色季拉山川滇高山栎种群结构对海拔梯度的响应
2023-11-13田莹卢杰王新靓杨琳
田莹,卢杰*,王新靓,杨琳
1.西藏农牧学院高原生态研究所,西藏 林芝 860000;2.西藏高原森林生态教育部重点实验室,西藏 林芝 860000;3.西藏林芝高山森林生态系统国家野外科学观测研究站,西藏 林芝 860000;4.西藏自治区高寒植被生态安全重点实验室,西藏 林芝 860000
植物种群结构不仅可以反映植物在不同时间阶段的生长情况,还可以反映植物在不同生境下的生长变化情况[1]。种群结构主要从高度结构、径级结构、冠幅结构等方面开展研究,可以反映出种群数量个体大小的配置分布情况也可以以此为依据推测植物种群未来的发展趋势[2]。研究植物的种群结构对于了解植物的生存现状以及对于资源利用和保护具有重要意义[3]。
川滇高山栎(Quercus aquifolioides)作为我国特有植物,主要分布在西藏、四川、云南、贵州等地,生长在海拔2000 m-4 500 m 的山坡向阳处或高山松林下,对于高海拔地区的水源涵养、水土保持、生物多样性保护以及生态系统的维护具有重要作用[4,5]。目前对川滇高山栎的研究主要集中在滇西北玉龙雪山[6]、川西折多山[7]以及西藏色季拉山[5]地区,研究内容涉及叶片的结构特征[8]、遗传多样性[6]等生理特性以及川滇高山栎基因克隆[9]、光合作用[10]、繁育技术研究[11]等方面;关于海拔梯度上的研究有不同海拔梯度上的光合特性的响应[12]、叶片营养元素的变化特征[13]、叶片的解剖结构特征[14]、生理生长特性[15]、遗传多样性分析[16]等,而对于色季拉山川滇高山栎在不同海拔梯度上种群结构特征的相关研究相对较少。本文综合分析色季拉山川滇高山栎在不同海拔梯度上的种群结构变化,增加对高海拔地区生态系统的认识以及高寒环境下高山植物对生境的适应[17]。本研究以西藏色季拉山川滇高山栎种群为研究对象,对其在不同海拔梯度上的种群结构变化规律进行探索研究,旨在为青藏高原地区的生态系统的维持和保护提供一定的数据理论支撑。
1 研究区概况
