基于农艺性状的榕江茶核心种质构建
2023-12-05吴河饶任青艳陈思冶黄大玉陈涛林
吴河饶,任青艳,陈 莹,崔 叶,陈思冶,黄大玉,陈涛林
(贵州大学 茶学院,贵阳 550025)
茶树种质资源是茶树品种选育的物质基础,也是茶产业可持续发展的战略性资源[1]。榕江茶(CamelliayungkiangensisH. T. Chang)分布于贵州省榕江县月亮山一带,本课题组前期的主要茶品质化学成分检测结果显示,其混合样品茶多酚含量很高(33%),咖啡碱含量极低(0.10%),而作为咖啡碱合成前体的可可碱含量较高(3.77%),属于典型的“高茶多酚、低咖啡碱、高可可碱”类型的资源,而且加工红茶品质独特[2],在茶树新品种选育、茶叶新产品开发和茶叶深加工等方面具有很好的开发潜力和利用价值。然而,由于该野生茶资源群体较大,个体数量较多,对该资源的高效利用和保护构成了挑战。因此,合理构建该资源的核心种质,对其优异种质的发掘利用和高效保护具有重要的指导意义[3]。
Frankel[4]早在1984年提出核心种质的概念,即从原始种质中选取一部分能以最小资源数量最大程度代表整体种质遗传多样性的资源。近年来,核心种质构建的研究进展如火如荼,目前已在苹果[5]、花生[6]、玉米[7]等植物中构建核心种质。同时,核心种质的构建方法也在不断发展。李自超等[8]基于水稻表型数据构建出初级核心种质,利用生化指标和群体表型性状进行压缩得到二级核心种质,最后利用分子标记对二级核心种质再次压缩,完成了初级核心种质的动态更新,最终得到核心种质。此外,植物核心种质不再单一地进行构建,而是转向研究与利用的趋势发展,如水稻微核心种质产量相关的QTL定位[9]、甜菜重要农艺性状相关等位基因的开发[10]、中国大豆与国际大豆核心种质的遗传多样性比较[11]等。目前关于茶树种质资源鉴定和遗传多样性分析的报道较多,如蒋会兵等[12]、杨盛美等[13]和张明泽等[14]分别利用形态学、生化成分和分子标记等手段对茶树的遗传多样性进行了分析研究,但针对茶树核心种质资源构建的研究鲜有报道。
本研究以分布于贵州黔东南苗族侗族自治州榕江县月亮山境内的118份榕江茶种质资源为材料,在性状调查和品质成分测定数据整理工作的基础上,通过比较基于2种遗传距离(标准化欧氏距离、马氏距离)、4种聚类方法(离差平方和法、非加权组平均法、最长距离法、最短距离法)和7种总体取样规模(10%、15%、20%、25%、30%、35%、40%)排列组合构建56份备选核心种质,对备选核心种质进行有效性检验,最终获得榕江茶核心种质构建的最佳方案并构建其初级核心种质,并对构建的榕江茶核心种质进行评价和验证,为进一步研究榕江茶核心种质,高效收集、利用榕江茶种质资源提供理论依据。
