柑橘13个多胚品种同源四倍体高效发掘与分子鉴定
2023-11-21陈昊谢善鹏解凯东肖公傲周锐伍小萌吴群邓家锐敖义俊刘高平郭文武
陈昊 谢善鹏 解凯东 肖公傲 周锐 伍小萌 吴群 邓家锐 敖义俊 刘高平 郭文武


摘 要:【目的】基于柑橘珠心细胞存在自然加倍的特点,实生播种发掘千山红蜜橘等13个品种的同源四倍体。【方法】种子催芽萌发后实生播种,待幼苗长出3片以上真叶后,依据形态特征初选法“观根辨叶看油胞”从实生苗中筛选疑似多倍体,再通过流式细胞仪倍性分析与根尖染色体压片计数对疑似多倍体进一步鉴定倍性,并通过SSR分子标记分析鉴定所获多倍体的遗传来源。【结果】基于形态初选,分别从9个地方特色品种千山红蜜橘、八月橘、衢橘、早橘、扁平橘、瓯柑、狮头柑、冰糖橙、锦蜜冰糖橙和4个砧木品种磨坪香橙、日本香橙、枳雀、油皮金柑的343、499、892、385、519、290、457、241、119、690、828、114、129株实生苗中发掘获得2、1、3、2、7、3、1、3、1、3、17、1、2株疑似多倍体;用流式细胞仪对以上46株疑似多倍体进行倍性鉴定,获得45株四倍体和1株衢橘六倍体,采用根尖染色体计数法验证了上述结果;SSR分子鉴定表明,13个品种的45株四倍体所扩增条带大小与其相应二倍体亲本完全一致,推测其可能是由二倍体亲本珠心细胞自然加倍形成的同源四倍体。【结论】发掘的四倍体资源不仅丰富了我国柑橘多倍体类型,而且为我国柑橘无核育种和矮化广适砧木选育奠定了珍贵的材料基础。
关键词:柑橘;多倍体;流式细胞仪;SSR分子标记;无核育种
中图分类号:S666 文献标志码:A 文章编号:1009-9980(2023)11-2297-10
Efficient exploration and SSR identification of autotetraploids from the seedlings of thirteen apomictic Citrus genotypes
CHEN Hao1, XIE Shanpeng1, XIE Kaidong1, XIAO Gongao1, ZHOU Rui2, WU Xiaomeng1, WU Qun3, DENG Jiarui4, AO Yijun4, LIU Gaoping5, GUO Wenwu1*
(1National Key Laboratory for Germplasm Innovation & Utilization of Horticultural Crops, Huazhong Agricultural University, Wuhan 430070, Hubei, China; 2Chenzhou Institute of Agricultural Science, Chenzhou 423000, Hunan, China; 3Quzhou Academy of Agricultural and Forestry Sciences, Quzhou 324000, Zhejiang, China; 4Chenggu Fruit Technology Guidance Station, Chenggu 723200, Shaanxi, China; 5Huangyan Fruit Tree Technology Promotion General Station, Taizhou 318020, Zhejiang, China)
Abstract: 【Objective】 Citrus is a crucial part of Chinese fruit crops. There are abundant citrus germplasm resources in China, but many excellent local varieties are gradually eliminated by the market due to the problem of numerous seeds within the fruit. The fruits of triploid plants are generally seedless because of their sterile male and female gametes. Therefore, triploid production is a promising strategy to breed seedless cultivars in citrus. Triploids can be obtained by interploidy crossing between diploids and tetraploids. However, the tetraploid germplasm is rare, which limits the application of this strategy. Exploration of tetraploids is an important prerequisite for triploid production with the aim to improve the seedy local cultivars in our country. For the rootstock improvement, tetraploid plants are also valuable resources because of their higher metabolite content, and better resistance than their diploid parents. In this study, we planned to explore tetraploid plants from 13 local cultivars in our country by using the traits of spontaneous doubling of the nucellar cells in polyembryonic citrus varieties. The exploration of tetraploids from the above 13 local cultivars will not only provide excellent tetraploid parents for the production of triploid plants, but also lay the foundation for the basic research about the effect of genome duplication on some important trait change, such as dwarfing, extensive adaptivity and higher medicinal value in tetraploids. 