色季拉山位于西藏自治区林芝地区林芝县以东(94°43′24″~94°43′1″E,29°44′14″~29°44′15″N),属念青唐古拉山向南延伸的余脉,是尼洋河流域与帕隆藏布江的分水岭,由于印度洋水汽在色季拉山两边的分布量不同,两条流域的降水量等有着较大差异,受印度洋的西南季风以及雅鲁藏布江暖湿气流的影响色季拉山干湿季分明,每年6—9 月份为雨季,降水量高,占全年的80%,年均降水量为1 134 mm,年均蒸发量544 mm,最冷月的平均气温为0.0℃~2.8℃,最低气温在-14℃,最热月的平均气温为11.5℃~18.2℃,最高温度在11℃,无霜期180 d,土壤类型为山地棕壤[18,19]。该区域乔木层主要有急尖长苞冷杉(Abies georgeivar.smithii)、高山松(Pinus densata)、西藏红杉(Larix griffithii)、林芝云杉(Picea likiangensisvar.linzhiensis),林下灌木层主要有雪层杜鹃(Rhododendron nivale),雪山杜鹃(R.aganniphum)、林芝杜鹃(R.nyingchiense)、方枝柏(Juniperus saltuaria)、西南花楸(Sorbus rehderiana)、长尾槭(Acer caudatu)和腰果小檗(Berberis johannis)等植物。
2 研究方法
2.1 样带设置
在色季拉山海拔3 400 m~3 780 m 之间设置一条由低海拔向高海拔延伸的50 m×700 m 的样带,样带的坡度25~35°,采用相邻格子样方法将样带分成14 个50 m×50 m 的样方,每一个样方四周用铁丝围起。在小样方中进行调查,利用测高仪和卷尺测量记录样方中所有高度>1.5 m 乔木的树高、胸径、冠幅、坐标,同时利用卷尺测量川滇高山栎幼树幼苗的高度、胸径、冠幅。海拔3 400 m~3 526 m 之间区域为低海拔区域,海拔3 526 m~3 652 m 之间区域为中海拔区域,海拔3 652 m~3 780 m 之间区域为高海拔区域。
2.2 种群结构划分
根据川滇高山栎的生存环境、生活史特征和调查的数据,本文在参考卢杰[20,21]等和谢立红等[22]研究方法的基础上,利用空间代替时间的方法,即用立木径级结构分析川滇高山栎的种群结构。将川滇高山栎胸(地)径划分为XI 级、高度划分为X 级、冠幅划分为Ⅷ级,川滇高山栎的冠幅以圆的面积进行计算。具体划分指标如表1 所示。统计各径级、高度级、冠幅级每一级别在每一样方内的数量,分别绘制分布结构图,分析种群结构。