1 材料和方法
1.1 试验材料
该研究以分布在贵州省黔东南苗族侗族自治州榕江县月亮山境内(平均海拔约1 200 m)的榕江茶(Camelliayungkiangensis)为对象。在实地考察的基础上选取具有代表性的单株资源121份,依地理位置分别编号为RJ001-RJ121。对每份单株资源进行GPS定位,对其整株和芽叶形态进行观测和记录。根据陈亮等[15]编著的《茶树种质资源描述规范和数据标准》规定的方法,于2022年4月中旬对118份榕江茶单株资源的形态特征进行观测记录,选取的性状主要包括树型、叶尖形状、叶片着生角度、叶基形状、叶身、叶面、叶缘、叶质、叶齿深度、叶齿锐度、叶齿密度、成熟叶颜色、树姿等13个质量性状,叶长、一芽二叶长度、叶宽、叶脉对数、叶面积、百芽重等6个数量性状,以及水浸出物、茶多酚、游离氨基酸、咖啡碱含量等4个品质指标,共计23个农艺性状。对每份资源的数量性状(叶长、叶宽、叶面积和叶脉对数)随机测量33张叶片,取其平均值;随机选取100个标准一芽二叶,其总重量记为百芽重,从中随机选取33个测其长度,取其平均值记为一芽二叶长;水浸出物、茶多酚、游离氨基酸等品质成分的含量分别按照相应的国家标准(GB/T 8305—2013、GB/T 8313—2018、GB/T 8314—2013)进行测定,咖啡碱含量测定参考王丽丽等[16]和杨金川等[17]的方法。一芽二叶的基本生化成分共计实测121株,其中数据齐全的单株资源共有118株。
1.2 数据的赋值标准
按照陈亮等[15]于2005年编著的《茶树种质资源描述规范和数据标准》规定的方法,对每份资源的质量性状均重复观测10次,按照等级数量编码方法进行分级赋值。
1.3 核心种质的构建方法
1.3.1 数据分组与聚类分析的方法
依据各榕江茶种质资源的叶齿锐度将118份种质资源划分为3组。参考前人总结出的层次聚类法进行系统聚类[19]。组内聚类得到聚类树状图,从遗传距离最近的组内,选择具有特殊农艺性状的1份材料,若组内无具有特殊农艺性状的材料则随机选择1份进入下一轮聚类,重复多次聚类,直至剩余材料总数达到每组所需的取样材料数。聚类程序和部分统计计算由SPSS 23.0和WPS 2019软件完成,图表由Origin 22.0和Adobe Photoshop 2022进行绘制。
1.3.2 构建策略的筛选