【Methods】 After the mature fruits were harvested, the seeds were extracted and the seed coats were peeled off, and then they were placed in a thermostat to accelerate germination. When the seeds germinated, they were sown in pots and cultivated in a plant growth chamber. After the seedlings grew up with three or more leaves, putative polyploids were screened according to the morphological feature showing lower height, shorter taproots, less lateral roots, thicker and rounder leaves and declined oil gland density. The ploidy levels of these putative polyploids were further confirmed by flow cytometric analysis and the observation on root tip chromosome numbers. After determination of the ploidy level, some morphological traits, including plant height, root length and diameter, lateral root number, stem diameter, leaf thickness and shape index of the tetraploids and their corresponding diploid parents were measured at the same developmental stage. SSR analysis was used to identify the genetic origin of the explored tetraploids with at least three pairs of SSR primers selected for each cultivar. 【Results】 The polyembryonic degree of seeds from each cultivar was firstly determined and it showed that the seeds of all 13 cultivars were polyembryonic. Among them, Qu tangerine had the highest number of embryos with an average of 9.4 embryos per seed and Bingtang sweet orange had the lowest number of embryos with an average of 2.2 embryos per seed. Based on the morphological trait screening, we identified 2, 1, 3, 2, 7, 3, 1, 3, 1, 3, 17, 1 and 2 putative polyploids respectively from 343, 499, 892, 385, 519, 290, 457, 241, 119, 690, 828, 114 and 129 seedlings of Qianshanhong tangerine, Bayue tangerine, Qu tangerine, Zao tangerine, Bianping tangerine, Ougan tangerine, Shitougan, Bingtang sweet orange, Jinmi sweet orange, Moping Xiangcheng, Japanese Xiangcheng, Zhique and Youpi kumquat. After further confirmation of ploidy levels concerning above putative tetraploids, we obtained 45 tetraploids and one hexaploid plant from Qu tangerine, with an average occurrence rate of 0.85%, among which the rate of Japanese xiangcheng was the highest with 2.05% and the rate of Bayue tangerine was the lowest with 0.20%. The exploration time from seed germination to obtaining tetraploid seedlings varied among cultivars, with the longest time (42 days) used