西瓜连作根际土壤微生物群落演替特征
2023-11-18郭晗玥王东升阮杨乔亦铸张芸滔李玲黄启为郭世伟凌宁沈其荣
郭晗玥,王东升,阮杨,乔亦铸,张芸滔,李玲,黄启为,郭世伟,凌宁,沈其荣
西瓜连作根际土壤微生物群落演替特征

1南京农业大学资源与环境科学学院/国家有机肥重点实验室/江苏省固体有机废物利用重点实验室,南京 210095;2南京市蔬菜科学研究所,南京210042
【目的】研究连作对西瓜根际土壤中细菌、真菌群落构建和潜在功能的影响,明确根际微生物对环境变化的适应性,为西瓜连作障碍的生态防治和农田土壤的健康保持提供理论基础。【方法】以未连作(对照)、连作2茬和连作 6茬的西瓜根际土壤为研究对象,利用16S rRNA和ITS高通量测序技术解析西瓜连作对根际土壤细菌、真菌群落特征和演替的影响。【结果】随着连作茬次的增加,西瓜根际土壤中细菌多样性指数呈现先下降后上升趋势,真菌多样性指数显著下降。在细菌属水平,多茬次的连作降低了西瓜根际土壤中鞘氨醇单胞菌属()和溶杆菌属()的相对丰度;在真菌属水平,镰刀菌属()相对丰度随连作茬次增加呈增加趋势。与对照相比,连作的网络复杂度更高,但网络结构稳定性低。此外,相比于对照,连作6茬显著减少有害物质的生物降解途径以及氨基酸的代谢途径的相对丰度,并且在连作6茬后真菌群落中病原真菌相对丰度显著增多。在群落演替过程中,随机性过程主导连作下西瓜根际细菌群落构建,而确定性过程主导连作下西瓜根际真菌群落构建。【结论】连作引起根际细菌和真菌的群落特征、功能组成和演替过程变化,细菌群落关键功能的减少、病原真菌相对丰度的增加以及微生物群落网络稳定性的减小可能是导致西瓜连作障碍发生的重要因素。
西瓜;连作土壤;根际微生物群落;功能预测;群落演替
0 引言
【研究意义】西瓜(Thunb. Matsum and Naka)是我国乃至世界主要的夏季水果之一[1],其具有较高的经济价值以及巨大的市场需求,促使农户利用现代农业技术进行集约化生产。然而西瓜连年单作现象十分普遍[2],导致西瓜连作障碍愈发严重且成为了限制西瓜产业持续稳定发展的重要因素[3]。长期连作也会引起土壤理化性质改变、土壤病原菌富集以及自毒物质的积累,最终导致作物生长受限[4-5]。土壤微生物在保持土壤功能和根际生态系统可持续性方面起着至关重要的作用,且通常被认为是土壤健康的敏感生物指标[6-7]。研究连作对西瓜根际土壤中细菌、真菌群落结构和多样性的影响,对防治西瓜连作障碍和保持农田土壤的健康具有重要意义。【前人研究进展】有研究表明,对连作土壤进行灭菌能够显著提高植物的生长和品质[8],这表明生物因素是植物连作土壤病害发生的主要原因。植物根际拥有大量微生物,它们参与土壤养分循环,影响植物对养分的吸收与利用,对于植物生长起着至关重要的作用[9-10]。植物根际同时也是土传病原菌进入植物的必经之地,在入侵植物体之前,病原菌会先与土著微生物竞争生态位,再从根际感染植物体[5,11]。过往研究发现,长期的单一作物栽培所引起的土壤性质恶化以及植物根系分泌物在内的化感作用会改变根际微生物群落特征[12-14]。有研究表明,连作会简化土壤微生物群落生态网络并降低网络稳定性,这可能与土壤理化性质恶化、土壤肥力下降密切相关[15]。此外,了解微生物群落组装的生态过程有助于识别微生物群落组成如何响应环境变化,其中确定性过程和随机性过程基于生态位理论和中性理论,通常用于解释微生物群落构建[16]。确定性过程包括非生物(pH、盐度、有机碳含量[17-18])和生物(捕食、共生和寄生等[19])因素,决定物种的存在与否和相对丰度,与生态选择相关;随机过程包括不可预测的扰动、概率性的散布和随机的出生-死亡事件等,这些变化不是由环境决定的适应性结果。确定性过程和随机过程在构建生态群落中同时起作用来调节群落构建[20-21]。【本研究切入点】目前对西瓜长期连作体系下根际土壤微生物群落特征及其构建过程研究较少,这对于预测根际微生物在调节植物-微生物生态系统功能中的作用是至关重要的。【拟解决的关键问题】本研究以不同连作茬次的西瓜为研究对象,采用16S rRNA和ITS高通量测序技术,研究连作对西瓜根际土壤细菌和真菌群落组成的影响,同时结合PICRUSt2和FUNGuild分别分析连作土壤细菌群落功能和真菌营养型变化,并探究连作对根际微生物分子生态网络复杂性和稳定性的影响,最后通过计算Modified Stochasticity Ratio(校正的随机率)来解析根际微生物群落构建机制。总之,研究有助于揭示西瓜连作的障碍机制,从而为西瓜的种植和产业可持续发展提供理论支持。
1 材料与方法
1.1 研究地点概况
在南京市蔬菜花卉科学研究所(31°43′N,118°47′E)建立了一个长期试验大棚。试验地土壤为黄棕壤,年平均日照时数2 016.4 h,年均气温14.7℃。
1.2 试验设计和土壤样品采集
本试验在同一地块种植品种为8424西瓜,种植面积为28 m2,共种植32株,种植间距为0.45 m,其生长期间日均温度38℃,夜均温度24℃。自2014年,每年进行春(3月)、秋(8月)两茬种植。本试验共设置3个连作时间点进行采样,分别于2014(西瓜未连作,对照组CK)、2015(连作2茬,CC2)、2017年(连作6茬,CC6)的5月采集西瓜根际土壤样品。在每一个时间点随机采取9株长势相似西瓜的根际土,其中每3株合并为一个混合样本,即每个时间点有3个重复样本。共获得9个样本(3个时间点×3次重复)。对于根际土壤的采集,将根部完整取出,并轻微摇动去除松散的可见土壤[22]。将根样品切成约2 cm小段,装入50 mL无菌无酶离心管中,加入适量无菌蒸馏水,置于摇床中以180 r/min转速振荡30 min,洗涤紧贴根部表面的土壤,之后将离心管中的根取出,并以8 000 r/min离心10 min得到根际土壤样品[23],将收集的根际土壤保存在-80℃冰箱中进行后续分析。