在分组和层次聚类分析的基础上,组内取样方法采用对数比例法(表1),在控制群体遗传冗余度的同时,适当增加稀有叶齿锐度的取样数量,以平衡叶齿锐度的取样[20]。对3个聚类分析基本环节进行分析,即两种常用遗传距离:欧氏距离(Euclidean distance)、马氏距离(Mahalanobis distance);4种聚类方法:离差平方和法(Ward’s method)、非加权组平均法(unweighted pair-group method with arithmetic means, UPGMA)、最长距离法(complete linkage)、最短距离法(single linkage);7种总体取样规模:10%、15%、20%、25%、30%、35%、40%。将不同的遗传距离、聚类方法和总体取样规模进行排列组合,共形成56种构建策略,分别对构建56份备选核心种质进行讨论。同时,采用表型保留比例(RPR)、变异系数变化率(VR)、极差符合率(CR)和Shannon-Weaver多样性指数(I)等4个评价指标比较备选核心种质优劣并选择最佳构建策略。
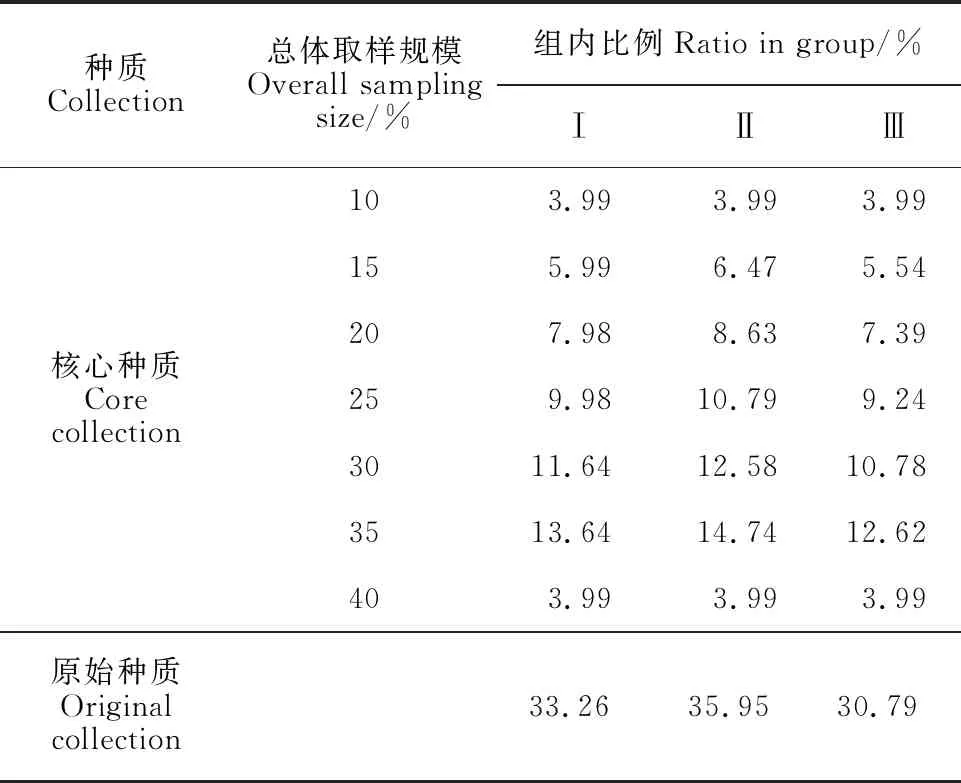
表1 对数比例策略在不同总体取样规模下的各组取样比例
1.4 核心种质的验证
利用获得的最佳构建方案构建榕江茶初级核心种质。对已构建完成的初级核心种质进行评价,采用特征值和评价指标检验核心种质有效性。
特征值检验利用原始种质和初级核心种质的23个性状的最小值、最大值、均值、标准差、变异系数、Shannon-Weaver多样性指数6个特征值检验核心种质的均匀程度和变异范围。评价指标检验利用表型保留比例(RPR)、变异系数变化率(VR)、极差符合率(CR)、最大值变化率(CRmax)、最小值变化率(CRmin)、平均值变化率(CRmean)、均值差异百分率(MD)和方差差异百分率(VD)8个评价指标验证初级核心种质有效性。同时对原始种质和初级核心种质的23个农艺性状进行t检验和F检验,验证核心种质对原始种质的代表性和差异显著性。特征值和评价指标参考刘遵春等[5]和刘娟[3]的公式计算。
2 结果与分析
2.1 总体取样规模的筛选
依据叶齿锐度将118份榕江茶种质资源划分为锐、中、钝等3组,组内取样方法使用对数比例法。分别计算56个备选构建策略核心种质的5个评价指标,即Shannon-Weaver多样性指数(I)、变异系数变化率(VR)、表型保留比例(RPR)、极差符合率(CR)和均值差异百分率(MD)。由表2可知,除10%总体取样比例的备选核心种质外,各备选核心种质的VR均高于107%,RPR均高于94%;I均高于1.12,CR均在92%以上。