in Youpi kumquat and the shortest time (23 days) in Shitougan. The morphological traits of tetraploids and their corresponding diploid seedlings from nine cultivars of Qianshanhong tangerine, Qu tangerine, Zao tangerine, Bianping tangerine, Ougan tangerine, Bingtang sweet orange, Moping Xiangcheng, Japanese Xiangcheng and Youpi kumquat were measured. For plant height, tap root length, lateral root numbers and leaf thickness, the tetraploid seedlings of seven cultivars showed significant differences with their diploid parents. For taproot and stem diameter, only the tetraploid seedlings explored from Bingtang sweet orange and Japanese Xiangcheng had significant difference with their diploid parents. For leaf shape index, the tetraploid seedings from Bianping tangerine and Moping Xiangcheng exhibited significant differences with their diploid seedlings. In conclusion, most tetraploid seedlings of all nine cultivars showed lower plant height, shorter and thicker taproot, less lateral root number, thicker and rounder leaves than those of their diploid parents. These results provide supports for the screening of putative tetraploids based on morphological trait observation. For analyzing the genetic origin of the tetraploids obtained in this study, at least three SSR markers were used in each genotype. The results showed that the bands of all 45 tetraploids were identical with those of their corresponding diploids, indicating that all the 45 tetraploids might originate from the spontaneous chromosome doubling of nucellar cells of their corresponding diploids. In addition, the bands of the hexaploid from Qu tangerine were also identical with their diploid parent. We speculated that it might derive from chromosome doubling of a triploid zygotic cell, which formed by selfing of a FDR-type 2n gamete with a normal n gamete, and both gametes were produced by Qu tangerine. 【Conclusion】 This study verified that morphological screening combined with flow cytometry ploidy determination and SSR analysis is an efficient approach to exploring polyploid seedlings from apomictic citrus. Using this method, 45 autotetraploid and one hexaploid plants were obtained from 13 apomictic citrus genotypes. These newly discovered tetraploids are potentially valuable for not only genetic improvement of some elite local citrus cultivars with seeds produced by triploids using interploidy hybridization, but also selection of the promising rootstocks with dwarf, multi-resistance and broad adaptability characteristics to improve the ability to resist various abiotic and biotic stresses.
Key words: Citrus; Polyploidy; Flow cytometry; Simple sequence repeat; Seedless breeding