1.3 DNA提取和Illumina MiSeq测序
使用FastDNA™(MP Biomedicals, Cleveland, OH, USA)土壤DNA提取试剂盒从0.5 g湿重土壤中提取根际土壤微生物总DNA。之后使用Qubit®DNA浓度检测试剂盒测定提取的DNA质量。分别使用引物515F/907R和ITS1F/ITS2R引物扩增细菌基因V4—V5区和真菌基因ITS1区。使用Illumina Miseq系统进行测序。
1.4 数据统计与分析
使用MOTHUR软件对下机数据进行质量控制[24]。具体而言,将短序列以及低质量序列进行去除,通过数据库比对,去除存在的嵌合体序列。将剩余的高质量序列以97%的阈值生成可操作分类单元(operational taxonomic units,OTUs)并选取代表序列进行分类学分析,再基于样本最小序列数对所有样本抽平后计算chao1、richness和shannon多样性指数。基于 Bray-Curtis 距离用“R(v4.0.2)”中的“vegan”包进行主坐标分析(PCoA)。使用“pheatmap”和“ggplot2”包在R环境中揭示不同土壤样品中细菌和真菌物种在门水平和属水平上的差异。采用PICRUSt2软件对细菌OTU丰度表进行标准化后进行功能基因预测分析,将OTU信息与KEGG(Kyoto Encyclo-pediaof Genesand Genomes)数据库进行比对,获得三级功能基因的丰度,参考KEGG的二级代谢通路的功能基因类别划分归类,获得功能基因组成。为了确定真菌营养模式分类群,使用FUNGuild进行注释[25],并保留probable和highly probable置信度的数据,以及single mode和mixed mode的数据。
使用随机矩阵理论(RMT)自动确定合适的网络阈值[26],基于所有处理的细菌和真菌总OTU丰度矩阵进行物种共发生网络分析并使用Gephi0.9.2得到细菌总网络和真菌总网络。之后从总节点和边文件中抽提出每个处理所拥有的节点和边,得到每个处理的细菌子网络和真菌子网络。通过计算网络各节点的模块内连通性()和模块间连通性(),识别网络中潜在关键物种[27]。通过计算和值的大小,将网络关键物种划分为三类:模块中心(模块内高度连接的节点,>2.5和<0.62)、网络中心(整个网络内高度连接的节点,>2.5和>0.62)和网络连接节点(连接不同模块的节点,<2.5和>0.62)[26]。使用Gephi0.9.2绘制网络图。使用自然连通度指标确定网络结构的稳定性[28],通过去除网络中的节点来模拟受到的外界扰动,并计算节点去除后网络的稳定性,以评估网络在受到随机攻击后稳定性下降的速度。最后,使用校正的随机率(modified stochasticity ratio,MST)来量化确定性和随机过程在群落组装过程中的相对重要性,以50%作为更确定性(MST<50%)和更随机(MST>50%)集合之间的分界点[16]。
2 结果
2.1 连作对西瓜根际细菌和真菌群落多样性和组成的影响
不同连作茬次下西瓜根际细菌群落的chao1、richness和shannon指数随着连作茬次的延长呈现先下降后上升的趋势(图1-A),而真菌群落的相应指数显著降低(图1-B)。这表明,连作在一定程度上有利于细菌群落丰富度的增加,而降低了真菌群落丰富度。基于Bray-Curtis距离的主坐标分析(PCoA)对对照、连作2茬、连作6茬的根际细菌和真菌群落组成特征进行可视化,结果表明,不同连作茬次中根际细菌和真菌群落聚集成不同的簇,也说明了随着连作茬次的增加,微生物群落组成发生显著改变(图2-A,Adonis2=0.71,=0.003;图2-B,Adonis2=0.84,=0.004)。
我们将平均相对丰度大于1%的细菌门和真菌门划分为优势菌门,3种不同连作茬次的根际土壤中细菌优势门分别是变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)(图2-C)。在细菌群落中,变形菌门在3种不同连作茬次中土壤的比例均最高,占比为83.3%—47.1%,并且随着连作茬次增加呈先减少后增加趋势。相比于对照,拟杆菌门和厚壁菌门的相对丰度在连作2茬中增加了22.2%和11.2%,而在连作6茬中分别增加了5.1%和0.76%(图2-C)。属于优势真菌门的是子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、壶菌门(Chytridiomycota)(图2-D)。真菌群落中,子囊菌门和担子菌门不仅相对丰度最高,且随连作茬次表现出不同的差异变化。在真菌群落中,子囊菌门在3个不同连作茬次土壤中的占比均最高,为32.16%—66.1%,并且其相对丰度随连作茬次增加呈先减少后增加趋势。而担子菌门的相对丰度呈先增加后减少趋势(图2-D)。在属水平,相比于对照,连作2茬和6茬细菌群落中鞘氨醇单胞菌属()相对丰度分别减少了4.5%和0.6%,鞘氨醇菌属()相对丰度分别减少了7.6%和7.2%,溶杆菌属()相对丰度分别减少了2.5%和1.6%;连作6茬与对照相比,剑菌属()和芽孢杆菌属()相对丰度分别减少了1.1%和0.2%(图2-E)。真菌群落中,相比于对照,连作2茬和6茬的镰刀菌属()相对丰度分别增加了4.1%和60.6%(图2-F)。这些结果表明连作过程明显改变了根际细菌、真菌群落结构,且群落组成相似度随连作茬次的增加而降低。
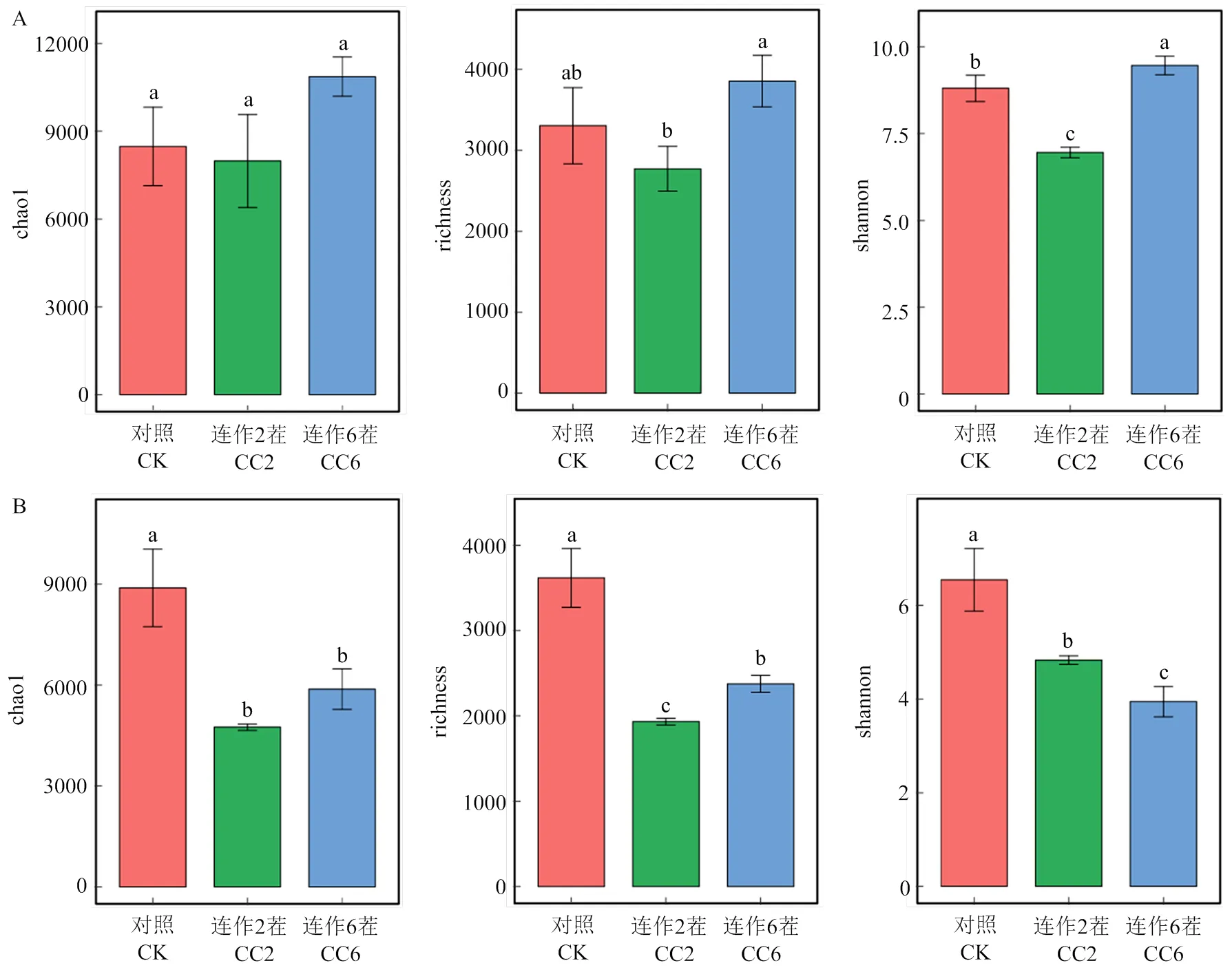
