氨基酸对荔枝愈伤组织增殖及体胚发生体系优化的研究
2023-11-21王果刘耀婷王家保李焕苓
王果 刘耀婷 王家保 李焕苓
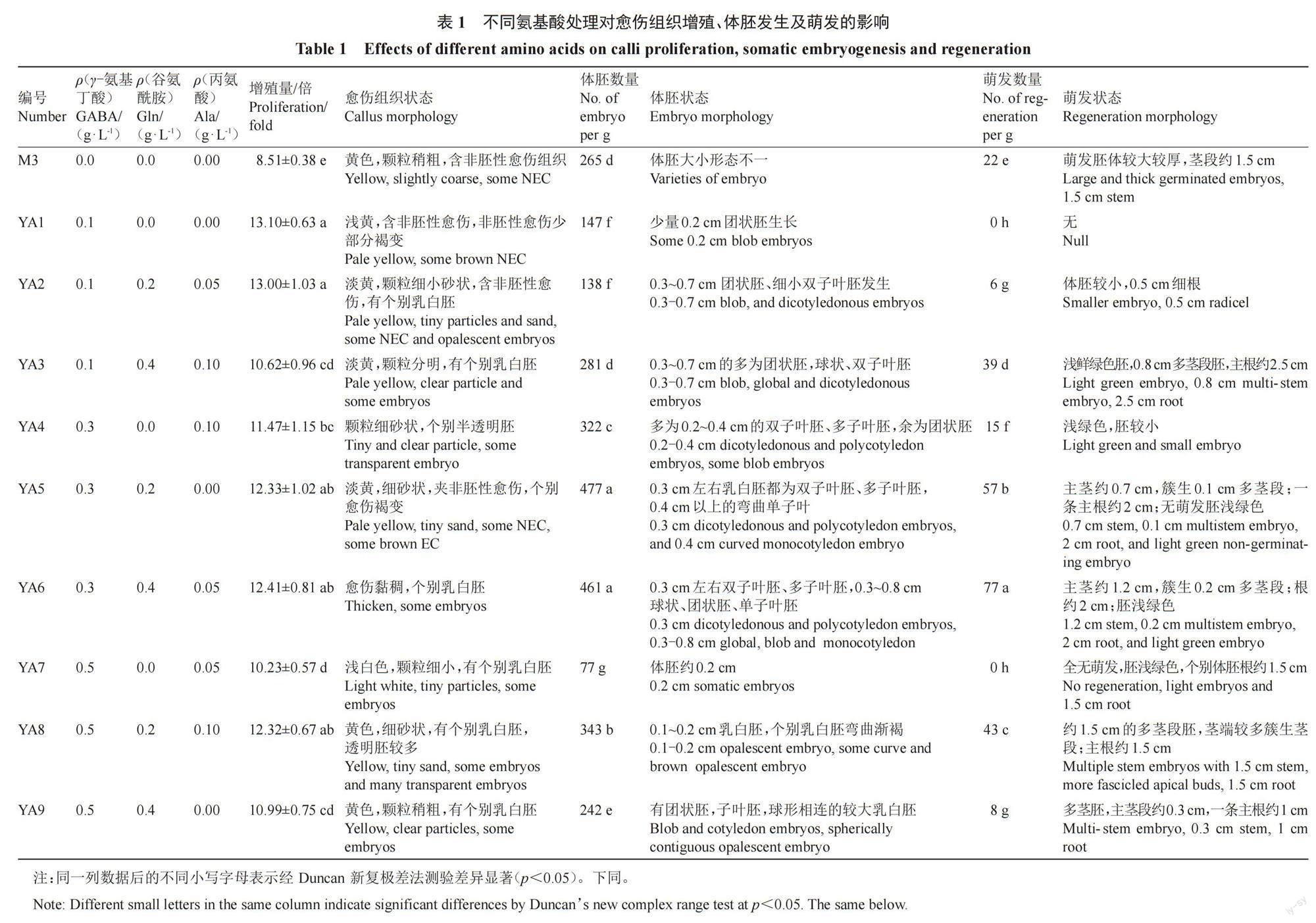

摘 要:【目的】探究氨基酸对荔枝愈伤组织增殖及体胚诱导的影响。【方法】采用L9(34)正交设计,分别在妃子笑荔枝愈伤组织继代及体胚诱导阶段添加不同氨基酸,比较其对愈伤组织增殖及体胚诱导的影响。【结果】在愈伤组织增殖阶段培养基中添加氨基酸均显著提高愈伤组织增殖率,最优处理为基础继代培养基添加0.1 g·L-1 γ-氨基丁酸+0.2 g·L-1谷氨酰胺,愈伤组织增殖率为初始接种量的14.04倍。与对照相比,氨基酸处理组的胚性愈伤组织淡黄,颗粒较小,细胞呈椭圆形,细胞质浓厚,多为正在分裂状态;非胚性愈伤组织浅白,水渍严重,着生于顶部或者边缘,易于剔除。在愈伤组织增殖阶段培养基中添加γ-氨基丁酸、谷氨酰胺和丙氨酸对后续的体胚发生及萌发影响极显著(p<0.01),最优配方为基础继代培养基添加0.3 g·L-1 γ-氨基丁酸 + 0.4 g·L-1谷氨酰胺+0.05 g·L-1丙氨酸,每克愈伤组织可获得461个体胚,77株组培苗。在体胚发生阶段培养基中添加氨基酸,获得的体胚发生及萌发最优配方为基础诱导培养基添加0.3 g·L-1 γ-氨基丁酸+0.4 g·L-1谷氨酰胺+0.05 g·L-1丙氨酸,每克愈伤组织可获得270个体胚,72株组培苗。【结论】氨基酸可调节愈伤组织胚性,提高愈伤组织增殖率,促进体胚发生及萌发。
关键词:荔枝;愈伤组织;体胚发生;体胚萌发;氨基酸
中图分类号:S667.1 文献标志码:A 文章编号:1009-9980(2023)11-2466-11
Study on the optimization of amino acids on proliferation of calli and somatic embryogenesis in Litchi chinensis ‘Feizixiao
WANG Guo1, LIU Yaoting1, 2, WANG Jiabao1*, LI Huanling 1
(1Environments and Plant Protection Institute, Chinese Academy of Tropical Agriculture Sciences/Danzhou Scientific Observing and Experimental Station of Agro-Environment, Ministry of Agriculture and Rural Affairs, P.R. China, Danzhou 571737, Hainan, China; 2College of Tropical Crops, Hainan University, Haikou 570228, Hainan, China)
Abstract: 【Objective】 Clonal propagation is an essential tool in biotechnology and breeding programs. Among the available clonal propagation methods, somatic embryogenesis is the most attractive technique for the large-scale propagation of plants and the long-term conservation of different embryogenic cell lines without changing their initial characteristics. Additionally, somatic embryogenesis is an ideal system for cell biology and molecular studies, including the study of the cell cycle, cell division, cell differentiation, and for understanding different genetic and epigenetic mechanisms for gene expression and phenotypic variation in plants. However, for many species, in vitro clonal propagation is currently difficult or inefficient. The low efficiency of somatic embryo induction is among the major problems hampering efficient regeneration and limiting practical applications. Therefore, high-quality somatic