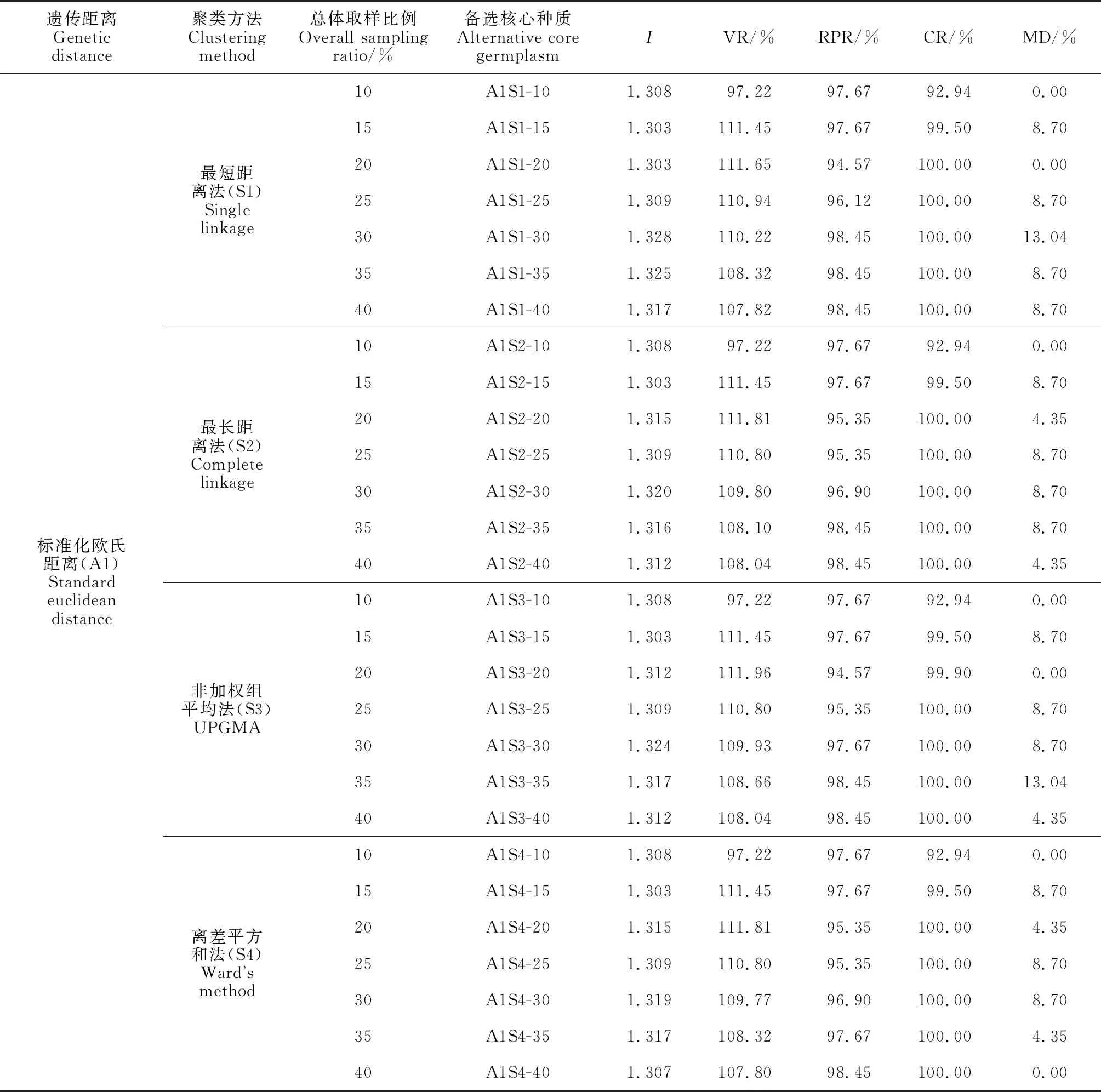
表2 不同构建策略的比较
从表2还可以看出,以欧式距离聚类的核心种质中,Shannon-Weaver多样性指数随取样规模升高表现先上升后下降的变化趋势,在总体取样规模为30%时达到最大值;而在以马式距离聚类的核心种质中,Shannon-Weaver多样性指数随取样规模升高表现的规律性不明显,总体取样规模在30%、35%和40%时,多样性指数较大,且三者间的差异较小。这说明低取样规模覆盖品种太少,不利于保存足够丰富且均匀的变异;随着总体取样规模的不断升高,核心种质的遗传冗余度也随之升高。
同时,核心种质的VR在总体取样规模为20%时均高于110%,以后随着取样规模的上升会逐渐降低;核心种质的RPR随着总体取样规模的升高呈递增趋势,在取样规模达30%时上升幅度趋于平缓;核心种质的CR在取样规模为10%和20%时普遍表现不佳;除A1S3-35、A1S1-30 2个核心种质外,其余核心种质MD均小于10%,未表现出明显规律性。
综合7种取样规模的5种评价指标,当总体取样规模达到30%时,核心种质拥有较高的遗传多样性指数,且能够在基本保留全部性状的前提下保持较高的VR和100%的CR,因此,30%的取样比例是榕江茶核心种质构建的最佳总体取样规模。
2.2 遗传距离的筛选
在确定取样规模为30%的基础上,采用标准化欧氏距离构建的核心种质,CR均为100%,VR均高于108%,Shannon-Weaver多样性指数(I)也能够保持在1.3以上,除A1S2-30、A1S4-30核心种质的VR略低于采用马氏距离构建出的核心种质A2S2-30、A2S4-30外,其余核心种质的Shannon-Weaver多样性指数、VR和RPR均优于以马氏距离所构建的核心种质(表2)。这进一步验证了标准化欧氏距离更适于处理含有较多不同类型性状的种质资源群体的观点[22]。
综上所述,以标准化欧氏距离构建的核心种质中,CR、VR和Shannon-Weaver多样性指数的结果更加优异,构建出的核心种质变异丰富且分布均匀,能够更好地代表原种质的遗传多样性,而VR与以马氏距离构建的核心种质有个别相对偏低,但其平均值相对较高,差异并不显著。因此在构建榕江茶核心种质时,采用标准化欧氏距离效果较好。
2.3 聚类方法的筛选