收稿日期:2023-08-02 接受日期:2023-08-21
基金项目:国家自然科学基金项目(32172525);南宁市科技重大专项(20222061);浙江省黄岩区农业揭榜挂帅项目(JBGS2022-HYNY02);华中农业大学深圳营养健康研究院项目(SZYJY2022009);国家柑橘产业技术体系项目(CARS-26)
作者简介:陈昊,男,在读硕士研究生,研究方向为果树生物技术与种质创制。Tel:027-87287393,E-mail:chenhao99@webmail.hzau.edu.cn
*通信作者 Author for correspondence. Tel:027-87281543,E-mail:guoww@mail.hzau.edu.cn
柑橘是我国南方最重要的水果,在我国农村经济发展与乡村振兴中发挥了巨大的作用[1]。我国柑橘栽培历史悠长,种质资源繁多,但许多优良的地方品种因种子多而逐渐被淘汰[2]。无核是柑橘遗传改良的重要育种目标之一[3],三倍体是天然的不育类型,创制三倍体是实现柑橘果实无核化的有效途径[4]。利用二倍体与四倍体倍性杂交是培育柑橘三倍体的主要方法[5],但我国一些地方特色品种四倍体资源仍较为缺乏,限制了该育种策略的应用。因此,发掘柑橘四倍体是利用倍性杂交策略改良我国地方特色品种种子多等性状的重要前提条件。此外,与二倍体相比,同源四倍体大部分表现为器官更大[6]、代谢物含量更高[7]、抗性更强[8]和植株矮化或致矮[9]等特点,发掘或创制柑橘砧木同源四倍体,对培育矮化、广适和多抗砧木品种具有重要的产业应用价值。
柑橘属及近缘属大多数品种均具有多胚现象[10]。据报道,少数珠心细胞在发育过程中会发生自然加倍,形成染色体加倍的珠心胚萌发后进而形成同源四倍体(或称双二倍体),其基因型与二倍体完全一致[11]。利用这一特性,梁武军等[9, 12]通过露地实生播种结合叶片形态观察发掘获得了枳、香橙、早金甜橙等20余个砧木和柑橘接穗品种的同源四倍体。但是,种子露地播种易受季节和天气等环境因素影响,存在萌发率低且初选准确率不高的缺点。针对该问题,周锐等[13]对上述技术流程进行了改良,提出了“观根辨叶看油胞”发掘四倍体的方法,该方法采用种子催芽后生长室播种代替露地播种,综合叶片、根系和油胞等多组织形态特征(如四倍体相较二倍体根短粗且侧根少、叶片厚、叶形指数小、油胞密度低等)筛选疑似多倍体,极大提高了发掘效率。基于该方法,谢善鹏等[14]以常山胡柚等6个柑橘地方特色品种為材料,发掘获得了53株四倍体新种质,准确率46.7%,从播种到获得四倍体耗时平均38 d。张成磊等[15]利用该方法从1289株山金柑实生后代中发掘同源四倍体8株,准确率100%,证明了“观根辨叶看油胞”结合流式细胞仪倍性鉴定是获得柑橘多胚品种同源四倍体的便捷高效的策略。
千山红蜜橘、八月橘、衢橘、早橘、扁平橘、瓯柑、冰糖橙和油皮金柑是我国陕西、广东、浙江和湖南等地的优良地方特色品种,但均存在种子较多和食用不便的问题;香橙、枳雀和狮头柑除了作为优良的砧木资源外,还具有一定的药用价值。发掘上述材料的多倍体新种质,不仅能为柑橘三倍体无核种质创制提供优良的四倍体育种亲本,而且对培育矮化、适应性强的四倍体砧木和药用价值高的柑橘资源具有重要研究意义和实践价值。
1 材料和方法
1.1 试验材料
9个地方特色品种:千山红蜜橘(Citrus reticulata Blanco)、八月橘(C. reticulata Blanco)、衢橘(C. reticulata Blanco)、早橘(C. reticulata Blanco)、扁平橘(C. Depressa Hayata)、瓯柑(C. reticulata Suavissima)、冰糖橙(C. sinensis L. Osbeck)、锦蜜冰糖橙(C. sinensis L. Osbeck)和油皮金柑(Fortunella japonica Swingle);4个砧木品种:磨坪香橙(C. junos Siebold ex Tanaka)、日本香橙(C. junos Siebold ex Tanaka)、枳雀(C. wilsonii Tanaka)和狮头柑(C. speciosa Hort. ex Tseng)。
八月橘采自广东省深圳市;冰糖橙、锦蜜冰糖橙采自湖南省郴州市;狮头柑、枳雀、日本香橙和千山红蜜橘采自陕西省城固县;磨坪香橙采自湖北省秭归县;衢橘、早橘和扁平橘采自浙江省衢州市;瓯柑采自浙江省温州市;油皮金柑采自广西融安县。
1.2 实生播种及初选疑似多倍体
种子实生播种参考谢善鹏等[14]的方法,实生幼苗形态初选疑似多倍体参考周锐等[13]的方法。从成熟果实剥取种子后,用1 mol·L-1 KOH溶液浸泡3~5 min溶解果胶,用清水搓洗干净后剥去种皮,置于垫有湿纱布的玻璃皿于28 ℃恒温暗催芽3~5 d;待胚根长至0.5~1.0 cm时,播种于塑料钵并覆膜保湿,置于生长室(温度25±1 ℃,光照16 h)培养。待幼苗长出3片以上真叶后,依据形态特征初选法“观根辨叶看油胞”从实生苗中筛选出疑似多倍体。
1.3 倍性分析与鉴定
流式细胞仪(Cyflow space, Sysmex, Japan)倍性分析参考解凯东等[16]的方法。用一步法试剂盒(Sysmex, Germany)对样品进行裂解和染色后上机检测,以二倍体(2n=2x=18)植株叶片(荧光强度X≈50)为对照,分析待测样品倍性,若待测样品为三倍体(2n=3x=27),则其荧光强度X≈75;若待测样品为四倍体(2n=4x=36),则其荧光强度则为X≈100。植株根尖染色体压片计数参考Xia等[17]的方法。植株倍性确定后,统计各个品种四倍体的自然发生频率(四倍体株数/群体株数)。
1.4 四倍体幼苗形态指标测定
参考谢善鹏等[14]的方法对相同发育时期的四倍体及二倍体实生苗形态指标进行测定。形态指标主要包括植株高度、主根长度、侧根数目、根粗(根茎分界处向下1 cm处的根直径)、茎粗(根茎分界处向上1 cm处的茎直径)、叶片厚度和叶形指数(叶片长度/叶片宽度)。每项指标均随机取样测定3个生物学重复,每个品种二倍体及四倍体实生苗各测量9株。用Excel 2016对数据进行处理,并用t-test(p<0.05)对数据进行显著性分析。
