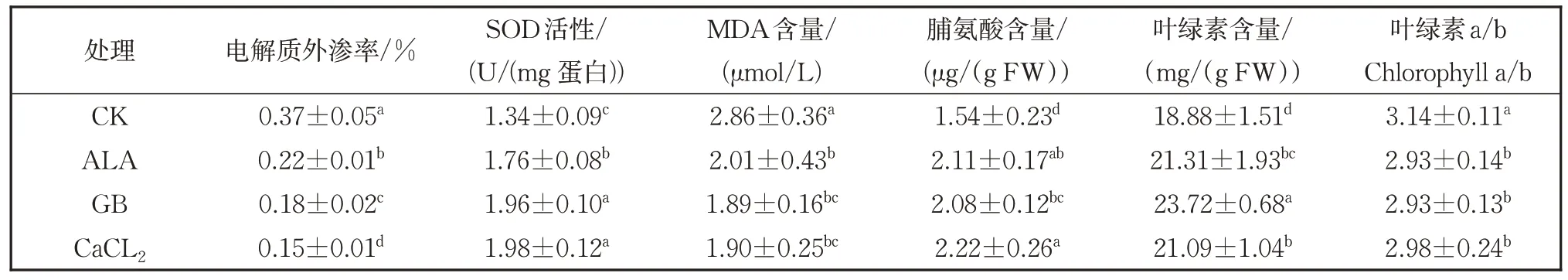
表3 不同化学物质对苗期番茄电解质外渗率、SOD活性、MDA、脯氨酸、叶绿素及叶绿素a/b的影响Table 3 The effects of different chemicals on electrolytes leakage, SOD activities, MDA, proline, chlorophyll and chlorophyll a/b of tomato seedlings.
3 讨论
植物为维持低温逆境下正常生理代谢,细胞体内发生一系列生理生化反应,内源物质会有一些特异性变化,而外源物质有可能与这些内源物质一道,提高植物抗逆能力,如逆境诱发钙离子的特异性变化,诱发钙信号与下游的钙结合蛋白进行转导,适应逆境胁迫。外施钙离子可能发挥与内源钙离子类似的作用诱发低温信号,促进抗逆性状的表达。此外,低温下,植物体内会积累一系列渗透调节物质,如甜菜碱(GB),能有效防止低温下细胞质的外渗。外施GB,在植物体中形成稳定复杂的蛋白膜,保护光合结构和酶的转录和翻译机制,作为分子伴侣干预低温等逆境胁迫导致的酶的折叠[21]。外源ALA 能够被动被植物吸收,补充内源ALA 的不足,促进叶绿素合成转化,改善植物的光合作用[22-23]。本研究表明,叶面喷施Ca-Cl2、GB 和ALA 三种化学物质,能显著提高苗期番茄的耐低温及弱光胁迫的能力,番茄净光合速率显著高于对照,冷害指数也显著低于对照。研究还表明,3 种化学物质对增强植物低温和弱光胁迫的贡献有所不同,Ca2+对于植物耐低温贡献更大,ALA 对增强植物耐弱光贡献更大。作为植物细胞体内重要的第二信使,Ca2+参与低温信号的转导,诱导植物的抗逆反应[24-26]。其次,Ca2+作为磷脂磷酸根和蛋白羧基连接的纽带,有稳定细胞壁和细胞膜双层磷脂结构,提高各种保护酶的活性[27]。ALA 可以有效促进植物叶绿素的合成,提高光合效率[28-30],对于维持植物光照不足情况的正常生长具有重要作用。GB 作为一种羧基季铵化合物,是植物体内重要的渗透调节物质,对多种逆境胁迫下细胞内保持低渗透压、维持水分平衡起作用,可以有效保护原生质免受逆境下的脱水胁迫[31-32],同时还具有提高逆境下膜保护酶活性和叶绿素含量[33-35]。低浓度的GB 能调节植物生长,提高植物抗性[36]。
此外,外源化学物质有可能通过改善代谢反应增强植物的抗逆性。本研究三种化学物质处理,经低温或弱光逆境胁迫后,番茄幼苗叶片气孔导度均高于对照,而气孔导度的增强对光合作用有促进效应[37-38]。另外,气孔导度增加引起蒸腾速率增强,而蒸腾作用是植物吸收和运输水分、矿质盐类和有机物质的主要动力,与干物质积累呈正相关,提升植物的抗逆能力。胁迫后恢复5 d 后的结果表明,3 种化学物质对于番茄幼苗的恢复作用非常明显。电解质外渗率(电导率)能够反映植物细胞膜遭受逆境胁迫后的破坏程度,是衡量植物遭受胁迫最直接的指标[39]。CaCl2、GB、ALA 三种物质处理后的番茄幼苗经低温弱光胁迫后,其电解质外渗率显著低于对照,尤以CaCl2的效果最好,说明外施这三种物质可以有效地增强植物细胞膜稳定性。SOD 是植物体内主要的过氧化酶,可以有效清除逆境胁迫下产生的活性氧[40-41]。三种物质处理后叶片SOD 活性高于对照,尤其是CaCl2和GB 处理最佳,说明外施三种物质可以增强植物清除活性氧的能力。MDA(丙二醛)作为膜脂过氧化的产物,一定程度上可以反映细胞膜受损伤的程度的大小[42],本研究二者呈显著负相关,显著系数达到-0.949 7。三种物质处理后番茄叶片MDA 含量均显著低于对照。
