海拔和遮阴对缺苞箭竹大小的影响
2023-11-13朱育旗夏勇
朱育旗,夏勇
1.四川林业集团有限公司,四川 成都 610041;2.四川省林业物资供销有限公司,四川 成都 610081
缺苞箭竹(Fargesia denudata)自然分布于甘肃南部、四川北部海拔1 900~3 200 m 的针阔叶或针叶林混交林下,是岷山北部大熊猫重要的主食竹之一[1]和重要林下层片。对缺苞箭竹的大量研究始于20 世纪80 年代岷山竹类开花,20 世纪的研究主要包括缺苞箭竹的生态特性[2]与分布[3]、种子贮存[4]、幼苗生长[5]、物候[6]和天然更新[7];21 世纪初则集中于土壤养分[8,9]、生物量分配及养分贮量[10-12]、叶片养分再分配[13]、凋落物养分归还及养分利用效率的影响[14]、种子生理[15]、2004 年缺苞箭竹开花情况[16]、克隆生长[17]、控制实验中光强对缺苞箭竹叶片衰老生理[18]和气体交换参数(光合生理)[19]的影响;以及最近缺苞箭竹的重金属富集能力[20]、营养成分和氨基酸的海拔差异[21]、饲用品质[22]、林窗微环境对缺苞箭竹气体交换参数[23]和可食性的影响[24]、春季干旱条件下的氮分配策略[25]和未来全球变化下的分布情景[26]。
但这些研究没有回答“缺苞箭竹的大小及其协调性是否受海拔和遮阴的影响?”这一基本问题。植物形态需要一定的协调性,以维持其机械稳定性[27,28]和对抗风暴[29]、冰雪等自然灾害的影响[30]。但植物形状协调性的研究多集中在受灾频繁地区的优势树种(如风暴[29]和大雪[31])、功能性状重要组成的小枝[27,28]和叶片[32]亦有涉及,少有研究涉及下层植物的植株。因此,考虑到可达性和易操作性、在对老河沟自然保护区充分踏查后,本文以较高和较低海拔、3 种遮阴生境下的缺苞箭竹为对象,在群落调查的基础上,阐明海拔和遮阴对缺苞箭竹大小及其协调性的作用,以期为老河沟乃至岷山北部大熊猫栖息地的竹类定向管理提供基础数据和科学依据。
1 材料与方法
1.1 研究区概况