A:不同连作茬次下根际细菌群落chao1、richness和shannon指数;B:不同连作茬次下根际真菌群落chao1、richness和shannon指数。不同的字母表示不同处理间细菌、真菌丰度的显著差异(P<0.05)
2.2 连作对西瓜根际细菌和真菌群落预测功能的影响
采用PICRUSt2对各个连作茬次中根际细菌群落进行功能预测。在三级功能水平的预测功能中,对照的西瓜根际细菌群落共得到297个代谢通路,相较于其他两个时期,富集了50个代谢通路;连作2茬共得到295个代谢通路,相较于其他两个时期,富集了41个代谢通路;连作6茬共得到298个代谢通路,相较于其他两个时期,富集了26个代谢通路(图3-a)。将所获得相对丰度高的代谢通路进行二级功能水平归类,结果显示,除其他次生代谢物的生物合成外,其余功能基因的相对丰度随连作茬次变化均有不同的响应(图3-c)。相比于对照,连作2茬会显著减少碳水化合物、萜类和多酮类化合物的代谢、细胞生长和死亡等代谢途径相对丰度,而增加了复制和修复、辅助因子和维生素的代谢、运输和分解代谢等途径;连作6茬会显著减少有害物质的生物降解和代谢及其他氨基酸的代谢等途径的相对丰度,反而增加了多糖的生物合成和代谢途径(图3-c)。真菌功能群使用FUNGuild进行预测,其中连作6茬会显著富集病理型(pathotroph)真菌(图3-b)。其中相比于对照,连作6茬会显著富Plant pathogen(植物病原体)类型真菌,而Soil saprotroph(土壤腐生生物)会显著减少(图3-d)。

A:不同连作茬次下根际细菌群落主坐标分析(PCoA);B:不同连作茬次下根际真菌群落主坐标分析(PCoA);C:不同连作茬次下根际细菌群落门水平组成;D:不同连作茬次下根际真菌群落门水平组成;E:不同连作茬次下根际细菌群落属水平组成;F:不同连作茬次下根际真菌群落属水平组成

a:不同连作茬次中KEGG三级代谢通路相对丰度差异变化。括号中的数字分别代表着比其他两个处理显著富集的代谢通路数量和预测得到的代谢通路总数;b:不同连作茬次中根际真菌营养型相对丰度的差异分析,括号中的数字代表着比其他两个处理显著富集的真菌营养型数量;c:饼图表示将图a中富集的三级代谢通路归类于二级代谢通路;d:不同连作茬次中根际真菌营养亚型相对丰度的差异分析
2.3 连作对西瓜根际细菌和真菌群落共生网络结构和关键物种的影响
基于OTU丰度矩阵进行物种共发生网络分析并得到细菌总网络和子网络以及真菌总网络和子网络(图4-a)(见方法2.4)。结果显示,连作2茬与连作6茬的根际细菌和真菌网络比对照具有更多的网络节点、平均度和种间相互作用,而模块化指数随着连作茬次增加而下降(表1)。变形菌门在3个连作茬次(对照组、连作2茬和连作6茬,分别占62.6%、63.4%和65.9%)的网络互作关系中占主导地位,与其他物种相互联系的数量最多(图4-a)。这表明与对照相比,连作增加了网络中变形菌门物种的比例。在真菌群落网络中,子囊菌门在3个连作茬次中(对照组、连作2茬和连作6茬,分别占73.2%、70.7%和69.6%)的网络互作关系中占主导地位,与其他物种相互联系的数量最多(图4-d)。这表明与对照相比,连作减少了网络中子囊菌门物种的比例。此外,对照、连作2茬与连作6茬的真菌共生网络中正相关连接数占比分别为41.33%、51.48%和55.30%(表1),表明在连作过程中根际真菌间趋于竞争关系而不是合作关系。

表1 不同连作茬次下西瓜根际细菌和真菌网络拓扑结构
网络结构稳定性的结果显示,在细菌群落中,虽然连作6茬的根际初始网络自然连通性最高(0.0078),其次是连作2茬(0.0075)和对照网络(0.0068),但是当去除20%的网络节点时,连作6茬的自然连通性急剧降至0.0028;当去除40%的网络节点时,连作2茬的自然连通性急剧降至0.0024(图4-b)。随着节点不断被移除,对照网络的连通度下降趋势(Slope=-0.004)比连作2茬(Slope=-0.005)和连作6茬(Slope=-0.006)更加平稳(图4-b)。而真菌群落中,连作6茬的根际初始网络自然连通性仍是最高(0.085),其次是对照组(0.023)和连作2茬(0.013)网络,但是当去除40%的网络节点时,连作6茬的自然连通性急剧降至0.0071,而连作2茬网络随着节点不断被移除,其自然连通性下降趋势(Slope=-0.011)比连作2茬(Slope=-0.021)和连作6茬(Slope=-0.077)更加平稳(图4-e);对照网络在去除20%的网络节点后,其自然连通性稳定在一定水平(图4-e)。这些结果表明在移除网络节点后,对照的根际微生物网络结构仍保持相对较高的稳定性。
对照的细菌群落共拥有9个关键物种(图4-c),网络模块中心(module hubs)主要来源于变形菌门和厚壁菌门,其中主要来自于假单胞菌属()、芽孢杆菌属()和鞘氨醇单胞菌属()等。对照的真菌群落共拥有1个关键物种(图4-f),网络模块中心(module hubs)主要来源于子囊菌门的闭小囊菌属()。
2.4 连作对西瓜根际细菌和真菌群落构建过程的影响
使用校正的随机率(MST)定量生态过程中的随机性和确定性。结果表明,在细菌群落中,随机性过程在对照和连作6茬的细菌群落内占主导,而在连作2茬的细菌群落中确定性过程占主导(图5-A);在真菌群落中,确定性过程在连作2茬和连作6茬中占主导(图5-B)。

A:不同连作茬次下根际细菌群落的MST柱状图;B:不同连作茬次下根际真菌群落的MST柱状图。不同的字母表示不同处理间的显著差异(P<0.05)
3 讨论
3.1 西瓜连作对根际土壤细菌和真菌群落组成和功能的影响