embryo production is one of the most important factors for using somatic embryogenesis protocols in commercial propagation and breeding programs. Litchi (Litchi chinensis Sonn.) is one of the treasure fruits cultivated in the south of China. Breeding elite litchi varieties with resistance to diseases is very important to the litchi industry. Many studies on the somatic embryogenesis of litchi have been reported. However, there has been no detailed report on the the effect of amino acids on callus proliferation and somatic embryo induction in litchi. The study aimed to investigate the effects of amino acids on the efficiency of somatic embryogenesis in Feizixiao litchi and apply these findings to improve the protocol for the induction of somatic embryos.【Methods】 The calli Feizixiao litchi were used as materials, the effects of different amino acids on callus proliferation, somatic embryogenesis and regeneration were studied using an L9(34) orthogonal experiment.【Results】 The results showed that the effects of glutamine (Gln) and γ-aminobutyric acid (GABA) on the callus morphology and proliferation rate were significant (p<0.01), while the effect of alanine (Ala) was not significant. The effects of the three amino acids on the proliferation rate of callus were in the order of Gln>GABA>Ala. The method of the callus proliferation was established. The optimal medium for the callus proliferation was: MS + 1 mg·L-1 2,4-D + 0.1 g·L-1 GABA + 0.05 g·L-1 Ala. After 20 days of culture, the proliferation rate of callus was 14.04 times as many as that of the control. Compared with the control group, the boundary between embryogenic callus and non-embryogenic calli in the amino acid treatment was clear and easy to distinguish.The embryogenic calli were granular and light yellow, while the non-embryonic calli were pale, heavily water stained, and was located at the top or edge and was easy to pick out. The callus cells treated with amino acids were elliptical with a dense cytoplasm and were mostly dividing. The callus cells on the control medium were divided into clumps. The effects of the Gln, GABA, and Ala on somatic embryogenesis and germination were