利用30%总体取样规模和标准化欧氏距离,比较4种聚类方法构建出的备选核心种质,结合表2发现,用最短距离法构建的核心种质表现最优,Shannon-Weaver多样性指数最高,变异系数变化率高达110.22%,极差符合率达100%,能够保留原种质全部极差,表型保留比例(98.45%)略低,但缺少的2个性状后续可以将其补充。与最短距离法相比,最长距离法和离差平方和法构建的核心种质的表型保留比例较低;而非加权组平均法构建的结果与其相近,但其Shannon-Weaver多样性指数(1.324)、变异系数变化率(109.93%)、表型保留比例(97.67%)均略低于最短距离法构建的核心种质。这与多年生作物中,最短距离法对白桦[23]核心种质构建效果最好的研究结果相吻合。
从全部56份备选核心种质来看,采用最短距离法、30%总体取样规模和标准欧氏距离构建的核心种质在能够保留全部性状的基础上,拥有最高的遗传多样性指数,相较于其他构建策略综合表现最好,在保证较少遗传冗余度的同时能够很好地代表原种质遗传多样性,是构建榕江茶核心种质的最佳策略。
2.4 核心种质的评价
采用标准化欧氏距离、最短距离法和30%总体取样规模构建初级核心种质(A1S1-30),在初选核心种质取样量上,人工定向补充2份具有特殊性状但未选入的种质资源,最终确定38份榕江茶资源组成核心种质,取样比例占原种质的32.20%。对其进行特征值和评价指标的计算,并与原种质进行比较(表3),可见核心种质与原种质23个性状的6个特征值相似性较高,且核心种质各性状的遗传多样性指数有一定程度的提升。
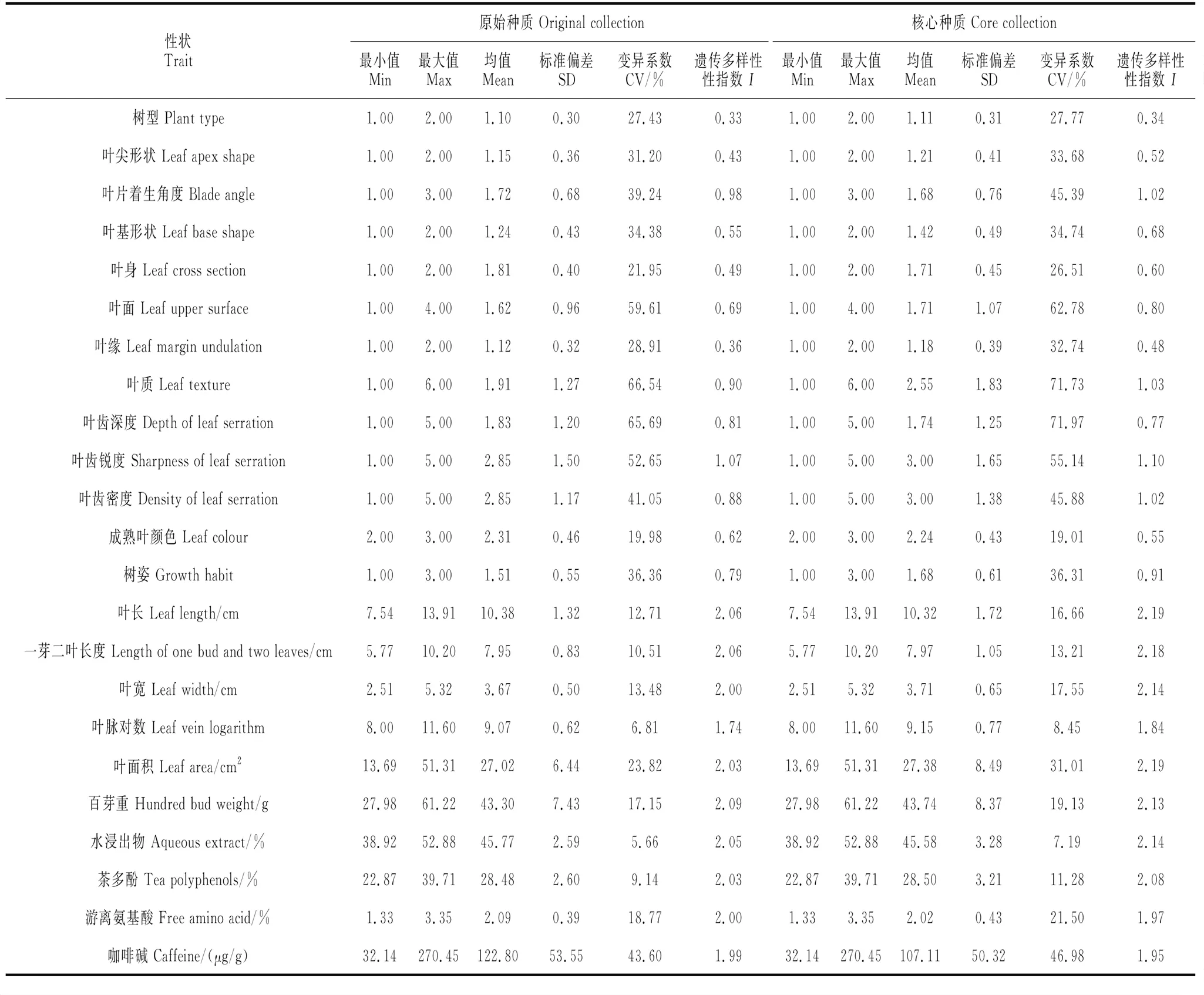
表3 榕江茶原始种质与核心种质性状特征值比较
同时,t检验结果(表4)表明,原种质与核心种质的叶基、叶缘、树姿3个性状具有显著差异,方差齐性检验结果表明原种质与核心种质的百芽重、一芽二叶长、叶缘3个性状具有显著差异。这主要由于叶缘、叶基2个性状中存在有较多的稀有性状,且分布极不均衡,百芽重、一芽二叶长和树姿3个性状的变异幅度较大,分布较为极端。核心种质在此基础上平衡了变异的分布,有效保留了原种质的遗传多样性。

表4 核心种质与原始种质各性状评价参数和t检验、F检验
另外,核心种质和原种质的主成分分析结果(表5)显示,原种质在第10个主成分时特征值开始小于1,第9个主成分的特征值为1.04,累计贡献率为71.99%;核心种质在第11个主成分时特征值开始小于1,第10个主成分的特征值为1.06,累计贡献率为82.27%。可见,核心种质的贡献率要高于原种质,有效地避免了核心种质的冗余。

表5 原种质和核心种质主成分分析的特征值和累积贡献率