在本研究中,利用Illumina Miseq高通量技术分析了连作后西瓜根际土壤细菌和真菌群落结构。我们发现随着连作茬次的增加,细菌和真菌群落特征差异较大。这可能是由于在连作系统中,相同的田间管理模式、植物化感物质的长期累积以及同类型根系分泌物长期释放的生态效应,会使微生物区系发生显著改变[13,29]。具体来说,相比于对照,西瓜根际细菌群落的richness和shannon指数在连作2茬和6茬中表现出先下降后上升的趋势,而chao1指数没有显著的变化,这与部分前人的研究结果一致[30-31]。多样性指数出现下降的原因可能是在2茬的连作过程中肥料的添加会暂时的增加营养物质,土壤中的部分物种对营养物质的添加反应迅速,进而通过快速繁殖占领寡营养微生物的生态位,导致微生物多样性立即下降[32]。而连作6茬过程中,随着养分长期大量的积累,包括肥料的连续性投入和植物分泌物和残体遗留,土壤中生态位会在一定程度上扩张,会有更多的有机物质和养分易被微生物利用,从而促进了细菌的生长,有利于根际土壤中细菌多样性的提高。有研究表明,变形菌门和酸杆菌门在养分循环中起重要作用,并且也与抑制病害等功能相关[33-35]。其中鞘氨醇单胞菌属(变形菌门)具有降解多环芳烃和降低纤维素的能力[36],溶杆菌属(变形菌门)对真菌、革兰氏阴性细菌、革兰氏阳性细菌和线虫都有突出的拮抗作用[37]。而本研究结果中,相对于对照,连作后的变形菌门和酸杆菌门的相对丰度都呈现降低的趋势,而且鞘氨醇单胞菌属和芽孢杆菌属等的相对丰度降低,从而导致植物抵御病原菌感染的能力下降,并且这种降低也会增加发生土壤病害的风险。而连作后的真菌群落相比于对照,chao1、richness和shannon指数显著降低,这与吴凤芝等的研究结果相一致[38]。有研究指出,微生物多样性的减少是发生土壤病害的重要原因[5],而在本研究中真菌群落的多样性指数在连作后显著降低,可能增加发生土壤病害的风险。在真菌群落中,有研究表明镰刀菌属(子囊菌门)为致病菌,会引起西瓜、黄瓜枯萎病的发生[39-40],而本研究结果显示出镰刀菌属的相对丰度在连作6茬后显著增加。在CHEN等[41]的研究中得出,真菌病原体的增加和有益真菌的减少可能是导致连作植物生长和产量下降的重要因素。因此,连作会改变微生物群落多样性和组成,破坏根际生态平衡,从而更加容易引起连作障碍。
我们利用PICRUSt2对不同连作茬次下根际细菌群落进行功能预测,发现根际细菌群落积极参与了基本的代谢过程。其中碳水化合物代谢途径与固氮、溶磷等作用有着密切的关联,而且该途径的产物为其他物质的合成提供了原料[42];有害物质的生物降解和代谢途径包括了苯甲酸降解、多环芳烃降解和二甲苯降解途径,这些途径与植物抗逆能力密切相关;氨基酸代谢与脱氨和转氨等作用息息相关,并且主要参与了植物氮素循环[43]。而本研究的结果表明,连作会减少根际细菌对碳源的利用、有害物质的降解及次生代谢物的合成代谢途径。尤其是相比于对照,连作6茬后的根际细菌氨基酸代谢相对丰度显著降低,这可能表明连作后根际环境不利于细菌的繁殖、代谢和合成物质,并且土壤养分循环可能受到一定程度的抑制。因此,在连作下,一些比较重要的代谢功能(如,碳水化合物代谢途径,有害物质的降解和次生代谢物的合成代谢途径)的减少或者丧失会是形成连作障碍的主要原因之一[44]。此外,我们利用FUNGuild来确定真菌分类群的营养模式。结果显示,相比于对照,连作6茬会显著增加病原真菌,这种真菌会通过损害宿主细胞而获取营养[45],并且营养亚型中植物病原体(如镰刀菌属)的相对丰度也会显著增加,这些也可能是引起连作障碍的主要原因之一。
3.2 西瓜连作对根际土壤细菌和真菌群落网络结构的影响
物种间相互作用是影响土壤病害的发生一种重要的生物因素,在群落构建过程中发挥重要作用[46],与关键生态过程和宿主健康密切相关[47]。本研究结果表明,多茬次的连作使细菌和真菌群落产生更复杂的潜在相互作用,这可能是因为植物残体量的增加,以及前茬作物产生的各种分泌物和遗传效应影响土壤微生物群落[48]。对照的网络直径和模块化均大于连作2茬和连作6茬,并且有研究指出具有更高模块化的微生物网络结构更能抵抗环境干扰[44,49]。因此,连作会导致根际微生物对环境变化的抵抗力减弱,容易引起连作障碍的发生。另外,本研究结果表明,在去除一定比例的网络节点后,连作6茬的网络稳定性指数急剧下降,表明连作使西瓜根际网络结构更加脆弱。因此,连作时长可能是影响根际网络稳定性的关键因素。
网络中潜在的关键物种在维持多物种共存和生态系统功能方面可能至关重要[50-51]。我们观察到,在对照的西瓜根际微生物网络中,关键类群有芽孢杆菌属()、鞘氨醇菌属()、剑菌属()和鞘氨醇单胞菌()等菌属。据报道芽孢杆菌属具有分解土壤中有机物的能力,如纤维素和木质素[52];鞘氨醇单胞菌属的成员具有降解复杂有机物质、拮抗植物病原真菌以及分泌胞外多糖等多种生态功能[53];鞘氨醇菌属能降解多环芳烃或酚类有机物[54],适应环境能力强[55];剑菌属能够作为有益菌接种至土壤中促进植物的生长[56],有助于维持植物激素平衡、控制根系发育、促进营养获取以及预防宿主植物疾病[57]。而在连作6茬后这些关键类群的相对丰度相比于对照发生了明显的减少,因此连作土壤病害的发生可能是由于关键类群的减少或者缺失。
3.3 西瓜连作对根际土壤细菌和真菌群落组装的影响