significant (p<0.01). The effects of the three amino acids on the induction of somatic embryos and germination were in the order of Gln>GABA>Ala. The optimal treatment regimen was YA5: MS + 1 mg·L-1 2,4-D + 0.3 g·L-1 GABA + 0.2 g·L-1 Gln. The optimized program used for somatic embryo induction produced 477 embryos per gram of calli, and most of the somatic embryos were deformed curved monocotyledon embryos, while the smaller somatic embryos were dicotyledonous or polycotyledon embryos. The second medium for inducting somatic embryos was YA6 (MS + 1 mg·L-1 2,4-D + 0.3 g·L-1 GABA + 0.4 g·L-1 Gln + 0.05 g·L-1 Ala) with the induction number of 461 embryos per gram of calli, and the forms of the somatic embryos on YA6 were different, including globular embryos, clumped embryos, dicotyledonous embryos, and polycotyledon embryos. The best medium for inducing somatic embryos was the YA6 treatment, through which 77 regenerations were obtained per gram of callus, followed by the YA11 (MS + 1 mg·L-1 2, 4-D + 0.3 g·L-1 GABA + 0.4 g·L-1 Gln + 0.1 g·L-1 Ala) and YA5 treatments with 62 and 60 regenerations per gram of calli, respectively. The somatic embryo greening and germination in the amino acid treatments occurred later than that in the control. The regeneration appeared in the form of multiplying bud spots, the stems was bright green, the stem tips were bright white and bright green, and the somatic bodies were mostly light green with some red color. In the control, the somatic embryos were larger and thicker, and the stems were about 1.5 cm in length. The effects of the Gln, GABA, and Ala on the somatic embryogenesis and germination were significant (p<0.01). The effects of the three amino acids on the induction of somatic embryos were in the order of GABA>Gln>Ala. The effects of the three amino acids on the regeneration were in the order of Gln>GABA>Ala. The optimal treatment regimen (YAE6) for the somatic embryogenesis and germination was as follows: MS + 1 mg·L-1 2,4-D + 0.3 g·L-1 GABA + 0.4 g·L-1 Gln + 0.05 g·L-1 Ala with 270 embryos and 72 regenerations per gram of calli.