研究地位于四川省平武县境内的老河沟自然保护区(下文简称老河沟)[33],北纬32°36'22''—32°25'52''、东经104°32'42''—104°45'25'',海拔1 250~3 504 m,总面积约110 km2;属北亚热带山地湿润季风气候,冬季干燥寒冷,盛夏湿润凉爽,7 月平均温度最高(23.37℃),1 月温度最低(平均2.77℃),7 月水温最高(平均25℃),年均温13.7℃左右;原有植被破坏后,1 800 m 以下低中山河谷天然更新形成野核桃(Juglanscathayensis)林为主的低山次生落叶阔叶林、海拔较高的地段常形成水青冈(Faguslongipetiolata)、椴树(Tilia tuan)、桦木(Betulaspp.)、槭树(Acerspp.)等落叶树种的常绿与落叶阔叶混交林,1 800—2 600 m 地段形成大量的桦木等次生落叶阔叶林,2 600~3 200 m 的亚高山地区,尚有部分以岷江冷杉(Abiesfaxoniana)、麦吊云杉(Picea brachytyla)为优势种的暗针叶林,3 200 m以上为亚高山草甸和流石滩植被。
1.2 样地布设与调查
在踏查的基础上,2014 年4 月选择老河沟主沟阴坡中下部的次生林为对象,在缺苞箭竹分布的较高(约2 995 m,后文简称高海拔)和较低海拔(2 142 m,后文简称低海拔)地段边缘选择坡向、坡度和坡位基本一致、易到达的、郁闭度相当的森林区域,乔木层主要树种为落叶阔叶的椴树、红桦(B.albosinensis)和槭树。在林下、林缘和全光照3 个遮阴梯度各设置3 个5 m×5 m 样方,对其中的缺苞箭竹进行群落调查,之后在其中选择中等大小的30 个有效植株(生长正常、未被大熊猫取食)实测其基径(0.01 mm,得力DL91300,德利集团有限公司,浙江宁波)和株高(0.01 cm,得力7.5 m 高精度钢卷尺,德利集团有限公司,浙江宁波)。鉴于缺苞箭竹的年龄较难确定,将所有植株作为一个整体来了解其大小在海拔和遮阴梯度上的变化。借鉴树木[30]或小枝[27]纤细率(slenderness)来定义缺苞箭竹的形状指数,即形状指数=株高/基径。
1.3 数据分析