为了更好地了解微生物群落组成如何响应环境变化,我们探究了西瓜根际微生物群落组装的生态过程。在本研究中,当短期连作时(连作2茬),一些非生物环境因素(如pH、盐度、有机碳含量)的选择(环境过滤和物种间相互的拮抗与协同作用)会使细菌群落构建过程从随机性转变为确定性[20];但在较长连作茬次下,细菌种群更容易受到随机性事件的影响,这可以用体型-可塑性假说来解释[58],即较小的生物更可能受到扩散限制而不是物种分类的影响,因为较小的生物(如,细菌类群)具有更大的代谢可塑性和更大的环境耐受性,因此可以生活在不同的栖息地[59];相对于真菌的生活史策略而言,细菌类群的生存策略更加倾向于r策略,且在长期连作下,养分较为充足,细菌的生长没有受到环境的限制,趋向于随机性过程。而在真菌群落中,随机性过程在对照中更为突出,但连作后逐渐转变为确定性过程。这可能是一些环境变量对根际微生物群落的组装的潜在影响,如植物分泌物和残体的增加、土壤理化性质的变化等。环境因素的差异也可能随着连作茬次的增加而增加,并在根际微生物群落构建中发挥关键作用[60]。值得注意的是,当地环境的变化逐渐增加了真菌群落的确定性选择的重要性,而细菌群落的随机性却占主导,这可能是因为长期连作引起环境变化是由真菌性病害引起的(即由尖孢镰刀菌引起的西瓜枯萎病),植物可能会向环境释放特定的次生物质来招募更多的病原性真菌[12-14],真菌群落因此会受到更强的环境选择。同时细菌和真菌群落的MST值也呈现先降低后增大的趋势,表明随机性和确定性过程共同调控着连作环境下根际微生物群落的构建。
4 结论
多茬次的连作显著影响了西瓜根际细菌和真菌群落结构,包括有益菌属的相对丰度减少和致病菌属的相对丰度增加,也会导致微生物群落网络的相互作用更加复杂,但群落稳定性和对环境变化的抵抗性减弱。此外,细菌群落功能组成发生显著变化,包括有害物质的生物降解和代谢,氨基酸的代谢等功能基因相对丰度在长期连作后降低;而真菌群落中病原真菌的相对丰度在连作6茬后显著增多。多茬次的连作也会使微生物群落组装的机制发生变化,随机性和确定性过程共同调控着连作环境下根际微生物群落的构建。因此,在实际生产中,应注意通过轮作或施用微生物肥料来维持西瓜根际土壤微生态的稳定,以缓解连作障碍。本研究结果为理解连作与微生物群落结构差异的关系提供了新视角,并揭示了长期连作下微生物群落构建过程和潜在机制,同时为开发微生物肥料进行土壤改良,进而缓解连作障碍提供理论依据。
[1] 田晴, 高丹美, 李慧, 刘守伟, 周新刚, 吴凤芝. 小麦根系分泌物对西瓜连作土壤真菌群落结构的影响. 中国农业科学, 2020, 53(5): 1018-1028. doi:10.3864/j.issn.0578-1752.2020.05.013.
TIAN Q, GAO D M, LI H, LIU S W, ZHOU X G, WU F Z. Effects of wheat root exudates on the structure of fungi community in continuous cropping watermelon soil. Scientia Agricultura Sinica, 2020, 53(5): 1018-1028. doi:10.3864/j.issn.0578-1752.2020.05.013. (in Chinese)
[2] 耿士均, 刘刊, 商海燕, 权俊娇, 陆小平, 王波. 园艺作物连作障碍的研究进展. 北方园艺, 2012(7): 190-195.
GENG S J, LIU K, SHANG H Y, QUAN J J, LU X P, WANG B. Research progress of continuous cropping obstacle in horticultural plants. Northern Horticulture, 2012(7): 190-195. (in Chinese)
[3] MARTYN R. Fusarium wilt of watermelon: 120 years of research. Horticultural Reviews, 2014, 42: 349-442.
[4] 滕凯, 陈前锋, 周志成, 向青松, 张敏, 尹华群, 刘勇军. 烟草连作障碍与土壤理化性质及微生物多样性特征的关联. 微生物学通报, 2020, 47(9): 2848-2856.
TENG K, CHEN Q F, ZHOU Z C, XIANG Q S, ZHANG M, YIN H Q, LIU Y J. Effect of soil physical and chemical properties and microbial community on continuous cropping obstacles in tobacco field. Microbiology China, 2020, 47(9): 2848-2856. (in Chinese)
[5] YUAN J, WEN T, ZHANG H, ZHAO M L, PENTON C R, THOMASHOW L S, SHEN Q R. Predicting disease occurrence with high accuracy based on soil macroecological patterns of Fusarium wilt. The ISME Journal, 2020, 14(12): 2936-2950.
[6] DESSAUX Y, GRANDCLÉMENT C, FAURE D. Engineering the rhizosphere. Trends in Plant Science, 2016, 21(3): 266-278.
[7] BORDENSTEIN S R, THEIS K R. Host biology in light of the microbiome: Ten principles of holobionts and hologenomes. PLoS Biology, 2015, 13(8): e1002226.