此外,为了进一步对核心种质库进行确认,基于23个农艺性状主成分分析提取的第1、第2和第3主成分因子得分绘制核心种质和原种质的分布图(图1)。从图1可看出,补充种质进一步丰富了核心种质的分布范围,核心种质均匀分布在原始种质范围内,没有重叠现象,说明核心种质库去除了冗余,具有良好的代表性。

图1 核心种质和原种质的PCA分析Fig.1 PCA analysis of core germplasm and original germplasm
综上,构建完成的初级核心种质共包含38份榕江茶种质资源(表6),其5个评价指标VR、RPR、CR、MD、VD分别为113.90%、100%、100.00%、13.04%、13.04%,具有较高的遗传多样性指数和丰富的变异系数变化率,完整保留了原种质的极差和全部性状。榕江茶初级核心种质的极差符合率(CR)大于80%,且均值差异百分率(MD)小于20%,完全满足初级核心种质的构建指标[19]。
表6 榕江茶初级核心种质名录
3 讨 论
核心种质构建的结果很大程度上取决于对构建策略的选择。核心种质构建策略主要包括取样方法、取样比例、评价方法等,因不同作物的遗传构成和遗传分化不同,其代表性存在差异[23],应筛选出适合每种作物自身特点的方法策略进行核心种质的构建。由于计算和分类讨论的局限性,对构建策略中的所有因素进行分析是十分繁琐的。因此,本研究采用部分因素组合讨论的方法,针对不同的遗传距离、聚类方法和总体取样规模相结合构建榕江茶核心种质。本研究采用固定的组内取样方法,在固定因素中,随机取样策略虽可以保持原种质的遗传多样性形式,但随机取样是尽可能地获得原种质的无偏样本,而无法保留原种质有效的遗传多样性。而对数比例法在控制榕江茶种质资源群体遗传冗余度的同时,适当增加了稀有性状的取样数量。因此,本研究采用组内对数取样法的前提下进行后续分析,此方法已在辣椒(Capsicumspp.)[20]、切花小菊(Chrysanthemummorifolium)[21]等研究中取得了很好的应用效果。
因不同的材料在取样方式、生长环境和自身的遗传多样性等方面都存在一定差异。目前,总体取样规模没有统一的界定,多为原始种质的5%~40%[25]。因此,总体取样规模应当根据作物种类,原种质群体大小和群体组成具体分析。沈志军等[25]认为部分木本植物杂交周期长,核心种质要尽可能多地保留原种质的表型,取样比例要相应提高。由于本研究的实验材料数量较少,为了确保核心种质取样的合理性和系统性,设定10%为总体取样规模的最低值。