对数据进行正态性和方差同质性检验,如果不符合正态分布则将各参数进行以10 为底的对数转换,使之符合正态分布。绘制箱图展现缺苞箭竹基径、株高和形状指数的总体分布,采用单因素方差分析(One-way Anova)检验海拔和遮阴对缺苞箭竹基径、株高和形状指数的影响,用LSD 法(Fisher's Least Significant Difference)对基径、株高和形状指数在遮阴和海拔间的差异进行事后比较;用一般线性模型(General Linear Model,GLM)分析海拔遮阴的交互作用对基径、株高和形状指数的影响。以变异系数CV(coefficient of variation,CV%=标准差/平均值×100%)作为基径、株高和形状指数的可塑性指数[34]。采用皮尔逊相关性分析各性状的相关性。指标间总体显著相关的再用标准化主轴回归(Standardised Major Axis regression,SMA)探究其异速生长关系[35]:在SMATR v2.0(Standardised Major Axis Tests &Routines,http://www.bio.mq.edu.au/ecology/SMATR/)上以log 转换后计算指标间的异速生长指数和常数,检验斜率a的异质性,当斜率同质时检验y轴截距和斜率的变化并进行多重比较。其他分析在IBM SPSS 20.0(IBM Corp.,Armonk,NY,USA)上进行,绘图在OriginPro 2022(OriginLab Corp.,Northampton,MA,USA)上进行。所有数据均为平均值±标准差,各指数的分布范围为95%的置信区间(Confidence interval,95%CI),分析的显著性设定为0.05。
2 结果与分析
2.1 缺苞箭竹的基径和株高
老河沟缺苞箭竹的基径总体为8.65 ± 1.67 mm、介于8.47~8.82 mm 之间(见图1A,左边为箱图、右边为数据分布,下同),株高(见图1C)和形状指数(见图1E)分别为243.2 ± 62.8 cm 和282.4 ±53.6、介于236.7~249.7 cm 和276.8~287.9 之间。这三个指标的变异系数分别为19.3%、25.8%和19%,表明缺苞箭竹的大小和形状在老河沟均具有一定的可塑性。