1.5 植物基因组DNA提取和SSR分子鉴定
参考Cheng等[18]的方法提取基因组DNA,SSR分子标记参考谢善鹏等[14]的方法。SSR引物见表1,由北京擎科生物科技股份有限公司武汉分公司合成。PCR扩增体系为10 μL:2 × Rapid Taq Master Mix 5 μL,ddH2O 3.5 μL,DNA 1 μL,Primer F、R各0.25 μL(10 μmol·L-1)。PCR反应在ProFlex PCR仪(ABI, USA)进行,扩增程序:95 ℃预变性3 min,95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸10 s,35个循环,72 ℃延伸5 min,4 ℃保存。PCR扩增产物由全自动毛细管电泳(QIAxcel Advanced, Germany)分离,成像结果由仪器自带软件QIAxcel ScreenGel自动生成。
2 结果与分析
2.1 13个柑橘品种均为多胚品种
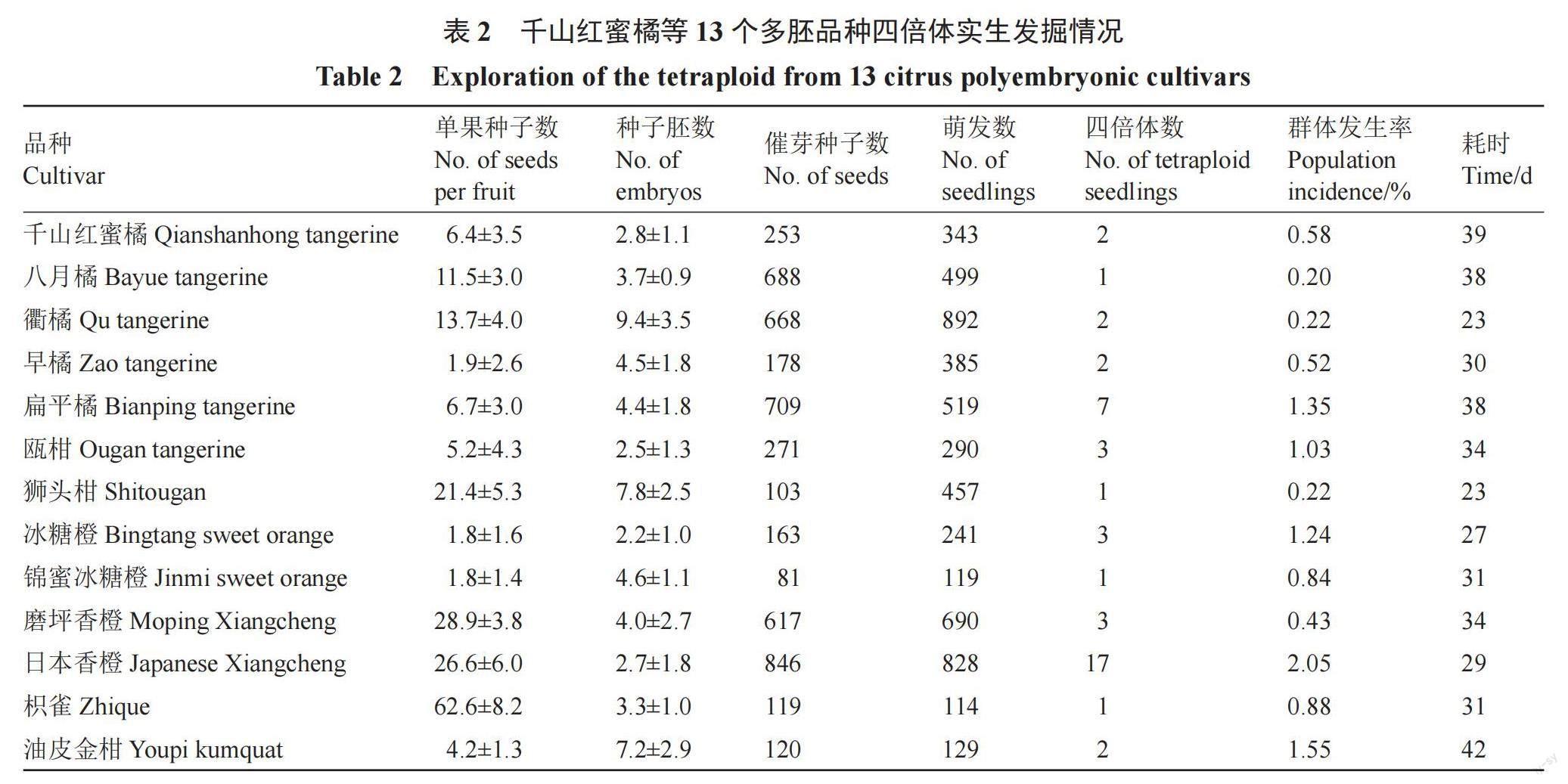
对千山红蜜橘、八月橘、衢橘、早橘、扁平橘、瓯柑、狮头柑、冰糖橙、锦蜜冰糖橙、磨坪香橙、日本香橙、枳雀和油皮金柑共13个品种的种子胚数进行调查(图1),每个品种至少统计20粒种子。结果表明(表2),上述13个品种均为多胚品种,种子多胚程度由低到高依次为冰糖橙、瓯柑、日本香橙、千山红蜜橘、枳雀、八月橘、磨坪香橙、扁平橘、早橘、锦蜜冰糖橙、油皮金柑、狮头柑和衢橘。其中,衢橘胚数最高,平均每粒种子9.4个胚;冰糖橙胚数最少,平均每粒种子2.2个胚。
2.2 实生发掘13个柑橘品种同源四倍体45株
依据“观根辨叶看油胞”形态学筛选法,分别从千山红蜜橘、八月橘、衢橘、早橘、扁平橘、瓯柑、狮头柑、冰糖橙、锦蜜冰糖橙、磨坪香橙、日本香橙、枳雀和油皮金柑13个品种的343、499、892、385、519、290、457、241、119、690、828、114、129株实生苗筛选获得2、1、3、2、7、3、1、3、1、3、17、1、2株疑似多倍体。用流式细胞仪对上述46株疑似多倍体进行倍性鉴定(图2),获得45株四倍体(千山红蜜橘2株、八月橘1株、衢橘2株、早橘2株、扁平橘7株、瓯柑3株、狮头柑1株、冰糖橙3株、锦蜜冰糖橙1株、磨坪香橙3株、日本香橙17株、枳雀1株、油皮金柑2株)和1株六倍体(衢橘)。进一步随机挑选日本香橙的二倍体和四倍体实生幼苗各3株,利用根尖染色体计数对其染色体数目进行鉴定,结果表明,流式细胞仪测得的四倍体染色体为36条,二倍体为18条,验证了流式细胞仪倍性分析的准确性。不同品种四倍体的群体发生率不同,发生率由低到高依次为八月橘、衢橘、狮头柑、磨坪香橙、早橘、千山红蜜橘、锦蜜冰糖橙、枳雀、瓯柑、冰糖橙、扁平橘、油皮金柑、日本香橙。其中,日本香橙群体发生率最高,为2.05%;八月橘群体发生率最低,为0.20%。从播种至获得四倍体植株,不同品种耗时23~42 d不等(表2)。
2.3 9个品种的四倍体与二倍体幼苗根、茎和叶片形态特征差异明显
对千山红蜜橘、衢橘、早橘、扁平橘、瓯柑、冰糖橙、磨坪香橙、日本香橙和油皮金柑9个品种发掘获得的四倍体及相应二倍体亲本实生幼苗的株高、主根长度、侧根数目、根粗、茎粗、叶片厚度、叶片长度和宽度等形态指标进行测定,以评价四倍体及其二倍体亲本实生幼苗的形态差异(图3)。结果(表3)表明,除冰糖橙和油皮金柑外,其余7个品种的四倍体幼苗株高显著低于其对应二倍体亲本;主根长度方面,除瓯柑和冰糖橙外,其余7个品种的四倍体幼苗主根长度显著低于其对应二倍体亲本;对于侧根数和根粗,除早橘和冰糖橙外,其余7个品种的四倍体幼苗侧根数显著低于其二倍体亲本,且仅有冰糖橙四倍体幼苗根显著粗于其二倍体亲本,其余品种根粗差异不明显;而在茎粗方面,大多数品种的四倍体幼苗与其二倍体亲本差异不明显,仅有日本香橙的四倍体茎显著粗于其二倍体亲本;在叶片厚度和叶形指数方面,除磨坪香橙和日本香橙外,其余7个品种的四倍体幼苗叶片厚度显著大于其二倍体亲本,扁平橘和磨坪香橙的四倍体幼苗的叶形指数显著小于其二倍体亲本,其余品种差异不显著。总的来说,大多数品种的四倍体幼苗与其对应二倍体相比,植株一般表现为株高显著降低,主根变短粗且侧根减少,叶片增厚、叶形指数变小等特点。
2.4 SSR分子鉴定
用19对SSR引物对从13个品种发掘获得的四倍体和六倍体植株进行SSR分子鉴定,每个品种至少选择3对多态性较好的引物进行鉴定(图4)。结果表明,13个品种的45株四倍体所扩增条带的大小与其相应二倍体亲本所扩增条带大小完全一致,推测其可能由二倍体亲本珠心细胞自然加倍形成的同源四倍体;衢橘六倍体条带也与其二倍体亲本基本一致,推测其可能由衢橘形成的2n配子与正常n配子自交形成三倍体合子后,再经染色体自然加倍形成。
3 讨 论
除枸橼类、柚类等以外,柑橘绝大多数品种的種子都具有多胚现象[10],即一粒种子萌发会产生多株幼苗,其中可能既有合子苗(有性苗)又有珠心苗(无性苗)。一方面,因合子胚在与珠心胚发育中营养竞争处于劣势地位[26],常导致合子胚败育,较少部分为有性杂种,很大程度上降低了杂交育种的效率;另一方面,由于珠心苗与母本遗传组成完全相同,属于无性后代,播种后的植株能保持母本优良性状,便于固定杂种优势[27],具有重要的育种应用价值。柑橘多胚品种珠心细胞自然加倍的现象被报道以来[5,28-30],大量多胚品种的同源四倍体资源被发掘[9,11-15]。特别是将植株外部特征(包括株高度、根长度和粗度、叶片颜色和厚度、油胞密度等指标)作为形态标记用于初选多倍体,使得柑橘多倍体发掘技术趋于完善,效率也得到很大提升,准确率超过80%[13]。本研究基于课题组前期建立的“观根辨叶看油胞”形态初选结合流式细胞仪倍性快速鉴定的方法,从千山红蜜橘、八月橘等13个柑橘多胚品种的5506株实生苗中发掘获得了四倍体45株和六倍体1株,不仅验证了利用“观根辨叶看油胞”形态观察初选柑橘多倍体的高效性,还在一定程度上丰富了柑橘多倍体种质资源,为未来更好地对我国柑橘地方特色资源进行无核改良或培育矮化、广适砧木奠定了宝贵的材料基础。