[8] 张树生, 杨兴明, 茆泽圣, 黄启为, 徐阳春, 沈其荣. 连作土灭菌对黄瓜(Cucumis sativus)生长和土壤微生物区系的影响. 生态学报, 2007, 27(5): 1809-1817.
ZHANG S S, YANG X M, MAO Z S, HUANG Q W, XU Y C, SHEN Q R. Effects of sterilization on growth of cucumber plants and soil microflora in a continuous mono-cropping soil. Acta Ecologica Sinica, 2007, 27(5): 1809-1817. (in Chinese)
[9] BRADFORD M A, MCCULLEY R L, CROWTHER T W, OLDFIELD E E, WOOD S A, FIERER N. Cross-biome patterns in soil microbial respiration predictable from evolutionary theory on thermal adaptation. Nature Ecology & Evolution, 2019, 3(2): 223-231.
[10] BAHRAM M, HILDEBRAND F, FORSLUND S K, ANDERSON J L, SOUDZILOVSKAIA N A, BODEGOM P M, BENGTSSON-PALME J, ANSLAN S, COELHO L P, HAREND H, HUERTA-CEPAS J, MEDEMA M H, MALTZ M R, MUNDRA S, OLSSON P A, PENT M, PÕLME S, SUNAGAWA S, RYBERG M, TEDERSOO L, BORK P. Structure and function of the global topsoil microbiome. Nature, 2018, 560(7717): 233-237.
[11] GAO Z Y, HAN M K, HU Y Y, LI Z Q, LIU C F, WANG X, TIAN Q, JIAO W J, HU J M, LIU L F, GUAN Z J, MA Z M. Effects of continuous cropping of sweet potato on the fungal community structure in rhizospheric soil. Frontiers in Microbiology, 2019, 10: 2269.
[12] HU L F, ROBERT C A M, CADOT S, ZHANG X, YE M, LI B B, MANZO D, CHERVET N, STEINGER T, VAN DER HEIJDEN M G A, SCHLAEPPI K, ERB M. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nature Communications, 2018, 9: 2738.
[13] LI X G, DING C F, HUA K, ZHANG T L, ZHANG Y N, ZHAO L, YANG Y R, LIU J G, WANG X X. Soil sickness of peanuts is attributable to modifications in soil microbes induced by peanut root exudates rather than to direct allelopathy. Soil Biology and Biochemistry, 2014, 78: 149-159.
[14] XU W H, LIU D, WU F Z, LIU S W. Root exudates of wheat are involved in suppression of Fusarium wilt in watermelon in watermelon-wheat companion cropping. European Journal of Plant Pathology, 2015, 141(1): 209-216.
[15] 曾维爱, 杨昭玥, 黄洋, 谷亚冰, 陶界锰, 刘勇军, 谢鹏飞, 蔡海林, 尹华群. 长期连作农田土壤细菌群落结构和共现网络拓扑性质对土壤理化性质的响应. 微生物学报, 2022, 62(6): 2403-2416.
ZENG W A, YANG Z Y, HUANG Y, GU Y B, TAO J M, LIU Y J, XIE P F, CAI H L, YIN H Q. Response of soil bacterial community structure and co-occurrence network topology properties to soil physicochemical properties in long-term continuous cropping farmland. Acta Microbiologica Sinica, 2022, 62(6): 2403-2416. (in Chinese)
[16] NING D L, DENG Y, TIEDJE J M, ZHOU J Z. A general framework for quantitatively assessing ecological stochasticity. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(34): 16892-16898.
[17] GRYTA A, FRĄC M, OSZUST K. The application of the biolog EcoPlate approach in ecotoxicological evaluation of dairy sewage sludge. Applied Biochemistry and Biotechnology, 2014, 174(4): 1434-1443.
[18] GUO J J, LIU W B, ZHU C, LUO G W, KONG Y L, LING N, WANG M, DAI J Y, SHEN Q R, GUO S W. Bacterial rather than fungal community composition is associated with microbial activities and nutrient-use efficiencies in a paddy soil with short-term organic amendments. Plant and Soil, 2018, 424(1): 335-349.
[19] GUO H P, DONG P S, GAO F, HUANG L, WANG S P, WANG R Y, YAN M C, ZHANG D M. Sucrose addition directionally enhances bacterial community convergence and network stability of the shrimp culture system. NPJ Biofilms and Microbiomes, 2022, 8: 22.
[20] CHEN Q L, HU H W, YAN Z Z, LI C Y, NGUYEN B A T, SUN A Q, ZHU Y G, HE J Z. Deterministic selection dominates microbial community assembly in termite mounds. Soil Biology and Biochemistry, 2021, 152: 108073.
[21] BURNS A R, STEPHENS W Z, STAGAMAN K, WONG S, RAWLS J F, GUILLEMIN K, BOHANNAN B J. Contribution of neutral processes to the assembly of gut microbial communities in the zebrafish over host development. The ISME Journal, 2016, 10(3): 655-664.
[22] TOJU H, OKAYASU K, NOTAGUCHI M. Leaf-associated microbiomes of grafted tomato plants. Scientific Reports, 2019, 9: 1787.
[23] LING N, SONG Y, RAZA W, HUANG Q W, GUO S W, SHEN Q R. The response of root-associated bacterial community to the grafting of watermelon. Plant and Soil, 2015, 391(1): 253-264.
[24] SCHLOSS P D, WESTCOTT S L, RYABIN T, HALL J R, HARTMANN M, HOLLISTER E B, LESNIEWSKI R A, OAKLEY B B, PARKS D H, ROBINSON C J, SAHL J W, STRES B, THALLINGER G G, VAN HORN D J, WEBER C F. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Applied and Environmental Microbiology, 2009, 75(23): 7537-7541.
[25] NGUYEN N H, SONG Z W, BATES S T, BRANCO S, TEDERSOO L, MENKE J, SCHILLING J S, KENNEDY P G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecology, 2016, 20: 241-248.
[26] DENG Y, JIANG Y H, YANG Y F, HE Z L, LUO F, ZHOU J Z. Molecular ecological network analyses. BMC Bioinformatics, 2012, 13: 113.
[27] SHI S J, NUCCIO E E, SHI Z J, HE Z L, ZHOU J Z, FIRESTONE M K. The interconnected rhizosphere: high network complexity dominates rhizosphere assemblages. Ecology Letters, 2016, 19(8): 926-936.
[28] PENG G S, WU J. Optimal network topology for structural robustness based on natural connectivity. Physica A: Statistical Mechanics and Its Applications, 2016, 443: 212-220.
[29] LI X G, JOUSSET A, DE BOER W, CARRIÓN V J, ZHANG T L, WANG X X, KURAMAE E E. Legacy of land use history determines reprogramming of plant physiology by soil microbiome. The ISME Journal, 2019, 13(3): 738-751.