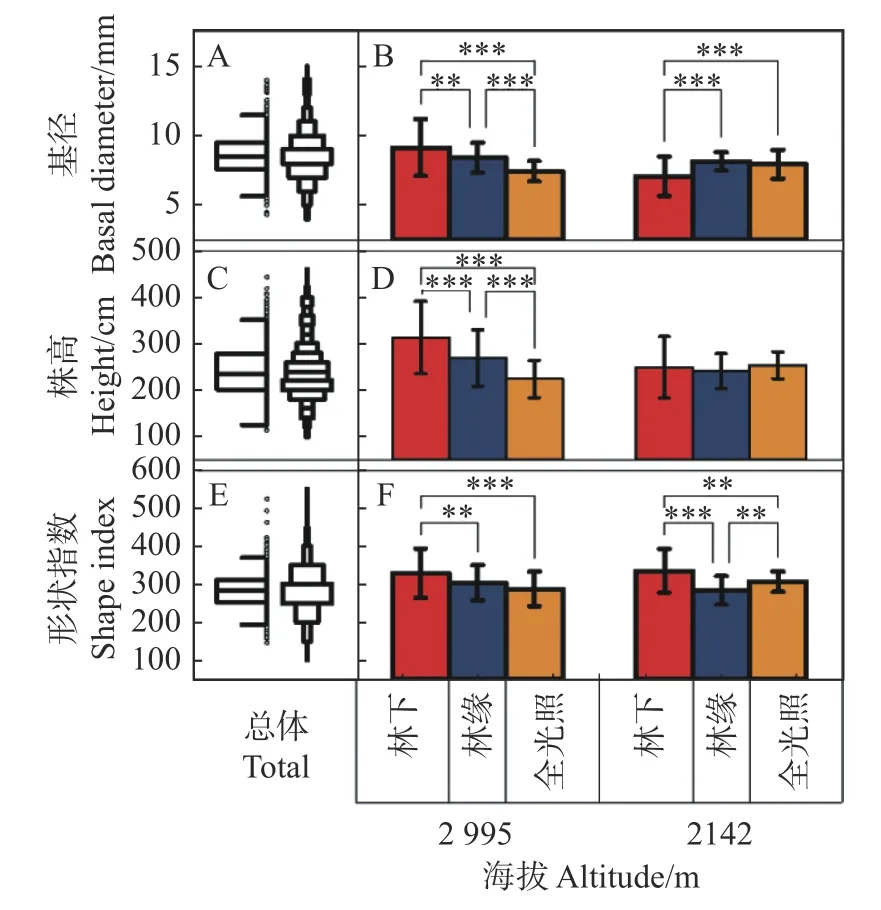
图1 老河沟缺苞箭竹的大小和形状Fig.1 The plant size and shape index of Fargesia denudata within Laohegou Nature Reserve
2.2 海拔和遮阴对植株大小的影响
老河沟各生境缺苞箭竹的基径及在同一海拔的多重比较见图1B。高海拔林下的基径最大(9.97±2.44 mm),极显著大于其他5 种生境(所有P<0.01;后文中所有极显著均为P<0.01,不再标注);高海拔林缘次之(9.11±1.29 mm),稍大于低海拔林缘(8.78±0.79 mm,P=0.228)、显著大于低海拔全光照(8.54±1.24 mm,P=0.037),与其他几种生境均有极显著差异。低海拔林下的则为最小(7.55±1.63 mm),稍小于高海拔全光照(7.93±0.86 mm,P=0.162),极显著小于其他生境。GLM 分析结果表明海拔、遮阴及其交互作用对基径均有极显著影响(见表1)。总体上(即数据合并后,下同),高海拔的基径(9.00±1.86mm)极显著大于低海拔(8.29±1.37 mm);林缘的基径(8.94±1.08 mm)稍大于(P=0.401)林下(8.76±2.40 mm),林缘极显著大于全光照(8.23±1.11 mm)、林下显著大于全光照(P=0.013)。
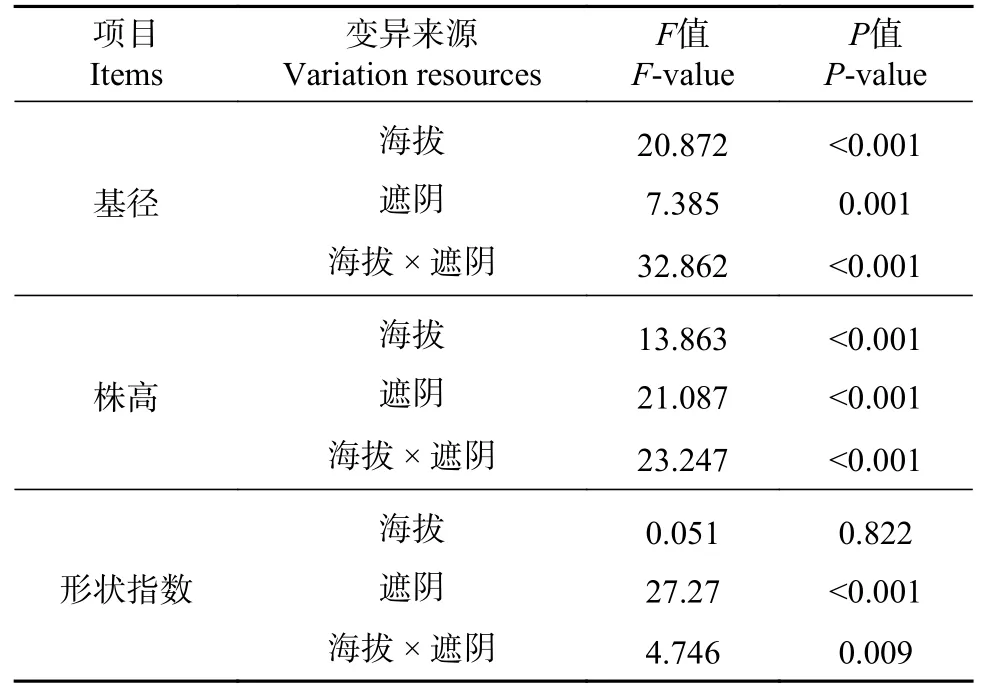
表1 基于一般线性模型的海拔和遮阴对缺苞箭竹的影响Tab.1 Effects of altitude and shade (F-and P-value) on Fargesia denudata based on a general linear model
老河沟各生境缺苞箭竹的株高及在同一海拔的多重比较见图1D。高海拔林下的植株最高(301.8±79.5 cm),极显著大于其他5 种生境。高海拔林缘次之(253.6±64.5 cm)、稍高于低海拔的林下(234.8±64.7 cm,P=0.065)和全光照(237.2±30.6 cm,P=0.109)、与其他几种生境均有极显著差异;以高海拔全光照下的植株最矮(207.0±40.2 cm),略矮于低海拔林缘(224.8±36.9 cm)、极显著矮于其他生境。