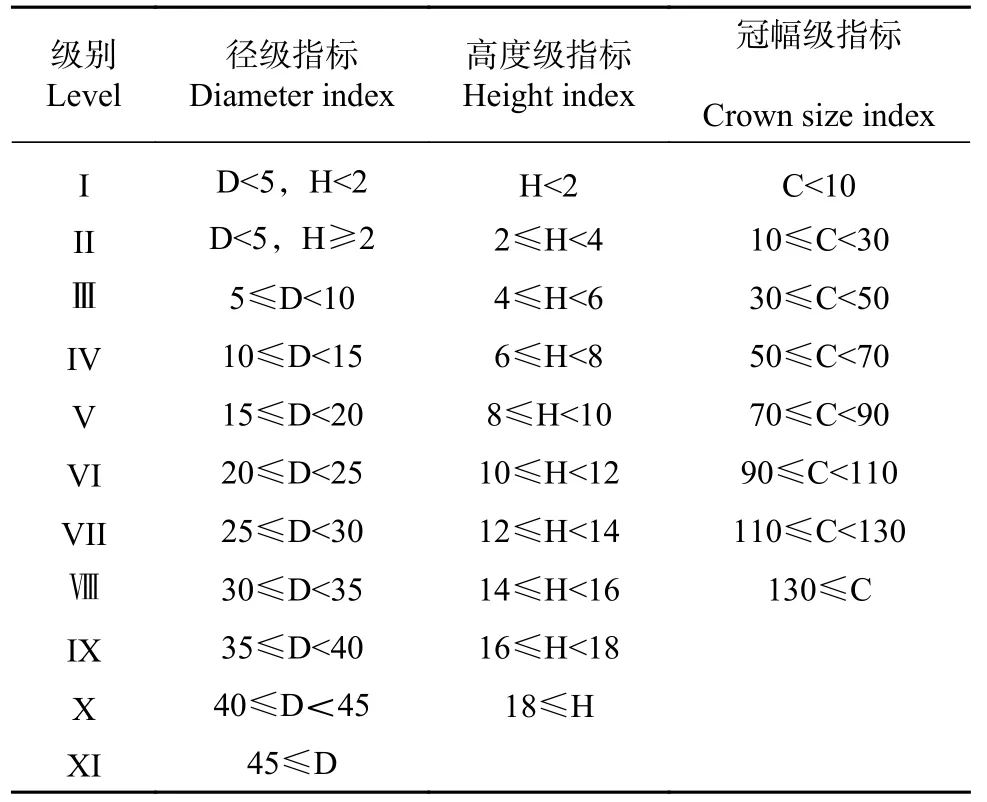
表1 川滇高山栎种群结构级划分Tab.1 Structural division level of Q.aquifolioides population
2.3 数据处理
采用Excel2016 软件将川滇高山栎的径级、高度级、冠幅级在每一样方内的分布数量统计出来并相应地做出每一级别的分布结构图。
3 结果与分析
3.1 种群结构特征
样带内川滇高山栎种群共1874 株,在海拔3 400 m~3 780 m 连续不间断的设置14 个50×50 m 的样方,样方号为横坐标,个体数为纵坐标,个体数随径级、高度级、冠幅级的分布如图1 所示,其中,DⅠ是幼苗和幼树,其个体数占总数的4.32%,株数为81 株;DⅡ是小树,个体数占12.32%,株数为231 株;川滇高山栎种群内幼苗幼树的数量很多,但其存活率较低,这可能是由于幼苗幼树自身对于环境的适应能力较强,在所处的环境中可以有较强的竞争力与其他物种竞争;DIII—DVIII 的个体数量由低径级向高径级呈现降低的趋势,此阶段的个体数占总数的75.5%,株数为1 415 株;DIX—DXI 的个体数较少,仅20 株,此阶段的个体数占总数的1.06%,最后两个径级个体数仅10 株,这可能是由于此阶段个体的竞争力和适应能力都比较低,单株胸径最大达到62.10 cm,胸径超过40 cm 的川滇高山栎有10 株。总体上,种群的个体数量随着胸径的增加而急剧减少,具体表现为小树、中树的数量丰富,但幼苗幼树向小树过渡的过程中树木个体大量死亡,种群的更新能力收到制约,种群的年龄结构整体属于增长性,但增长幅度较低。
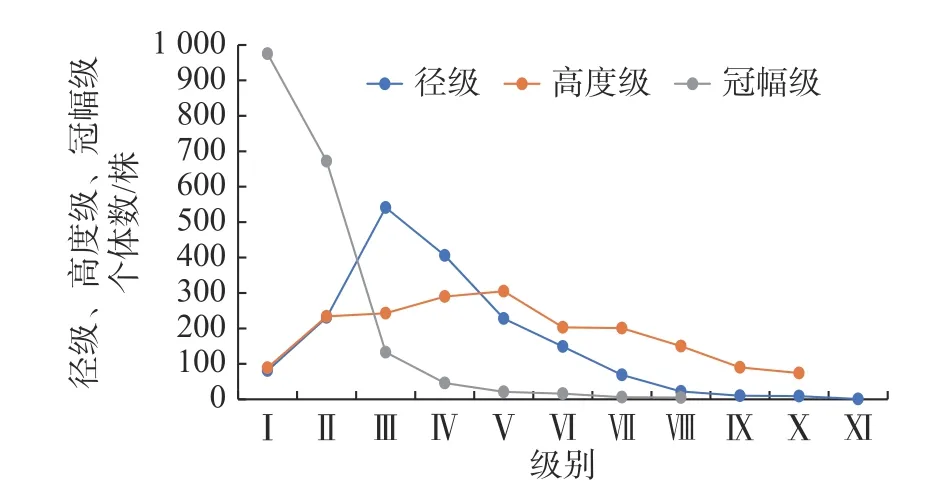
图1 川滇高山栎种群径级、高度级、冠幅级结构Fig.1 Diameter-,height-and crown-level structure of Q.aquifolioides population