[30] ZHU S Y, WANG Y Z, XU X M, LIU T M, WU D Q, ZHENG X, TANG S W, DAI Q Z. Potential use of high-throughput sequencing of soil microbial communities for estimating the adverse effects of continuous cropping on ramie (L. Gaud). PLoS ONE, 2018, 13(5): e0197095.
[31] 李晶晶, 续勇波. 连作年限对设施百合土壤微生物多样性的影响. 土壤通报, 2020, 51(2): 343-351.
LI J J, XU Y B. Effects of continuous cropping years of lily on soil microbial diversities under greenhouse cultivation. Chinese Journal of Soil Science, 2020, 51(2): 343-351. (in Chinese)
[32] VAN BRUGGEN A H C, SEMENOV A M. In search of biological indicators for soil health and disease suppression. Applied Soil Ecology, 2000, 15(1): 13-24.
[33] ZHANG Y, CAO C Y, PENG M, XU X J, ZHANG P, YU Q J, SUN T. Diversity of nitrogen-fixing, ammonia-oxidizing, and denitrifying bacteria in biological soil crusts of a revegetation area in Horqin Sandy Land, Northeast China. Ecological Engineering, 2014, 71: 71-79.
[34] MUNOZ R, TEELING H, AMANN R, ROSSELLÓ-MÓRA R. Ancestry and adaptive radiation of Bacteroidetes as assessed by comparative genomics. Systematic and Applied Microbiology, 2020, 43(2): 126065.
[35] KWAK M J, KONG H G, CHOI K, KWON S K, SONG J Y, LEE J, LEE P A, CHOI S Y, SEO M, LEE H J, JUNG E J, PARK H, ROY N, KIM H, LEE M M, RUBIN E M, LEE S W, KIM J F. Rhizosphere microbiome structure alters to enable wilt resistance in tomato. Nature Biotechnology, 2018, 36(11): 1100-1109.
[36] 王涛, 蓝慧, 田云, 卢向阳. 多环芳烃的微生物降解机制研究进展. 化学与生物工程, 2016, 33(2): 8-14.
WANG T, LAN H, TIAN Y, LU X Y. Research progress on microbial degradation mechanisms for polycyclic aromatic hydrocarbons. Chemistry & Bioengineering, 2016, 33(2): 8-14. (in Chinese)
[37] 姜英华, 胡白石, 刘凤权. 植物土传病原菌拮抗细菌的筛选与鉴定. 中国生物防治, 2005, 21(4): 260-264.
JIANG Y H, HU B S, LIU F Q. Selection and identification of antagonistic bacteria against soil-borne plant pathogens. Chinese Journal of Biological Control, 2005, 21(4): 260-264. (in Chinese)
[38] 吴凤芝, 王学征. 设施黄瓜连作和轮作中土壤微生物群落多样性的变化及其与产量品质的关系. 中国农业科学, 2007, 40(10): 2274-2280. doi:10.3321/j.issn: 0578-1752.2007.10.021.
WU F Z, WANG X Z. Effect of monocropping and rotation on soil microbial community diversity and cucumber yield, quality under protected cultivation. Scientia Agricultura Sinica, 2007, 40(10): 2274-2280. doi:10.3321/j.issn: 0578-1752.2007.10.021. (in Chinese)
[39] PLIEGO C, RAMOS C, DE VICENTE A, CAZORLA F M. Screening for candidate bacterial biocontrol agents against soilborne fungal plant pathogens. Plant and Soil, 2011, 340(1): 505-520.
[40] ČÍŽKOVÁ D, ŠRŮTKA P, KOLAŘÍK M, KUBÁTOVÁ A, PAŽOUTOVÁ S. Assessing the pathogenic effect of Fusarium, Geosmithia and Ophiostoma fungi from broad-leaved trees. Folia Microbiologica, 2005, 50(1): 59-62.
[41] CHEN M N, LI X, YANG Q L, CHI X Y, PAN L J, CHEN N, YANG Z, WANG T, WANG M, YU S L. Soil eukaryotic microorganism succession as affected by continuous cropping of peanut: Pathogenic and beneficial fungi were selected. PLoS ONE, 2012, 7(7): e40659.
[42] 徐伟, 葛阳阳, 陈翠婷, 马婷婷. 基于宏基因组技术分析传统红茶菌中菌群组成及其主要代谢通路. 食品工业科技, 2018, 39(5): 119-123, 129.
XU W, GE Y Y, CHEN C T, MA T T. Microorganism composition and main metabolic pathways analysis of traditional Kombucha by metagenomic technology. Science and Technology of Food Industry, 2018, 39(5): 119-123, 129. (in Chinese)
[43] 丁钰珮, 杜宇佳, 高广磊, 张英, 曹红雨, 朱宾宾, 杨思远, 张儆醒, 邱业, 刘惠林. 呼伦贝尔沙地樟子松人工林土壤细菌群落结构与功能预测. 生态学报, 2021, 41(10): 4131-4139.
DING Y P, DU Y J, GAO G L, ZHANG Y, CAO H Y, ZHU B B, YANG S Y, ZHANG J X, QIU Y, LIU H L. Soil bacterial community structure and functional prediction ofvar. mongolica plantations in the Hulun Buir Sandy Land. Acta Ecologica Sinica, 2021, 41(10): 4131-4139. (in Chinese)
[44] 李晓雪. 辣椒对土壤连作障碍的响应及其成因研究[D]. 保定: 河北农业大学, 2021.
LI X X. The mechanisms of continuous cropping obstacle formation of chilli pepper and the response of chilli pepper to continuously mono-cropped system[D]. Baoding: Hebei Agricultural University, 2021. (in Chinese)
[45] TOJU H, KISHIDA O, KATAYAMA N, TAKAGI K. Networks depicting the fine-scale co-occurrences of fungi in soil horizons. PLoS ONE, 2016, 11(11): e0165987.
[46] KONOPKA A, LINDEMANN S, FREDRICKSON J. Dynamics in microbial communities: Unraveling mechanisms to identify principles. The ISME Journal, 2015, 9(7): 1488-1495.
[47] LAYEGHIFARD M, HWANG D M, GUTTMAN D S. Disentangling interactions in the microbiome: A network perspective. Trends in Microbiology, 2017, 25(3): 217-228.
[48] NUCCIO E E, STARR E, KARAOZ U, BRODIE E L, ZHOU J Z, TRINGE S G, MALMSTROM R R, WOYKE T, BANFIELD J F, FIRESTONE M K, PETT-RIDGE J. Niche differentiation is spatially and temporally regulated in the rhizosphere. The ISME Journal, 2020, 14(4): 999-1014.