【Conclusion】 The optimal amino acid treatment regimen for the callus proliferation was the same as that for the embryo induction, but the number of somatic embryos and germinations during the course of the callus proliferation were higher than that during the course of the somatic embryogenesis stage, indicating that the mechanism of the promotion of the somatic embryogenesis by amino acids might be related to the regulation of the embryogenic callus.
Key words:Litchi (Litchi chinensis Sonn.); Callus; Somatic embryogenesis; Somatic embryo germination; Amino acid
收稿日期:2023-05-11 接受日期:2023-08-21
基金项目:国家重点研发计划(2019YFD1000900);财政部和农业农村部:国家现代农业产业技术体系资助(CARS-32)
作者简介:王果,女,助理研究員,硕士,主要从事果树生物技术育种工作。E-mail:wanglucai@sina.com
*通信作者 Author for correspondence. E-mail:fdabo@163.com
荔枝(Litchi chinensis Sonn.)是原产于我国南方的大宗特色水果,营养丰富、味甜肉脆,素享“中华之珍品”美誉,深受消费者喜爱[1]。近年来,荔枝产业发展对优质新品种的需求不断上升。在组织培养技术的基础上,转基因育种具有育种周期短、可实现性状的快速定向改良等优点,已经成为荔枝新品种选育的主要途径[2-3]。但荔枝组织培养中普遍存在基因型依赖,以及体胚诱导效率低、萌发成苗难等问题[4-5],严重阻碍荔枝生物技术的育种进程。因此,急需优化荔枝体胚发生体系,提高体胚发生及萌发效率。
大量研究表明,氨基酸可作为信号或能源物质,参与植物的各种生理生化反应,调节植物的生长发育[6]。黄雪松等[7]在荔枝核中检出丙氨酸(alanine,Ala)的含量最高,崔珊珊[8]、彭颖等[9]等指出γ-氨基丁酸(γ-aminobutyric acid,GABA)和Ala是妃子笑等主栽荔枝品种果汁中的主要氨基酸。Pescador等[10]发现谷氨酰胺(glutamine,Gln)和天冬酰胺(asparagine,Asn)是雪莲合子胚形成过程中的主要氨基酸,Gln、GABA和谷氨酸(glutamicacid,Glu)在雪莲体胚发生时含量较高。孙小玲[11]从基因转录水平分析了44个与氨基酸代谢相关基因在小麦成熟胚脱分化过程中的差异表达,发现34个基因均上调表达,9个基因均下调表达,而1个基因以下调表达为主,表明氨基酸相关基因在细胞增殖、分化和组织结构功能重建阶段发挥作用。Rahmouni等[12]发现3.42 mmol·L-1的Gln、Asn或Ala等9种氨基酸可促进橡木次生胚形成与增殖,但胱氨酸(cystine,Cys)等10种氨基酸抑制次生胚形成。Raharjo等[13]在添加0.4 g·L-1 Gln的培养基上诱导荔枝营养梢幼叶获得胚性愈伤组织,Das等[5]在荔枝体胚萌发时添加0.45 g·L-1 Gln获得组培苗。关于氨基酸对荔枝离体再生的研究鲜见报道,笔者在本研究中以妃子笑荔枝愈伤组织为材料,在培养基中添加不同浓度的氨基酸,从愈伤组织结构及其增殖率、体胚发生及萌发数量等方面进行观察,探索氨基酸对荔枝愈伤组织增殖及体胚发生的影响,以期为揭示氨基酸在荔枝组织培养中的功能提供更多依据,也为建立荔枝高效体胚发生体系与生物技术育种研究开辟新的途径。
1 材料和方法
1.1 材料
供试材料为妃子笑荔枝愈伤组织,于2015年3月采用花药诱导获得,在基础继代培养基M3(MS[14]+ 1 mg·L-1 2, 4-D)和继代培养基M4上(MS + 1 mg·L-1 2, 4-D + 0.5 mg·L-1 KT + 5 mg·L-1 AgNO3)以30 d为1个周期交替继代保存[15]。
1.2 方法
1.2.1 在继代培养基中添加氨基酸对愈伤组织增殖、体胚发生及体胚萌发的影响 将胚性愈伤组织分别接种到添加不同氨基酸的继代培养基上。愈伤组织继代培养基采用3因素3水平正交试验设计,以M3为基本培养基(对照组),添加GABA(0.1、0.3、0.5 g·L-1)、Gln(0、0.2、0.4 g·L-1)、Ala(0、0.05、0.10 g·L-1)各3个水平,共9组,编号依次为YA1~YA9。培养20 d后,将愈伤组织分别接种于基础体胚诱导培养基T3(以MS为基本培养基,添加0.1 mg·L-1 NAA+5 mg·L-1 KT)上,培养7周后记录体胚发生情况,计算每克愈伤组织诱导获得的体胚数量。