从图1 中可以看出:HI 的个体数较少,占总数的4.32%,株数为89 株;HII 川滇高山栎数量迅速上升到234 株,占总体的12.45%;HIII—HV 级的个体数平缓上升,共838 株,HV 的个体数最多,占总数的16.27%,株数为305 株;HVI—HIX 的个体数平缓下降,共644 株;HX 的个体数最少,只有74 株,单株最高可达80 m,高度超过20 m 的川滇高山栎有22 株。随着高度的增加,川滇高山栎种群的个体数呈现先增加后减少的趋势,总体来看,川滇高山栎种群的高度结构比较完整,10 个高度级均有个体分布。
由于川滇高山栎的幼苗幼树的个体较小,暂时未形成方便统计的冠幅,故在统计冠幅级时除去56 株幼苗幼树,剩余川滇高山栎个体数为1818 株,其冠幅级结构如图1 所示。可以看出:CI 的个体数较多,共919 株,占总数的50.55%;CI—CVIII 均呈现下降的表现,CII 到CIV 的个体数下降幅度较大,CVIII 的个体数最少,仅5 株,占总数的0.27%。总体上看,川滇高山栎个体数随着冠幅级的增大而减小。
3.2 幼苗幼树对海拔梯度的响应
DⅠ和HI 分别对应径级和高度级的幼苗幼树,由图2 和图3 可知,幼树幼苗在每一个样方内都有分布,总体分布情况是低海拔区域个体数量较多,中海拔区域个体数量减少,高海拔区域个体数量增加,幼苗幼树径级和高度级的分布方式基本上是相似的。
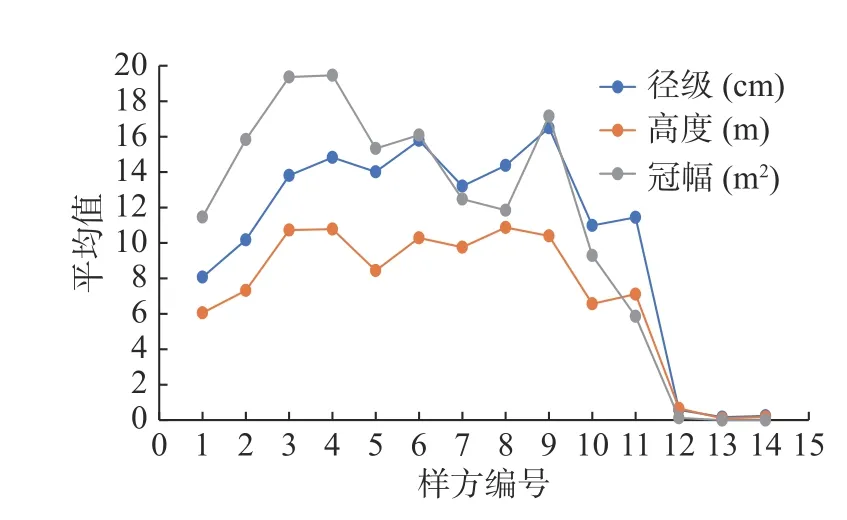
图2 川滇高山栎种群径级、高度级、冠幅级变化图Fig.2 Changes of diameter class,height class and crown class of Q.aquifolioides population
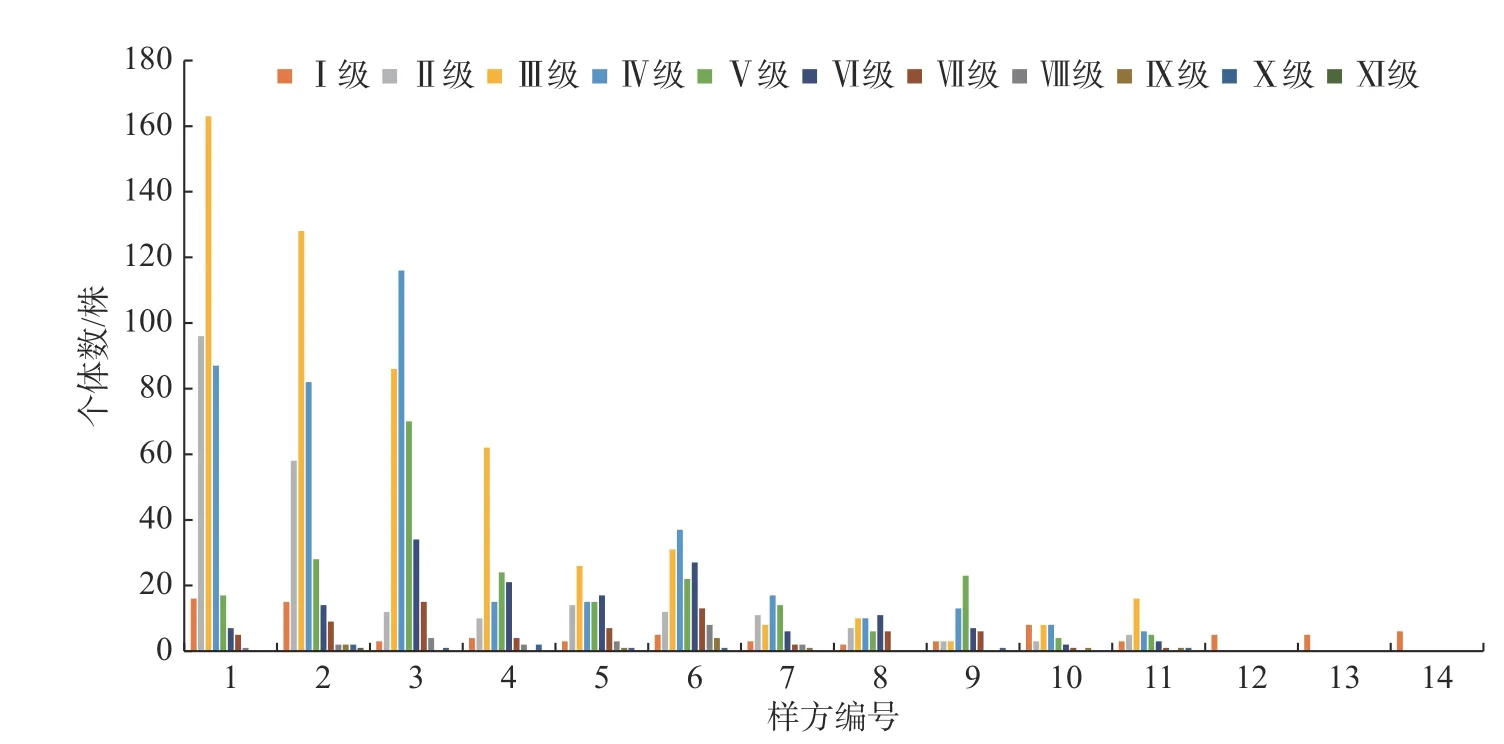
图3 川滇高山栎种群径级结构分布图Fig.3 Distribution map of the diameter class structure of Q.aquifolioides population
3.3 径级结构对海拔的响应