用SSR分子標记鉴定所获多倍体的遗传来源,13个品种的45株四倍体的扩增条带的大小与其相应二倍体亲本的条带完全一致,表明其可能由二倍体亲本珠心细胞自然加倍形成的同源四倍体,这与前人的研究结果基本一致[11-15]。而从衢橘中发掘获得的六倍体条带与其二倍体亲本条带也完全一致,未发现新的条带,推测其可能由衢橘产生的杂合型2n配子(第一次减数分裂异常产生,多数位点基因型与二倍体亲本一致)与其正常的n配子自交形成三倍体合子后再加倍形成,该假设后期可利用近着丝粒标记[17]或随机筛选更多的标记[31]对该六倍体进行基因分型以确定其遗传来源。与四倍体相比,关于柑橘六倍体的报道相对较少。梁武军等[32]通过三倍体葡萄柚胚培养获得了包括六倍体在内的一些不同倍性植株和疑似非整倍体植株。Guo等[33]通过细胞融合技术,创制获得了锦橙(二倍体)+HR(哈姆林甜橙+粗柠檬异源四倍体体细胞杂种)的异源六倍体。与其他倍性植株相比,六倍体植株生长速度极慢、植株极度矮小、叶片更小且厚、颜色更深,生产应用价值还需进一步评价。但是,作为基础研究材料,六倍体植株对开展柑橘倍性效应等相关基础研究具有重要价值。
4 结 论
通过“观根辨叶看油胞”形态学初选结合流式细胞仪倍性鉴定,快速高效发掘了千山红蜜橘等13个品种的45株同源四倍体以及1株衢橘六倍体,遗传鉴定显示所有四倍体均为其对应二倍体亲本的珠心细胞自然加倍形成的同源四倍体。这些四倍体新种质不仅为我国柑橘地方特色品种的无核改良提供了宝贵的四倍体育种亲本,同时也为选育矮化、多抗和适应性强的四倍体砧木奠定了丰富的材料基础。
致谢:本研究所用的磨坪香橙和油皮金柑果实分别由国家现代农业(柑橘)产业技术体系三峡库区脐橙综合试验站曹立新站长和柳州综合试验站陆文科站长提供,特此感谢!
参考文献 References:
[1] 郭文武,叶俊丽,邓秀新. 新中国果树科学研究70年:柑橘[J]. 果树学报,2019,36(10):1264-1272.
GUO Wenwu,YE Junli,DENG Xiuxin. Fruit scientific research in New China in the past 70 years:Citrus[J]. Journal of Fruit Science,2019,36(10):1264-1272.
[2] 解凯东,彭珺,袁东亚,强瑞瑞,谢善鹏,周锐,夏强明,伍小萌,柯甫志,刘高平,GROSSER J W,郭文武. 以本地早橘和槾橘为母本倍性杂交创制柑橘三倍体[J]. 中国农业科学,2020,53(23):4961-4968.
XIE Kaidong,PENG Jun,YUAN Dongya,QIANG Ruirui,XIE Shanpeng,ZHOU Rui,XIA Qiangming,WU Xiaomeng,KE Fuzhi,LIU Gaoping,GROSSER J W,GUO Wenwu. Production of Citrus triploids based on interploidy crossing with bendizao and man tangerines as female parents[J]. Scientia Agricultura Sinica,2020,53(23):4961-4968.
[3] XIE K D,YUAN D Y,WANG W,XIA Q M,WU X M,CHEN C W,CHEN C L,GROSSER J W,GUO W W. Citrus triploid recovery based on 2x × 4x crosses via an optimized embryo rescue approach[J]. Scientia Horticulturae,2019,252:104-109.
[4] NAVARRO L,ALEZA P,CUENCA J,JU?REZ J,PINA J A,ORTEGA C,NAVARRO A,ORTEGA V. The mandarin triploid breeding program in Spain[J]. Acta Horticulturae,2015(1065):389-395.
[5] OLLITRAULT P,DAMBIER D,LURO F,FROELICHER Y. Ploidy manipulation for breeding seedless triploid citrus[J]. Plant Breeding Reviews,2008,30:323-352.
[6] SUDO M,YASUDA K,YAHATA M,SATO M,TOMINAGA A,MUKAI H,MA G,KATO M,KUNITAKE H. Morphological characteristics,fruit qualities and evaluation of reproductive functions in autotetraploid Satsuma mandarin (Citrus unshiu Marcow.)[J]. Agronomy,2021,11(12):1-12.
[7] TAN F Q,ZHANG M,XIE K D,FAN Y J,SONG X,WANG R,WU X M,ZHANG H Y,GUO W W. Polyploidy remodels fruit metabolism by modifying carbon source utilization and metabolic flux in Ponkan mandarin (Citrus reticulata Blanco)[J]. Plant Science,2019,289:110276.
[8] WEI T L,WANG Y,LIU J H. Comparative transcriptome analysis reveals synergistic and disparate defense pathways in the leaves and roots of trifoliate orange (Poncirus trifoliata) autotetraploids with enhanced salt tolerance[J]. Horticulture Research,2020,7(1):1689-1702.