上述获得的体胚在以MS为基本培养基、添加1 mg·L-1 ABA+0.5 mg·L-1 IAA的成熟培养基(编号为C19)上成熟8周,置于萌发培养基(1/2 MS+1 mg·L-1 GA3,编号R7)上萌发,8周后统计每克愈伤组织的体胚萌发数量及并进行形态观察等。
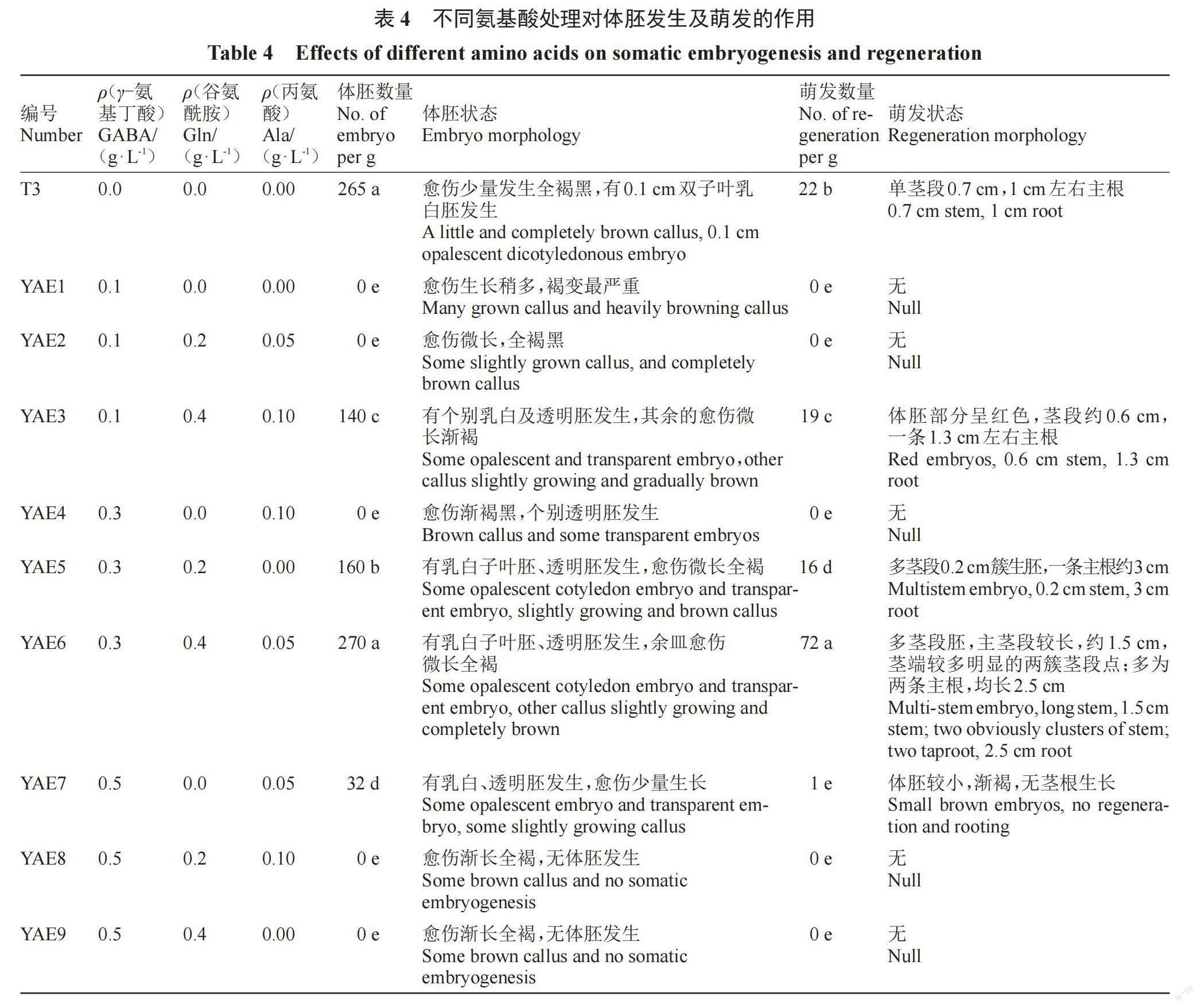
1.2.2 在体胚诱导培养基中添加不同氨基酸对体胚发生和萌发的影响 将在M3上继代增殖20 d的愈伤组织接种到添加不同氨基酸的体胚诱导培养基上,体胚诱导培养基采用3因素3水平正交试验设计,以T3为基本培养基(对照),添加GABA(0.1、0.3、0.5 g·L-1)、Gln(0、0.2、0.4 g·L-1)、Ala(0、0.05、0.10 g·L-1)各3个水平,共9组,编号依次为YAE1~YAE9。培养7周后记录体胚发生数量及形态。
体胚成熟与萌发同1.2.1。
1.2.3 愈伤组织增殖率及形态观察 上述培养了20 d后的愈伤组织,经清水法及甲醛-乙酸-乙醇(FAA)固定后采用苏木精染色法观察愈伤组织细胞形态,同时按下式计算增殖率[16]:
愈伤组织增殖率/倍=(20 d时愈伤组织量-愈伤组织初始接种量)/愈伤组织初始接种量。
1.2.4 数据统计分析 所有试验均重复2批次。每批次每处理各15皿,以每5皿为1个重复。采用DPS软件对数据进行单因素方差分析(One-way ANOVA)和Duncans多重比较。描述性统计值用2批6次重复的平均值表示。
2 结果与分析
2.1 在继代培养基中添加氨基酸对愈伤组织增殖的影响
由表1可知,氨基酸处理组的愈伤组织增殖率都显著高于对照。其中愈伤组织增殖率最高的是处理组YA1(MS + 1 mg·L-1 2, 4-D + 0.1 g·L-1 GABA),增殖量是初始接种量的13.10倍。
由图1和图2可知,添加氨基酸处理组的愈伤组织状态与对照存在明显差异。与对照相比,氨基酸处理组的胚性愈伤组织颗粒较细小,YA2的愈伤组织最细小,呈砂状;YA8和YA9有原胚分化(图1,绿框);胚性愈伤组织颜色淡黄,与非胚性愈伤组织界限明显,较易辨别;非胚性愈伤组织浅白,水渍化严重,且着生于顶部或者边缘,易于剔除(图1,红框)。氨基酸处理组的愈伤组织细胞呈椭圆形,细胞质浓厚,多为正在分裂状态(图2),而对照培养基的愈伤组织细胞已分裂成团。
2.2 在继代培养基中添加氨基酸对体胚诱导及萌发的影响
在氨基酸处理组增殖的愈伤组织诱导的体胚发生效率及状态存在明显差异(表1,图3),其中YA1、YA2、YA7和YA9的体胚发生数量显著低于对照,而YA4、YA5、YA6和YA8的体胚发生数量显著高于对照。体胚发生数量最高的是YA5(MS + 1 mg·L-1 2, 4-D + 0.3 g·L-1 GABA + 0.2 g·L-1 Gln),每克愈伤组织产生体胚数量高达477个。YA5上产生的较大体胚多为形态畸形的弯曲单子叶胚,较小的体胚为双子叶胚(图3,绿框)或多子叶胚(图3,红框)。处理组YA6(MS + 1 mg·L-1 2, 4-D + 0.3 g·L-1 GABA + 0.4 g·L-1 Gln + 0.05 g·L-1 Ala)上产生的体胚数量次之,每克愈伤组织产生461个体胚,体胚不同步,有球状胚、团状胚、双子叶及多子叶胚。
将上述所得体胚经成熟后转到萌发培养基上,发现氨基酸处理组的体胚呈现部分红色(图4-A,红框),在光下转绿及萌发的时间比对照晚约20 d,所得再生苗多为浅绿色。对照组上的体胚体积较大较厚,茎段长约1.5 cm。处理组YA1和YA7上的体胚全无萌发,仅长根。YA2、YA4和YA9上的体胚萌发数量较少,显著低于对照;YA3、YA5、YA6和YA8上的体胚萌发数量显著高于对照。体胚萌发数量最高的是YA6(MS + 1 mg·L-1 2, 4-D + 0.3 g·L-1 GABA + 0.4 g·L-1 Gln + 0.05 g·L-1 Ala),每克愈伤组织体胚萌发数量高达77株,萌发体胚多簇生0.2 cm茎段,主茎长平均为1.2 cm。
由愈傷组织增殖阶段正交试验结果的极差分析(表2)可知,GABA和Gln对愈伤组织的增殖作用影响较大,存在极显著差异(p<0.01),而Ala的影响不显著。水平优选愈伤组织增殖的组合为0.10 g·L-1 GABA + 0.20 g·L-1 Gln(编号YA10)。3种氨基酸对体胚发生及萌发作用存在极显著差异(p<0.01),3个影响因素的主次关系是:GABA>Gln>Ala。水平优选体胚发生及萌发的组合为0.30 g·L–1 GABA + 0.40 g·L-1 Gln + 0.10 g·L-1Ala(编号为YA11)。
2.3 优选的增殖培养基对愈伤组织增殖、体胚诱导及萌发的影响
优选的组合YA10(愈伤组织增殖率水平优选)、YA11(体胚发生及萌发水平优选)与最佳处理组YA1(愈伤组织增殖率最高)、YA5(体胚发生数量最多)和YA6(体胚萌发数最多)的结果(表3,图5)表明,氨基酸处理组的愈伤组织增殖率显著高于对照。其中YA10上的愈伤组织增殖率最高,其次依次为YA11、YA1、YA5和YA6。YA5和YA6上的体胚发生数量较高,每克愈伤组织可分别产生462、456个体胚,与对照及其他氨基酸处理组差异显著。YA6、YA11及YA5,体胚萌发数与对照差异显著,其中YA6上的体胚萌发数最高,每克愈伤组织可获得76株组培苗,其次依次是YA11、YA5,可分别获得62、60株组培苗。