由图3 可知,川滇高山栎种群的个体数量呈现先增后减的趋势,只有径级结构DI 的川滇高山栎在每一个样方中都有分布,其他径级的川滇高山栎从第12 号样方起直到14 号样方都没有分布。DII 和DIII 的个体数量大体上呈现从海拔3 400 m 到3 780 m逐渐减少的趋势;DIV 到DVIII 的个体数量由海拔3 400 m 到3 780 m 呈现增加→减少→增加→减少的变化动态,大致上个体数量还是呈现减少的趋势;DIX 和DX 在一号样方里都没有个体分布,在由海拔3 400 m 到3 780 m 的分布中呈现断层的情况,DIX 在6 号样方里的数量最多,DX 在2 号样方和4 号样方里的数量最多,总体上个体数量还是呈现减少的趋势;DXI 只在2 号样方里存在,数量较少,只有1 株。如图2 所示,川滇高山栎径级的大小随着海拔的升高呈现先增加后降低的变化趋势,在中海拔区域径级数值最大,随着海拔的继续升高,数值迅速减小。
3.4 高度结构对海拔的响应
如图4 所示,HI 的川滇高山栎在1—14 号样方里都有分布,HII 到HX 的川滇高山栎基本上在海拔3 700 m 以上的第12 号样方至14 号样方都没有个体分布,或有极少的个体数分布在第12 号样方。HI 的川滇高山栎在海拔3 400 m 地区的第1 号和第2 号样方的个体分布数量最多,到第3 号样方个体数量急速减少,第3 号样方至第13 号样方平均分布数量有3.9 株,到3 700 m 的第14 号样方个体数量有明显增加;HII—HIV 的个体数量可以明显看出呈现降低的趋势,从海拔3 400 m~3 780 m 有明显的数量降低的变化;HV—HX 的个体数量有明显的增加→降低→增加→降低的变化规律。在低海拔区域个体数量少,低海拔向中海拔的过渡区域个体数量明显增加,中海拔区域的个体数量又有明显地减少,中海拔到高海拔的过渡区域个体数量又有明显的增加,直到高海拔区域基本上没有个体的分布。如图2 所示,川滇高山栎高度的大小随着海拔的升高而升高,在中海拔区域时高度达到最大值,随着海拔的继续升高,高度值逐渐下降。
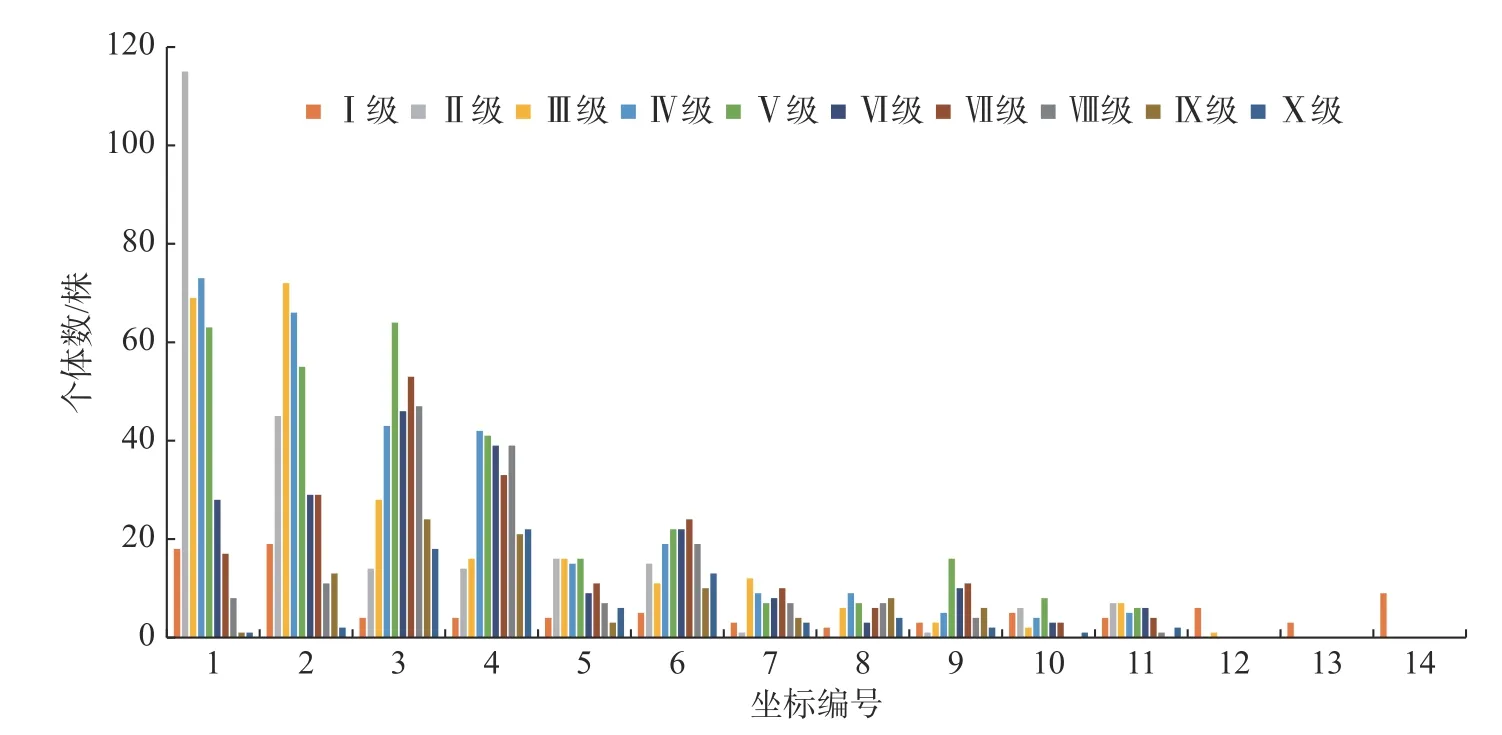
图4 川滇高山栎种群高度结构分布图Fig.4 Distribution map of height structure of Q.aquifolioides population
3.5 冠幅结构对海拔的响应