脯氨酸是植物蛋白组分之一,作为植物体内重要的渗透调节物质,可与细胞内化合物形成聚合物亲水胶体,保持原生质与环境的渗透平衡。此外,它以游离状态存在于植物体中,其亲水端可以与水分子结合,疏水端可以与蛋白质连结,有效防止酶和蛋白质因脱水而引起的变形[43-44]。本研究表明三种物质可有效地提高逆境下脯氨酸的含量。叶绿素含量能很好地反映植物光合能力的高低,三种物质处理后,逆境胁迫下的叶片叶绿素含量高于对照,尤其是ALA处理最为显著,其原因可能是由于ALA 作为卟啉化合物的前体,可以调节叶绿素和光敏素的合成,提高叶绿素和捕光系统II 的稳定性,对植物的光合作用具有重要作用[45]。此外三种物质处理后,叶绿素a/b 值有所下降,而叶绿素b 增加LHCP(捕光色素复合体)含量,提高光捕捉能力,另外对电子传递也具有重要意义[46]。
植物受到胁迫后,在长期的系统发育中形成了对外界的适应,抵抗和忍耐能力。即植物通过调整生命周期、改变形态和生理特征等方式来提升其抗逆性。从这个角度来说,植物抗逆性分逆境的规避、抵御和耐受三个方面。本研究只是对外源物质作用下植物逆境耐受性的增强效应进行了研究,对植物对逆境的适应机制未进行深入探讨。此外,植物诱导抗性的机理包括组织、细胞、分子结构与功能等多个层次,如植物在外界诱导激发下快速积累活性氧,引发局部过敏反应,产生防御信号进行传导,导致防御基因的表达,产生防御相关酶和次生代谢产物及相关蛋白抗性物质。本研究只是对个体尺度上植物抗逆性进行了研究,对抗逆基因及酶的表达、抗逆信号转导、抗逆基因及育种等尚未涉及,这也是本研究下一步需要拓展的方向。
4 结论
本研究表明,作为植物重要的外源调控物质,1 mmol/L CaCl2、2.5 mmol/L 甜菜碱、30 mg/L ALA 可以显著提高苗期番茄抗低温和弱光逆境的能力,无论在净光合速率、气孔导度、还是降低冷害受害性状方面,三种物质外观的表现均好于对照,其中1 mmol/L CaCl2增强苗期番茄抗低温效果最好,而30 mg/L ALA对增强番茄幼苗耐弱光能力更强。机理分析表明,三种物质能有效地增强植株抗氧化性,SOD 活性高于对照,作为过氧化产物的MDA含量也较低。喷施三种物质能有效地降低逆境下细胞膜的渗漏率,脯氨酸和可溶性糖等渗透调节物质保持较高的水平。此外喷施三种化学物质可以增加逆境下植物叶绿素含量,尤其是叶绿素b 含量,对于保护逆境下植物光合能力具有重要作用。
受条件限制,本研究植物抗逆机理只是采用了相对常规的指标,更深入的机理尚需进一步挖掘。在植物逆境胁迫分子机制及信号转导途径、植物逆境胁迫代谢组学研究,抗逆基因的表达及育种的应用、外源物质与内源物质如何协调等亟需探讨,这也是本研究进一步拓展的方向。