[49] YUAN M M, GUO X, WU L W, ZHANG Y, XIAO N J, NING D L, SHI Z, ZHOU X S, WU L Y, YANG Y F, TIEDJE J M, ZHOU J Z. Climate warming enhances microbial network complexity and stability. Nature Climate Change, 2021, 11(4): 343-348.
[50] HERREN C M, MCMAHON K D. Keystone taxa predict compositional change in microbial communities. Environmental Microbiology, 2018, 20(6): 2207-2217.
[51] CHEN L J, JIANG Y J, LIANG C, LUO Y, XU Q S, HAN C, ZHAO Q G, SUN B. Competitive interaction with keystone taxa induced negative priming under biochar amendments. Microbiome, 2019, 7(1): 77.
[52] SUN X L, XU Z H, XIE J Y, HESSELBERG-THOMSEN V, TAN T M, ZHENG D Y, STRUBE M L, DRAGOŠ A, SHEN Q R, ZHANG R F, KOVÁCS Á T.stimulates resident rhizospherefor plant health through metabolic interactions. The ISME Journal, 2022, 16(3): 774-787.
[53] WHITE D C, SUTTON S D, RINGELBERG D B. The genus Sphingomonas: Physiology and ecology. Current Opinion in Biotechnology, 1996, 7(3): 301-306.
[54] 阎洁, 余雪巍, 李鉴博, 顾海萍, 郭二辉. 一株菲降解细菌产生生物表面活性剂特性的研究. 生态环境学报, 2021, 30(8): 1683-1694.
YAN J, YU X W, LI J B, GU H P, GUO E H. Research on the characterization of surfactant produced by a phenanthrene-degrading strain. Ecology and Environmental Sciences, 2021, 30(8): 1683-1694. (in Chinese)
[55] ZHOU X Y, LIANG Y, REN G F, ZHENG K W, WU Y, ZENG X Y, ZHONG Y, YU Z Q, PENG P A. Biotransformation of tris(2- chloroethyl) phosphate (TCEP) in sediment microcosms and the adaptation of microbial communities to TCEP. Environmental Science & Technology, 2020, 54(9): 5489-5497.
[56] SCHMITZ L, YAN Z C, SCHNEIJDERBERG M, DE ROIJ M, PIJNENBURG R, ZHENG Q, FRANKEN C, DECHESNE A, TRINDADE L M, VAN VELZEN R, BISSELING T, GEURTS R, CHENG X. Synthetic bacterial community derived from a desert rhizosphere confers salt stress resilience to tomato in the presence of a soil microbiome. The ISME Journal, 2022, 16(8): 1907-1920.
[57] XU J, ZHANG Y Z, ZHANG P F, TRIVEDI P, RIERA N, WANG Y Y, LIU X, FAN G Y, TANG J L, COLETTA-FILHO H D, CUBERO J, DENG X L, ANCONA V, LU Z J, ZHONG B L, ROPER M C, CAPOTE N, CATARA V, PIETERSEN G, VERNIÈRE C, AL-SADI A M, LI L, YANG F, XU X, WANG J, YANG H M, JIN T, WANG N. The structure and function of the global citrus rhizosphere microbiome. Nature Communications, 2018, 9: 4894.
[58] ISABWE A, YAO H F, ZHANG S X, JIANG Y J, BREED M F, SUN X. Spatial assortment of soil organisms supports the size-plasticity hypothesis. ISME Communications, 2022, 2: 102.
[59] WU W X, LU H P, SASTRI A, YEH Y C, GONG G C, CHOU W C, HSIEH C H. Contrasting the relative importance of species sorting and dispersal limitation in shaping marine bacterial versus protist communities. The ISME Journal, 2018, 12(2): 485-494.
[60] DELGADO-BAQUERIZO M, MAESTRE F T, REICH P B, JEFFRIES T C, GAITAN J J, ENCINAR D, BERDUGO M, CAMPBELL C D, SINGH B K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nature Communications, 2016, 7: 10541.
Characteristics and Succession of Rhizosphere Soil Microbial Communities in Continuous Cropping Watermelon
GUO HanYue1, WANG DongSheng2, RUAN Yang1, QIAO YiZhu1, ZHANG YunTao1, LI Ling1, HUANG QiWei1, GUO ShiWei1, LING Ning1, SHEN QiRong1
1College of Resources and Environment Science, Nanjing Agricultural University/Key Laboratory of Organic-based Fertilizers of China/Jiangsu Provincial Key Laboratory for Solid Organic Waste Utilization, Nanjing 210095;2Nanjing Institute of Vegetable Science, Nanjing 210042
【Objective】The aim of this study was to investigate the effects of continuous cropping on the construction and potential functions of bacterial and fungal communities in the rhizosphere soil of watermelon, and to clarify the adaptability of rhizosphere microorganisms to environmental changes, so as to provide a theoretical basis for ecological control of watermelon continuous cropping obstacles and healthy maintenance of farmland. 【Method】In this study, the rhizosphere soil of watermelon without continuous cropping (CK), continuous cropping for 2 times and continuous cropping for 6 times was used as the research object. 16S rRNA and ITS high-throughput sequencing were used to analyze the effects of continuous cropping on the bacterial and fungal communities of rhizosphere soil of watermelon. 【Result】With the increasing continuous cropping times, the bacterial diversity index in the rhizosphere soil of watermelon showed a trend of first decreasing and then increasing, while the fungal diversity index decreased significantly. At the bacterial genus level, multiple times of continuous cropping decreased the relative abundance ofandin watermelon rhizosphere soil; at the fungal genus level, the relative abundance ofincreased with the continuous cropping times. Compared with CK, the network complexity of continuous cropping was higher, but the stability of network structure was lower. In addition, compared with CK, the relative abundance of biodegradation pathways of harmful substances and metabolic pathways of amino acids could be significantly reduced after continuous cropping for 6 times; the relative abundance of pathogenic fungi significantly increased in the fungal community after continuous cropping for 6 times. During community succession, the stochastic processes dominated rhizosphere bacterial community construction in watermelon under continuous cropping, while the deterministic processes dominated rhizosphere fungal community construction in watermelon. 【Conclusion】Continuous cropping caused changes in community characteristics, functional composition and succession process of rhizosphere bacteria and fungi. The decrease of key functions of bacterial community, the increase of pathologic fungi and the decrease of stability of microbial community network might be the important factors leading to occurrence of watermelon continuous cropping obstacles.
watermelon; continuous cropping soil; rhizosphere microbial community; function prediction; community succession
2022-11-19;
2023-01-09
国家自然科学基金(31972506)
郭晗玥,E-mail:2020103110@stu.njau.edu.cn。通信作者凌宁,E-mail:nling@njau.edu.cn
(责任编辑 李云霞)