如图5 可以看出,CI 的川滇高山栎种群在3 400 m到3 780 m 的每一个样方里都有个体分布,个体数量从海拔3 400 m~3 780 m 呈降低的趋势;CII—CV 的个体数量有明显的先增加后降低的变化动态,到高海拔区域就没有川滇高山栎种群个体分布;CVI—CVIII 只有低海拔区域的三四个样方有个体分布,且分布数量较少,共有25 株,占总数的1.48%。冠幅级越小,个体数量越多;冠幅级越大,个体数量越少。高冠幅级的个体主要分布在低海拔区域。如图2 所示,川滇高山栎冠幅的大小随着海拔的升高同样呈现上升的趋势,当到达低海拔和中海拔之间的区域时达到最大值,随着海拔的继续升高,冠幅值逐渐减小,当到达中海拔和高海拔之间的区域时,冠幅值迅速上升,随着海拔的持续升高,冠幅值逐渐减小。
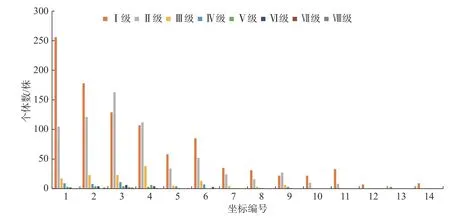
图5 川滇高山栎种群冠幅结构分布图Fig.5 Distribution map of canopy structure of Q.aquifolioides population
4 讨论
海拔差异会导致光照、土壤、降水和温度等环境因子产生变化,造成部分环境因子在小空间范围内进行重新配置,植物种群会根据环境因子的变化进行自身的种群个体分配[23,24],总体来说,川滇高山栎在由海拔3 400 m 到海拔3780 m 的成长过程中个体数量呈现先增加后降低的变化趋势,然而不同的种群结构对于海拔的变化呈现不同的变化动态。在卧龙巴朗山的川滇高山栎研究中,虽然在巴朗山的各个海拔梯度中,草本植物种数是占的比例最高,乔木植物种数是占的比例最低,但随着海拔的跨度变化,草本植物、灌木植物及乔木植物都会有不同的占地比例变化,随着海拔的升高,草本植物占地比例逐渐上升,乔木植物的占地比例逐渐降低,在海拔4 600 m 以上区域乔木植物的占地比例已经降为0%,这与色季拉山地区的川滇高山栎变化基本相似[25]。
4.1 海拔对川滇高山栎径级结构的影响
海拔梯度的变化会导致植物种群径级结构的变化,不同的海拔梯度植物种群的径级结构不同[26],在低海拔区域,径级各级的川滇高山栎个体数基本上都有分布,在高海拔区域,基本上只有DI 