GLM 分析结果表明海拔、遮阴及其交互作用对植株高度均有极显著影响(表1)。总体上,高海拔的植株(254.1±74.1 cm)极显著高于低海拔(232.3±46.5 cm);林下的植株(268.3±79.6 cm)极显著高于全光照(222.1±38.6 cm)和林缘(239.2±54.3 cm),林缘显著高于全光照(P=0.028)。
老河沟各生境缺苞箭竹的形状指数及在同一海拔的多重比较见图1F。低海拔林下的缺苞箭竹的形状指数最大(312.4±57.9),稍大于高海拔林下(306.8±67.1,P=0.533)、极显著大于其他4 种生境;高海拔林下的形状指数极显著大于其他4 种生境;低海拔林缘的形状指数最小(256.5±36.9)、稍小于高海拔全光照(261.7±46.5,P=0.569),极显著小于其他几种生境。GLM 分析结果表明遮阴、海拔和遮阴的交互作用对形状指数有极显著影响(见表1)。总体上,高海拔的形状指数(281.8±58.2)略低于低海拔(283.0±48.7,P=0.835);林下的形状指数(309.6±62.5)极显著大于林缘(266.7±45.2)和全光照(270.8±39.8)生境,后两者间无显著差异(P=0.525)。
2.3 箭竹大小与形状的关系
总体上,株高与基径(r=0.699)、形状指数与株高(r=0.625)均有极显著的相关关系,形状指数与基径有显著的负相关(r=-0.109,P=0.039)。lgS-lgD 的SMA 分析表明:总体上没有共斜率(P<0.01),仅高海拔全光照的lgS-lgD 有极显著的负线性关系(R2=0.242,斜率为-0.7553、介于-0.9477和-0.6020 之间且与-1 有显著差异、P=0.016;截距为3.145、介于2.984 和3.305 之间),其他的lgSlgD 均无线性关系(所有的P>0.05)。