经过综合比较7种总体取样规模后发现,30%总体取样规模的极差符合率均达100%,在此基础上拥有最高的遗传多样性指数和较高的表型保留比例,变异系数变化率基本稳定为108%~110%,综合表现最好。目前最为常用的遗传距离为马氏距离和欧式距离。但由于协方差矩阵在小规模核心种质构建中的问题,马氏距离并不适用于小规模核心种质构建[27]。本研究在构建核心种质时将性状数据进行标准化进而排除不同量纲的影响,通过对比4种评价指标后,发现标准化欧氏距离构建的榕江茶核心种质总体指标优于马氏距离,进一步验证了王建成[26]的观点;并与切花小菊[21]、豌豆(PisumsativumL.)[27]、毛花猕猴桃(ActinidiaerianthaBenth.)[28]等植物的研究结果相似。不同的聚类方法将植物分为不同类别,进而影响核心种质的代表性。李萌等[29]采用8种聚类方法构建高粱地方品种核心种质,结果表明,最长距离法构建效果优于其他7种。郎彬彬等[28]研究结果表明,类平均法构建毛花猕猴桃核心种质的效果最佳。在多年生植物中,最短距离法对白桦[22]核心种质构建效果最好。本研究中针对4种聚类方法进行了讨论,利用备选核心种质的评价指标衡量不同聚类方法的优劣,结果表明,采用最短距离法构建的核心种质在保持了原群体遗传变异的同时,使个体间的差异达到了最大化,多样性指数均值和变异系数变化率等方面优于其他方法,是构建榕江茶核心种质的最佳聚类方法。
农艺性状是植株基因与环境共同作用的结果,容易受环境影响,基于农艺性状数据构建核心种质库具有一定的局限性[31]。目前,利用分子标记技术构建核心种质是研究热点,如王倩等[31]利用高基元EST-SSR标记构建了黍稷(PanicummiliaceumL.)核心种质。童巧珍等[32]利用相关序列扩增多态性(SRAP)标记技术构建了15份百合核心种质。这些方法具有准确、高效、不受外界环境以及基因间互作影响等特点,能被很好地用来构建核心种质。因此,下一步将利用单核苷酸多态性(SNP)分子标记技术结合表型和生化品质成分进一步构建榕江茶核心种质,以期为有效利用其优异基因提供理论依据。同时,对核心种质、初级核心种质、保留种质等不同级别间的动态变化进行补充,提高核心种质有效性和实用性。
4 结 论
利用23个农艺性状数据对榕江茶核心种质进行构建时,以标准化欧氏距离、最短距离法和30%的取样规模结合进行逐步聚类是构建榕江茶核心种质的最适方法,最终得到38份核心种质,占原种质32.20%。核心种质能够很好地代表原种质的遗传多样性和遗传变异,为榕江茶种质资源的有效保存、利用奠定了理论基础。