的个体有分布;在低海拔区域,小树的数量较多,这一区域温度适宜,土壤湿润,水分充足,生存环境适宜,幼树幼苗可以顺利生长到小树,在中海拔区域的川滇高山栎的种子通过动物或水流把种子运输到低海拔区域,这也是导致低海拔区域幼树幼苗数量多的因素;中海拔区域,中树和大树的数量较多,在此区域的树木分布相对于低海拔区域较为分散,给了川滇高山栎充足横向生长的空间;在高海拔区域基本上只有幼树幼苗的分布,是中海拔区域的川滇高山栎的种子通过动物或风力的干预将种子运输到高海拔区域,幼树幼苗的生存能力较强,小树自身的生存能力无法适应高海拔的恶劣环境,低温、降水量大且没有树木的保护导致幼树无法生长到小树。虽然不同海拔梯度各径级的个体数量差异较大,但总体上个体数集中于中小径级,有充足的中小个体作为生长资源,此区域的川滇高山栎种群为增长型的年龄结构。程子敏[27]等研究冀北山地的山杏在不同海拔的种群结构时,同样有着海拔梯度不同,山杏种群的径级结构有所变化,在海拔越高时,山杏种群的径级分布范围越小,种群密度也越小,和川滇高山栎种群的变化趋势相似。江蓝[28]在研究戴云山黄山松种群的径级结构对海拔的响应时,其研究结果与本次川滇高山栎的研究结果同样有着相同之处,总体都表现为中低海拔以中径级、大径级的个体为主,高海拔以小径级的个体为主。但川滇高山栎从幼树幼苗向小树增长的时期时,有大量的幼苗幼树无法正常生长到下一阶段,海拔梯度影响川滇高山栎的生长,从而影响其种群数量[29]。
4.2 海拔对川滇高山栎高度结构的影响
在衡量植物种群生长状况及立地情况级时,树高是一个重要的衡量指标,川滇高山栎因海拔位置、立地条件不同,立木高度存在一定的差异[30]。在低海拔区域,川滇高山栎的高度结构集中在HI—HV,高大树木分布密集,缺少充足的空间,在此区域内川滇高山栎难以进一步生长,无法与其他树木竞争;在中海拔区域,树高集中在HVI—HX,高大树木分布相对于低海拔区域较为分散,具有较为充足的空间为川滇高山栎的竖向生长提供支撑,在中海拔区域,既有树木为川滇高山栎的生长提供保护支撑,又有充足的空间支持川滇高山栎的生长;在高海拔区域,树高集中在HI,这一区域的高大树木分布极为分散,无法为幼树幼苗的生长提供保护,加上温度降低、光照强烈以及降水过多,幼树无法顺利生长到小树。从此可以得出,随着海拔的升高,树高呈现先增加后下降的趋势,到高海拔区域几乎只有幼树和幼苗,中海拔区域的中树大树较多,幼树幼苗生长成树的概率较大,这一区域相对来说适合川滇高山栎的生长。