lgH-lgD 的SMA 分析表明(见表2、图2A 和图2C):总体上没有共斜率(P<0.01),各组lgSlgD 均有极显著的异速关系,所有的斜率均为正,表明缺苞箭竹的植株高度与基径成正相关,除低海拔-全光照的斜率小于1、但与1 没有显著的差异(P=0.321),其他的均大于1、且与1 有显著或极显著差异。高海拔-全光照的斜率最大,稍大于低海拔-林缘(P=0.784)和高海拔-林缘(P=0.858)、显著大于低海拔-林下(P=0.011)、极显著大于其他2 种生境;低海拔-全光照的最小,显著或极显著低于其他生境。截距以低海拔-全光照、高海拔-林下和低海拔-林下较高,另3 种生境则较小。
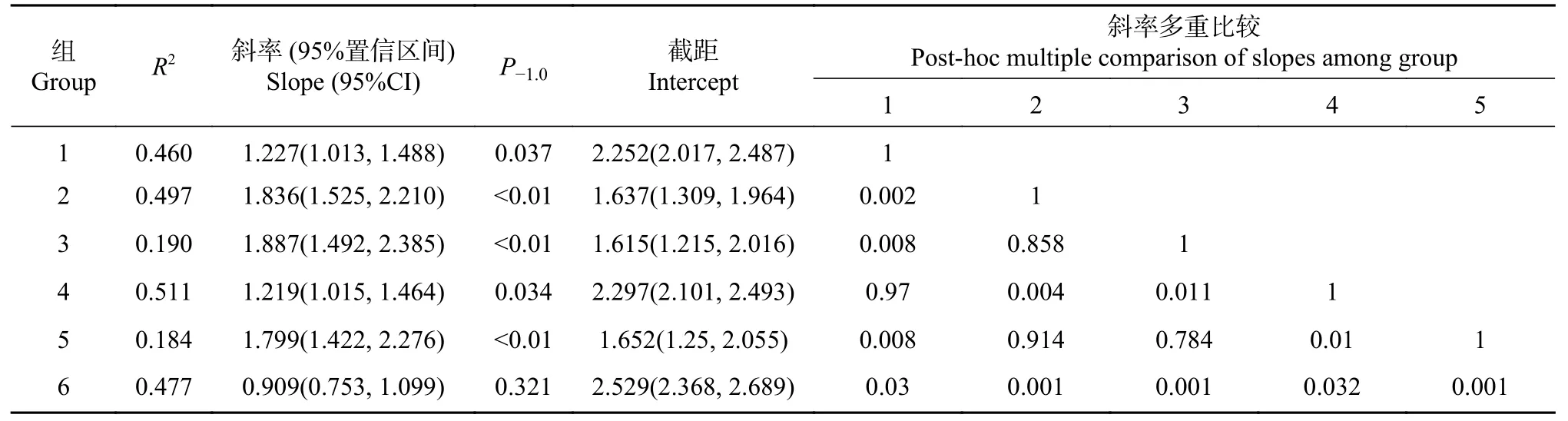
表2 缺苞箭竹基径与株高间关系的SMA 分析Tab.2 Relationships between height and base diameter of Fargesia denudata altitude based on SMA
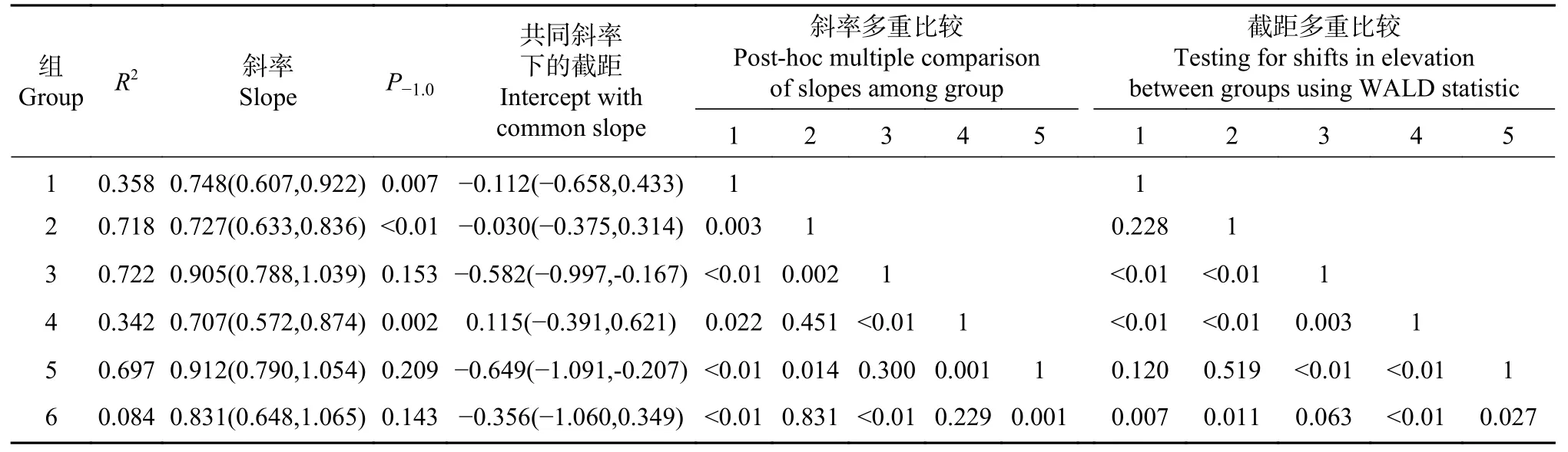
表3 低海拔缺苞箭竹高度与形状间关系的SMA 分析Tab.3 Relationships between height and shape index of Fargesia denudata based on SMA
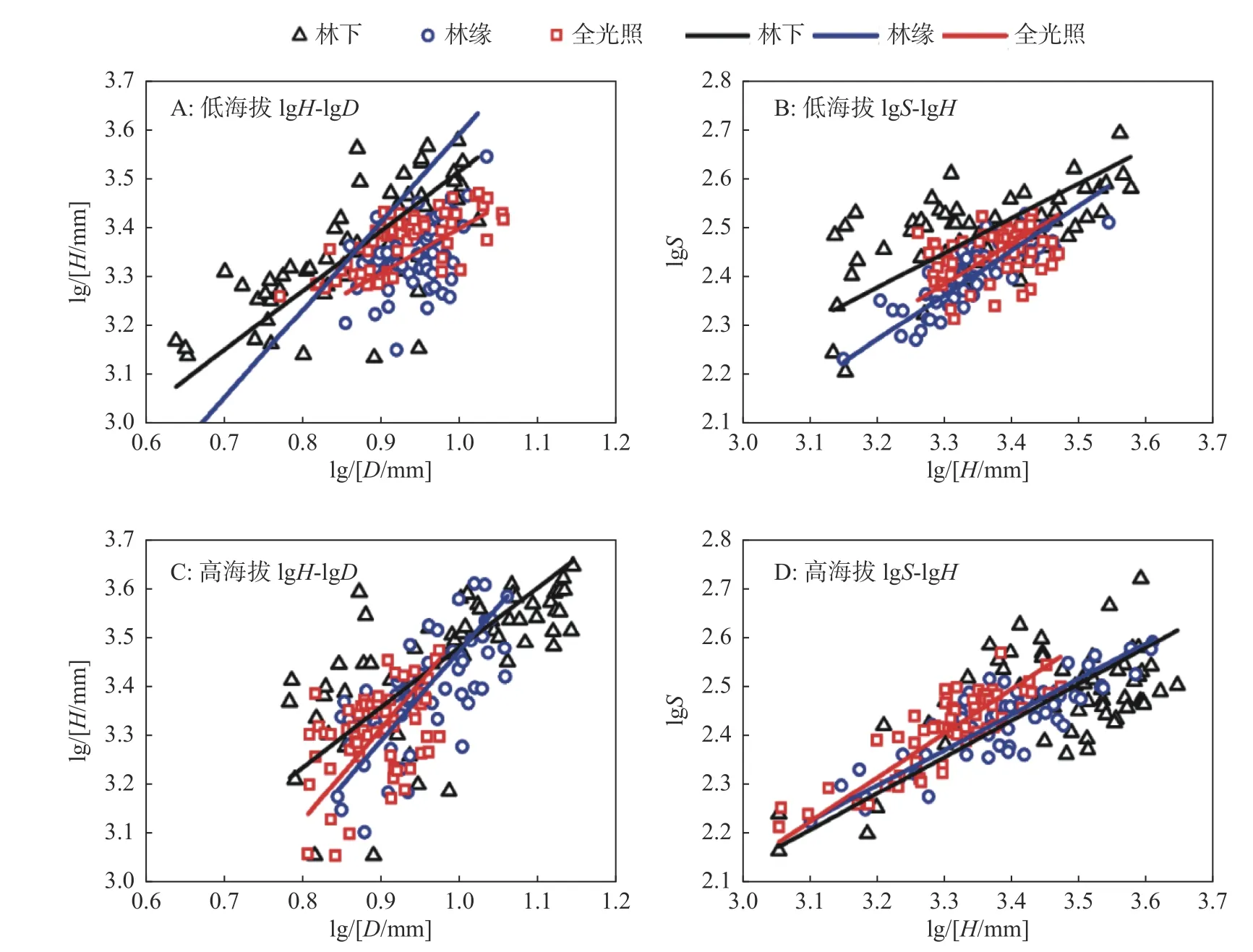
图2 缺苞箭竹基径-株高、株高-形状指数间的异速关系Fig.2 Allometric relationships between height-base diameter and height-shape index of Fargesia denudata based on SMA
lgS-lgH 的SMA 分析表明(见表2、图2B 和图2D):总体上有共斜率(P=0.097)0.816(0.760~0.875),且极显著小于1,即总体上表现为收益递减。所有斜率均小于1,但高海拔-全光照、低海拔-全光照和低海拔-林缘的斜率稍小于1,另3 种生境的斜率则极显著小于1。低海拔-林缘的斜率最大,稍大于高海拔-全光照和低海拔-全光照、显著大于高海拔-林缘、极显著大于其他2 种生境,低海拔-林下的斜率最小。截距则以低海拔-林下为最大、低海拔-林缘为最小。