[9] 梁武军,解凯东,郭大勇,谢宗周,伊华林,郭文武. 10个柑橘砧木类型同源四倍体的发掘与SSR鉴定[J]. 果树学报,2014,31(1):1-6.
LIANG Wujun,XIE Kaidong,GUO Dayong,XIE Zongzhou,YI Hualin,GUO Wenwu. Spontaneous generation and SSR molecular characterization of autotetraploids in ten citrus rootstocks[J]. Journal of Fruit Science,2014,31(1):1-6.
[10] 張斯淇,徐强,邓秀新. 无融合生殖与柑橘多胚现象的研究进展[J]. 植物科学学报,2014,32(1):88-96.
ZHANG Siqi,XU Qiang,DENG Xiuxin. Advances in apomixis and polyembryony research in Citrus plants[J]. Plant Science Journal,2014,32(1):88-96.
[11] 谢善鹏,杨雯惠,陈昊,肖公傲,解凯东,夏强明,伍小萌,郭文武. 国庆1号温州蜜柑珠心胚苗培育及四倍体发掘[J]. 果树学报,2023,40(2):309-315.
XIE Shanpeng,YANG Wenhui,CHEN Hao,XIAO Gongao,XIE Kaidong,XIA Qiangming,WU Xiaomeng,GUO Wenwu. Production of nucellar seedlings and exploration of tetraploid from Satsuma mandarin Guoqing No.1[J]. Journal of Fruit Science,2023,40(2):309-315.
[12] 梁武军,解凯东,郭大勇,谢宗周,徐强,伊华林,郭文武. 柑橘10个品种实生后代多倍体的发掘及SSR鉴定[J]. 园艺学报,2014,41(3):409-416.
LIANG Wujun,XIE Kaidong,GUO Dayong,XIE Zongzhou,XU Qiang,YI Hualin,GUO Wenwu. Spontaneous generation and SSR characterization of polyploids from ten Citrus cultivars[J]. Acta Horticulturae Sinica,2014,41(3):409-416.
[13] 周锐,解凯东,王伟,彭珺,谢善鹏,胡益波,伍小萌,郭文武. 依据多倍体形态特征快速高效发掘柑橘四倍体[J]. 园艺学报,2020,47(12):2451-2458.
ZHOU Rui,XIE Kaidong,WANG Wei,PENG Jun,XIE Shanpeng,HU Yibo,WU Xiaomeng,GUO Wenwu. Efficient identification of tetraploid plants from seedling populations of apomictic Citrus genotypes based on morphological characteristics[J]. Acta Horticulturae Sinica,2020,47(12):2451-2458.
[14] 谢善鹏,解凯东,夏强明,周锐,张成磊,郑浩,伍小萌,郭文武. 柑橘6个地方品种资源四倍体高效发掘及分子鉴定[J]. 果树学报,2022,39(1):1-9.
XIE Shanpeng,XIE Kaidong,XIA Qiangming,ZHOU Rui,ZHANG Chenglei,ZHENG Hao,WU Xiaomeng,GUO Wenwu. Efficient exploration and SSR identification of 53 doubled diploid seedlings from six local Citrus cultivars and germplasm resources[J]. Journal of Fruit Science,2022,39(1):1-9.
[15] 张成磊,周锐,谢善鹏,解凯东,夏强明,伍小萌,郭文武. 山金柑实生后代四倍体发掘及形态和代谢评价[J]. 植物科学学报,2022,40(1):47-53.
ZHANG Chenglei,ZHOU Rui,XIE Shanpeng,XIE Kaidong,XIA Qiangming,WU Xiaomeng,GUO Wenwu. Exploration and evaluation of morphological traits and primary metabolites of tetraploid seedlings from Hongkong kumquat (Fortunella hindsii Swingle)[J]. Plant Science Journal,2022,40(1):47-53.
[16] 解凯东,王惠芹,王晓培,梁武军,謝宗周,伊华林,邓秀新,GROSSER J W,郭文武. 单胚性二倍体为母本与异源四倍体杂交大规模创制柑橘三倍体[J]. 中国农业科学,2013,46(21):4550-4557.
XIE Kaidong,WANG Huiqin,WANG Xiaopei,LIANG Wujun,XIE Zongzhou,YI Hualin,DENG Xiuxin,GROSSER J W,GUO Wenwu. Extensive Citrus triploid breeding by crossing monoembryonic diploid females with allotetraploid male parents[J]. Scientia Agricultura Sinica,2013,46(21):4550-4557.
[17] XIA Q M,MIAO L K,XIE K D,YIN Z P,WU X M,CHEN C L,GROSSER J W,GUO W W Localization and characterization of Citrus centromeres by combining half-tetrad analysis and CenH3-associated sequence profiling[J]. Plant Cell Reports,2020,39(12):1609-1622.
[18] CHENG Y J,GUO W W,YI H L,PANG X M,DENG X X. An efficient protocol for genomic DNA extraction from Citrus species[J]. Plant Molecular Biology Reporter,2003,21(2):177-178.
[19] CHEN C X,ZHOU P,CHOI Y A,HUANG S,JR GMITTER F G. Mining and characterizing microsatellites from citrus ESTs[J]. Theoretical and Applied Genetics,2006,112(7):1248-1257.