马剑[31]等在研究祁连山青海云杉树高在不同海拔梯度的变化规律时得出云杉树高的变化规律是由低海拔向高海拔呈先上升后下降,从中得出热量条件是限制云杉生长的关键。刘兴良[32]等在研究巴郎山川滇高山栎的高度结构对海拔梯度的响应时,巴朗山的川滇高山栎的高度结构随着海拔的升高呈逐渐下降的趋势,在海拔升高的过程中,川滇高山栎的高度结构时而出现不规则的升高。海拔过高或过低都不适宜川滇高山栎的生长,不同海拔梯度的热量、降水、光照以及生存空间都会影响乔木的高度的变化。
4.3 海拔对川滇高山栎冠幅结构的影响
植物群落与环境的相互作用影响着植物的冠幅结构生长,随着海拔的升高,川滇高山栎在不同海拔梯度区域的冠幅大小有着明显差异[33],在低海拔区域,川滇高山栎的冠幅结构CI—CⅧ都有分布,CI—CIV 的个体数量较多,在低海拔区域高大树木以及各种其他植物分布较密集,没有充足的空间提供给川滇高山栎冠幅的生长,光照、水源的不足以及严寒空气都限制了川滇高山栎的冠幅生长;在中海拔区域,依旧是CI—CⅧ的川滇高山栎都有分布,但CI—CIV 比CV—CⅧ的个体数量较多,在中海拔区域有较为充足的空间以及各种影响因子支撑川滇高山栎的生长,种内之间的竞争导致川滇高山栎的冠幅无法都继续生长,只有少数的个体冠幅可以生长到较大数值;在高海拔区域基本上只有CI 的个体存在,虽然在高海拔区域有充足的空间支持川滇高山栎冠幅的生长,但由于环境因子的影响此区域的川滇高山栎幼树幼苗无法进一步生长为小树,所以只有幼树幼苗的冠幅。赵维俊[34]等人研究祁连山排露沟流域的青海云杉林在不同海拔梯度的生长特征时得出青海云杉的冠幅大小随着海拔的升高呈增加趋势,当达到一定的海拔时冠幅大小达到最大,随着海拔的继续增加,云杉的冠幅不断减小。陈甲瑞[35]等在研究藏东南高山松在不同海拔的生长关系时,得出高山松的冠幅随着海拔的升高呈先降低后升高的趋势,且在海拔2 800 m~3 100 m 达到数值的最低值,后随着海拔的升高冠幅的数值逐渐增加,冠幅的增加有助于扩大个体的光合作用,抵御寒冷空气。同样川滇高山栎的冠幅在中海拔区域达到最大,随着海拔的继续升高,冠幅呈现降低趋势,在高海拔区域只有幼树幼苗的个体存在,幼树幼苗无法进行下一阶段的生长。