3 讨论与结论
缺苞箭竹的基径稍大于王朗的更新幼苗竹、植株高度则稍矮,株高随基径的增速不如王朗的更新幼竹[36],原因可能是缺苞箭竹为更新幼苗、更倾向于将资源投入到长高以更好地捕获散射光。缺苞箭竹的属于典型的高寒地区小径竹,老河沟植株大小及其变异系数、以及基径-株高线性关系的斜率均远小于中国竹类的平均水平,主要是因为全国竹类众多[37]、秆形状粗大的竹类亦有较大比重[38]。
从全光照到林下,其生境变化类似于林窗中心到林下,其温度和光照逐渐降低、土壤养分也因此而发生变化[39]。这种变化将直接影响植物的光合生理[23]、生物量形成与分配[40,41]、幼苗存活[42]与种群动态[43],进而对下层植物和森林更新产生深刻影响。尽管老河沟高海拔林下的缺苞箭竹个体粗大,但总体仍表现为林缘的个体更粗壮,即缺苞箭竹需要一定的遮阴,这与王朗国家级自然保护区,3 000 m 左右的小林窗有利于形成粗壮的缺苞箭竹个体基本一致[17]。海拔和遮阴对缺苞箭竹高度的影响则较小,毕竟要有一定高度更有利于获得光照。无论林下还是林缘,有遮阴庇护的缺苞箭竹个体一般高于全光照、形状指数也更大,高海拔的全光照下尤其如此。这也反应的缺苞箭竹对全光照生境的适应:全光照的植物个体在冬季将承受更严重的冰雪压力,植株个体通常更加粗和具有更小的形状指数(纤细率)。这与乔木树种的表现总体一致,即纤细率越小的个体承受雪压等自然胁迫的能力越强[30,31]。
总体上,株高与基径的关系显著,除低海拔-全光照的lgH-lgD 表现为收益递减外,其余5 种生境的lgH-lgD 均表现收益递增,与树木[44]和竹类[37]相似。lgS-lgH 总体表现为收益递减。但同一遮阴条件下lgH-lgD、lgS-lgH 的斜率则大致相当,这说明生境对性状间的关系具有明显的塑造作用[27]。
综上,高海拔林下有利于缺苞箭竹长得更为高大。这与在白水江缺苞箭竹实际分布区的调查[3]和未来全球变化下分布区的模拟[26]相一致,即缺苞箭竹适应于高海拔地区的生境。但了解生境对缺苞箭竹生长的影响需要更多、更系统地性状研究,以为大熊猫国家公园竹类的定向管理提供充足的科学依据。