2.4 在体胚诱导培养基中添加氨基酸对体胚诱导和萌发的影响
在体胚诱导阶段添加氨基酸,体胚发生效率存在明显差异(表4,图6),YAE6上的体胚发生数量高于对照;而其余处理组上产生的体胚数量都远低于对照,YAE1、YAE2、YAE4、YAE8和YAE9上全无乳白体胚发生,仅有个别透明体胚产生,愈伤组织渐长全褐,尤其是YAE1上的愈伤组织全褐黑,褐变最严重。YAE3、YAE5及YAE7同时有乳白体胚及透明体胚产生,愈伤组织少量生长。YAE6上体胚多为单、双及多子叶胚,体胚比对照大,大小在0.5~1.3 cm。
氨基酸处理组的体胚萌发效率与体胚诱导效率呈正比,YAE6每克愈伤组织可获得72株组培苗,显著高于对照及其他处理。组培苗多为簇生茎,主茎较高,平均1.5 cm,根系茂盛,主根长约2.5 cm。YAE3和YAE5体胚萌发数量均低于对照,差异显著。YAE7上仅有一个体积较小的体胚出现芽点,无根生长,且体胚渐褐。
由表5分析可知,3种氨基酸对体胚发生及萌发的作用差异显著(p<0.01),其中GABA和Gln的影响较大。水平优选体胚发生及萌发的组合为0.30 g·L-1 GABA + 0.40 g·L-1 Gln + 0.05 g·L-1 Ala,即处理组YAE6。
3 讨 论
较多研究表明,在植物组织培养中添加氨基酸可改善愈伤组织状态,促进体胚发生及萌发。Kong等[17]研究表明添加0.1 g·L-1 Gln、0.1 g·L-1 精氨酸(Arginine,Arg)和0.088 g·L-1 Asn的氨基酸混合物促进椰子愈伤组织形成,尤其是脯氨酸(proline,Pro)对脆性胚性愈伤组织的形成至关重要。Gleeson等[18]也指出Pro在胁迫条件下刺激了落叶松、云杉和橡树等胚性培养物的生长。Carlsson等[19]证明Gln促进云杉原胚性细胞团的生长。这与本研究结果一致,氨基酸促进荔枝胚性愈伤组织增殖,且添加氨基酸处理组的胚性愈伤组织与非胚性愈伤组织状态差异明显,更易于区分。
氨基酸在植物离体再生中的作用因品种而异,品种不同、氨基酸种类及浓度不同都极大影响着植物的再生效率。Kumar等[20]发现Gln明显促进了天竺葵体胚发生。Daniel等[21]发现丝氨酸(serine,Ser)、Pro或Gln都促进秋葵体胚成熟及萌发,氨基酸组合[0.3 g·L-1 Gln+0.4 g·L-1 酪蛋白水解物(casein hydrolysate,CH)]上成熟的体胚萌发数更高,获得的植株叶片更大更绿。Ashok等[22]比较了不同浓度的5种氨基酸(Gln、甘氨酸、Arg、Asn和Cys)对2个黄瓜品种花药体胚发生和植株再生的影响,发现氨基酸浓度过高或者太低都不能提高体胚产量,合适浓度的氨基酸混合液作用效果最佳。Pintos等[23]研究指出3种氨基酸(Gln、Arg和Asn)组合或单独添加GABA都可促进橡树体胚生长,体胚质量及大小都增加,但单独添加这3种氨基酸对体胚的诱导与对照组相比没有明显差异。Maruyama等[24]研究了不同浓度氨基酸混合液对日本雪松体胚成熟的影响,发现2倍的氨基酸混合液(2 g·L-1 Glu,1 g·L-1 天冬氨酸,0.5 g·L-1 Arg,0.158 g·L-1 瓜氨酸,0.152 g·L-1 鸟氨酸,0.11 g·L-1 赖氨酸,0.08 g·L-1 Ala)作用效果最佳,明显促进体胚成熟,成熟子叶胚效率提高27倍以上。Asthana等[25]比较了不同浓度Gln对三叶无患子愈伤组织的体胚发生效率和结节状胚性结构的诱导率,发现0.2 g·L-1 Gln作用效果最好,但在培养基中添加0.5 g·L-1 Gln或CH反而显著抑制了栎结节状胚性结构诱导率及次生胚发生效率的提高[26]。本研究中,笔者根据荔枝种子及果肉中氨基酸含量,首次分别在愈傷组织增殖及体胚发生过程中添加含3种主要氨基酸的不同混合液(0.30 g·L-1 GABA、0.4 g·L-1 Gln及0.05 g·L-1 Ala),发现2个阶段不同氨基酸组合均显著促进荔枝体胚发生及萌发。这些结果的不一致可能是植物种类、氨基酸种类及浓度、培养系统不同所导致的。
4 结 论
外源氨基酸促进荔枝愈伤组织增殖及体胚发生,GABA和Gln在愈伤组织增殖及胚性保持中起重要作用。在愈伤组织增殖或体胚诱导阶段添加0.30 g·L-1 GABA + 0.40 g·L-1 Gln + 0.05 g·L-1 Ala,2个阶段每克愈伤组织获得的体胚及组培苗数量分别为461个、77株,270个、72株。在愈伤组织增殖过程中添加氨基酸的体胚发生效果优于在体胚诱导阶段添加,说明氨基酸促进体胚发生的机制可能与胚性愈伤组织的调控关系更大。
参考文献 References:
[1] HU G B,FENG J T,XIANG X,WANG J B,SALOJ?RVI J,LIU C M,WU Z X,ZHANG J S,LIANG X M,JIANG Z D,LIU W,OU L X,LI J W,FAN G Y,MAI Y X,CHEN C J,ZHANG X T,ZHENG J K,ZHANG Y Q,PENG H X,YAO L X,WAI C M,LUO X P,FU J X,TANG H B,LAN T Y,LAI B,SUN J H,WEI Y Z,LI H L,CHEN J Z,HUANG X M,YAN Q,LIU X,MCHALE L K,ROLLING W,GUYOT R,SANKOFF D,ZHENG C F,ALBERT V A,MING R,CHEN H B,XIA R,LI J G. Two divergent haplotypes from a highly heterozygous lychee genome suggest independent domestication events for early and late-maturing cultivars[J]. Nature Genetics,2022,54(1):73-83.
[2] DAS D K,RAHMAN A. Expression of a bacterial chitinase (ChiB) gene enhances antifungal potential in transgenic Litchi chinensis Sonn. (cv. Bedana)[J]. Current Trends in Biotechnology and Pharmacy,2010,4(3):820-833.
[3] WANG S J,WANG G,LI H L,LI F,WANG J B. Agrobacterium tumefaciens-mediated transformation of embryogenic callus and CRISPR/Cas9-mediated genome editing in ‘Feizixiao litchi[J]. Horticultural Plant Journal,2023,9(5):947-957.
[4] WANG G,LI H L,WANG S J,SUN J H,ZHANG X C,WANG J B. In vitro regeneration of Feizixiao litchi (Litchi chinensis Sonn.)[J]. African Journal of Biotechnology,2016,15(22):1026-1034.
[5] DAS D K,RAHMAN A,KUMARI D,KUMARI N. Synthetic seed preparation,germination and plantlet regeneration of Litchi (Litchi chinensis Sonn.)[J]. American Journal of Plant Sciences,2016,7(10):1395-1406.
[6] GALILI G,AVIN-WITTENBERG T,ANGELOVICI R,FERNIE A R. The role of photosynthesis and amino acid metabolism in the energy status during seed development[J]. Frontiers in Plant Science,2014,5:447.
[7] 黄雪松,陈杰. 测定荔枝核中的游离氨基酸[J]. 氨基酸和生物资源,2007,29(2):11-14.
HUANG Xuesong,CHEN Jie. Determination of free amino acids in Litchi stone[J]. Amino Acids & Biotic Resources,2007,29(2):11-14.
[8] 崔珊珊,胡卓炎,余恺,李雅萍,林文祥,余小林. 不同产地妃子笑荔枝果汁的氨基酸组分[J]. 食品科学,2011,32(12):269-273.
CUI Shanshan,HU Zhuoyan,YU Kai,LI Yaping,LIN Wenxiang,YU Xiaolin. Amino acid composition of Feizixiao Litchi juice from different geographic origins[J]. Food Science,2011,32(12):269-273.
[9] 彭颖,周如金. 不同品种荔枝果汁氨基酸和糖类的测定与分析[J]. 中国食品添加剂,2017(4):173-177.
PENG Ying,ZHOU Rujin. Determination and analysis of amino acids and sugars in different types of Litchi juice[J]. China Food Additives,2017(4):173-177.