[20] CUENCA J,FROELICHER Y,ALEZA P,JUAREZ J,NAVARRO L,OLLITRAULT P. Multilocus half-tetrad analysis and centromere mapping in citrus:evidence of SDR mechanism for 2n megagametophyte production and partial chiasma interference in mandarin cv. ‘Fortune[J]. Heredity,2011,107(5):462-470.
[21] GARC?A-LOR A,LURO F,NAVARRO L,OLLITRAULT P. Comparative use of InDel and SSR markers in deciphering the interspecific structure of cultivated citrus genetic diversity:A perspective for genetic association studies[J]. Molecular Genetics and Genomics,2012,287(1):77-94.
[22] FROELICHER Y,DAMBIER D,BASSENE J B,COSTANTINO G,LOTFY S,DIDOUT C,BEAUMONT V,BROTTIER P,RISTERUCCI A M,LURO F,OLLITRAULT P. Characterization of microsatellite markers in mandarin orange (Citrus reticulata Blanco)[J]. Molecular Ecology Resources,2008,8(1):119-122.
[23] XU Q,CHEN L L,RUAN X A,CHEN D J,ZHU A D,CHEN C L,BERTRAND D,JIAO W B,HAO B H,LYON M P,CHEN J J,GAO S,XING F,LAN H,CHANG J W,GE X H,LEI Y,HU Q,MIAO Y,WANG L,XIAO S X,BISWAS M K,ZENG W F,GUO F,CAO H B,YANG X M,XU X W,CHENG Y J,XU J,LIU J H,LUO O J,TANG Z H,GUO W W,KUANG H H,ZHANG H Y,ROOSE M L,NAGARAJAN N,DENG X X,RUAN Y J. The draft genome of sweet orange (Citrus sinensis)[J]. Nature Genetics,2013,45(1):59-66.
[24] BARKLEY N A,ROOSE M L,KRUEGER R R,FEDERICI C T. Assessing genetic diversity and population structure in a citrus germplasm collection utilizing simple sequence repeat markers (SSRs)[J]. Theoretical and Applied Genetics,2006,112(8):1519-1531.
[25] KIJAS J M H,THOMAS M R,FOWLER J C S,ROOSE M L. Integration of trinucleotide microsatellites into a linkage map of Citrus[J]. Theoretical and Applied Genetics,1997,94(5):701-706.
[26] KOLTUNOW A M. Apomixis:Embryo sacs and embryos formed without meiosis or fertilization in ovules[J]. The Plant Cell,1993,5(10):1425-1437.
[27] 宋谢天,田啸宇,王楠,周银,谢源源,谢宗周,柴利军,叶俊丽,邓秀新. 利用InDel标记筛选多胚山金柑珠心苗后代[J]. 果树学报,2023,40(7):1312-1317.
SONG Xietian,TIAN Xiaoyu,WANG Nan,ZHOU Yin,XIE Yuanyuan,XIE Zongzhou,CHAI Lijun,YE Junli,DENG Xiuxin. InDel marker-assisted selection of nucellar seedlings in polyembryonic Fortunella hindsii[J]. Journal of Fruit Science,2023,40(7):1312-1317.
[28] 洪柳,刘永忠,邓秀新. 柑成熟种子胚培养获得四倍体植株[J]. 园艺学报,2005,32(4):688-690.
HONG Liu,LIU Yongzhong,DENG Xiuxin. Obtaining of ponkan (Citrus reticulata Blanco) tetraploid by culturing embryos of mature seeds[J]. Acta Horticulturae Sinica,2005,32(4):688-690.
[29] ALLARIO T,BRUMOS J,COLMENERO-FLORES J M,TADEO F,FROELICHER Y,TALON M,NAVARRO L,OLLITRAULT P,MORILLON R. Large changes in anatomy and physiology between diploid Rangpur lime (Citrus limonia) and its autotetraploid are not associated with large changes in leaf gene expression[J]. Journal of Experimental Botany,2011,62(8):2507-2519.
[30] ALEZA P,FROELICHER Y,SCHWARZ S,AGUST? M,HERN?NDEZ M,JU?REZ J,LURO F,MORILLON R,NAVARRO L,OLLITRAULT P. Tetraploidization events by chromosome doubling of nucellar cells are frequent in apomictic citrus and are dependent on genotype and environment[J]. Annals of Botany,2011,108(1):37-50.
[31] XIE K D,WANG X P,BISWAS M K,LIANG W J,XU Q,GROSSER J W,GUO W W. 2n megagametophyte formed via SDR contributes to tetraploidization in polyembryonic ‘Nadorcott tangor crossed by citrus allotetraploids[J]. Plant Cell Reports,2014,33(10):1641-1650.
[32] 梁武軍,解凯东,谢宗周,徐强,伊华林,郭文武. 三倍体葡萄柚实生后代多倍体的发掘与SSR遗传鉴定[J]. 果树学报,2015,32(1):13-18.
LIANG Wujun,XIE Kaidong,XIE Zongzhou,XU Qiang,YI Hualin,GUO Wenwu. Exploitation of polyploids from open-pollinated triploid grapefruit progenies and their genetic identification by SSR molecular markers[J]. Journal of Fruit Science,2015,32(1):13-18.
[33] GUO W W,DENG X X. Intertribal hexaploid somatic hybrid plants regeneration from electrofusion between diploids of Citrus sinensis and its sexually incompatible relative,Clausena lansium[J]. Theoretical and Applied Genetics,1999,98(3/4):581-585.