[10] PESCADOR R,KERBAUY G B,ENDRES L,DE FREITAS FRAGA H P,GUERRA M P. Synthesis and accumulation of free amino acids during somatic and zygotic embryogenesis of Acca sellowiana (O. Berg.) Burret[J]. African Journal of Biotechnology,2013,12:3455-3465.
[11] 孫小玲. 氨基酸代谢相关基因在小麦成熟胚脱分化过程中的表达研究[D]. 郑州:河南农业大学,2009.
SUN Xiaoling. Expression analysis of genes related to the metabolism of amino acids during dedifferentiating mature wheat embryos[D]. Zhengzhou:Henan Agricultural University,2009.
[12] RAHMOUNI S,EL ANSARI Z N,BADOC A,MARTIN P,EL KBIACH M L,LAMARTI A. Effect of amino acids on secondary somatic embryogenesis of Moroccan cork oak (Quercus suber L.) tree[J]. American Journal of Plant Sciences,2020,11(5):626-641.
[13] RAHARJO S H T,LITZ R E. Somatic embryogenesis and plant regeneration of Litchi (Litchi chinensis Sonn.) from leaves of mature phase trees[J]. Plant Cell,Tissue and Organ Culture,2007,89(2):113-119.
[14] MURASHIGE T,SKOOG F. A revised medium for rapid growth and bio assays with tobacco tissue cultures[J]. Physiologia Plantarum,1962,15(3):473-497.
[15] 赖钟雄. 龙眼生物技术研究[M]. 福州:福建科学技术出版社,2003:67-70.
LAI Zhongxiong. Research of the longan biotechnology[M]. Fuzhou:Fujian Science & Technology Publishing House,2003:67-70.
[16] 王果,刘耀婷,李焕苓,李芳,王树军,王家保. 外源多胺对荔枝愈伤组织增殖及体胚发生的作用[J]. 果树学报,2021,38(12):2135-2147.
WANG Guo,LIU Yaoting,LI Huanling,LI Fang,WANG Shujun,WANG Jiabao. Effects of exogenous polyamine application on callus proliferation and somatic embryogenesis in Litchi chinensis ‘Feizixiao[J]. Journal of Fruit Science,2021,38(12):2135-2147.
[17] KONG E Y Y,BIDDLE J,KALAIPANDIAN S,ADKINS S W. Coconut callus initiation for cell suspension culture[J]. Plants,2023,12(4):968.
[18] GLEESON D,LELU-WALTER M A,PARKINSON M. Influence of exogenous L-proline on embryogenic cultures of larch (Larix leptoeuropaea Dengler),sitka spruce [Picea sitchensis (Bong.) Carr.] and oak (Quercus robur L.) subjected to cold and salt stress[J]. Annals of Forest Science,2004,61(2):125-128.
[19] CARLSSON J,SVENNERSTAM H,MORITZ T,EGERTSDOTTER U,GANETEG U. Correction:Nitrogen uptake and assimilation in proliferating embryogenic cultures of Norway spruce-investigating the specific role of glutamine[J]. PLoS One,2018,13(1):e0191208.
[20] KUMAR V,MOYO M,VAN STADEN J. Somatic embryogenesis of Pelargonium sidoides DC.[J]. Plant Cell,Tissue and Organ Culture,2015,121(3):571-577.
[21] DANIEL M A,DAVID R H A,CAESAR S A,RAMAKRISHNAN M,DURAIPANDIYAN V,IGNACIMUTHU S,AL-DHABI N A. Effect of l-glutamine and casein hydrolysate in the development of somatic embryos from cotyledonary leaf explants in okra (Abelmoschus esculentus L. Monech)[J]. South African Journal of Botany,2018,114:223-231.
[22] ASHOK K H G,MURTHY H N. Effect of sugars and amino acids on androgenesis of Cucumis sativus[J]. Plant Cell,Tissue and Organ Culture,2004,78(3):201-208.
[23] PINTOS B,MANZANERA J A,BUENO M A. Oak somatic and gametic embryos maturation is affected by charcoal and specific aminoacids mixture[J]. Annals of Forest Science,2010,67(2):205.
[24] MARUYAMA T E,UENO S,MORI H,KANEEDA T,MORIGUCHI Y. Factors influencing somatic embryo maturation in Sugi [Japanese Cedar,Cryptomeria japonica (Thunb. ex L. f.) D. Don][J]. Plants,2021,10(5):874.
[25] ASTHANA P,RAI M K,JAISWAL U. Somatic embryogenesis from sepal explants in Sapindus trifoliatus,a plant valuable in Herbal soap industry[J]. Industrial Crops and Products,2017,100:228-235.
[26] MART?NEZ M T,SAN JOS? M C,VIEITEZ A M,CERNADAS M J,BALLESTER A,CORREDOIRA E. Propagation of mature Quercus ilex L. (Holm Oak) trees by somatic embryogenesis[J]. Plant cell,Tissue and Organ Culture,2017,131:321-333.
