干扰方式对香蕉园入侵杂草群落种间关联的影响
2023-11-11只佳增周劲松赵丽娟刘学敏张建春
只佳增, 杜 浩, 周劲松, 赵丽娟, 刘学敏, 张建春
云南省红河热带农业科学研究所, 云南 河口 661300
植物种间联结性能够直观和有效地反映群落的结构、功能、动态以及种间的相互关联性,常用于测度植物种间关系(张央等,2022),种间联结性反映了群落种间的相互作用及物种与环境因子间的耦合关系,是维持群落多样性及生态系统功能的重要生态过程(吴焦焦等,2020)。物种的空间分布格局在一定程度可以反映种间的关联程度,当2个物种关联程度较高时,它们通常会出现在相同区域(Maetal.,2022)。植物群落的总体联结性体现种间总体关联性,可以用来表征群落演替的阶段,当总体联结性较低时,群落总体呈现负的关联性,群落处于演替前中期;当总体联结性较高时,群落处于演替稳定期(吴鑫磊等,2022)。种间关系具有明显的环境依赖性,这种依赖性在胁迫环境下可能更为复杂,并可能直接影响植物的生存。因此,从种间关联角度研究胁迫环境下植物功能性状的种间差异,有利于揭示种间关系在植物功能性状变异中的作用(程久菊等,2022; Amatetal.,2015)。
农田杂草发生普遍、危害严重,是困扰农业健康发展的一个重要因素,传统耕作方式中对杂草的干扰管理措施常用的有使用化学除草剂、人工挖除和割草机除草等,不同的耕作方式是造成农田生态系统的干扰程度差异的主要因素(闵安民等,2005)。干扰对杂草种子库的种类组成有一定的影响,强干扰有利于一年生杂草和外来物种生存,免耕或减少干扰有利于多年生杂草的发生(李秉华等,2017)。我国农田耕作制度长期以来强调“清耕除草”,人为活动的干扰较为频繁,杂草演替更迭较快,致使农田杂草外来种的入侵已成为十分普遍的现象(郝丽芬等,2022)。外来入侵杂草会因其强大的适应能力及进入到新环境后占有生态位等打破生态系统平衡,排斥原有物种,导致当地植物多样性下降(Chenetal.,2020)。然而,有些入侵杂草由于根系浅、自然建群能力强以及植株较为低矮的特性,在生物防治中具有较大的应用潜力(左玉环等,2019)。纪红等(2021)通过研究阔叶丰花草SpermacocealataAublet对荔枝园杂草群落多样性的影响发现,阔叶丰花草定植改变了杂草群落结构,降低了恶性杂草牛筋草Eleusineindica(L.) Gaertn.、马唐Digitariasanguinalis(L.) Scop.等的重要值, 有利于荔枝园杂草管理。目前,国内关于干扰对农田生态系统杂草种间关系影响的报道较少,而杂草的种间联结性能够更加直观和有效地反映群落的结构、动态以及种间的相互关系,通过研究入侵杂草群落种间关联格局,从种间关联角度预测种群竞争结局和群落演替的方向,对预测农田杂草群落演替,特别是预测不同耕作方式对农田杂草群落的演替及有效防治杂草具有重要指导意义(孟祥杰等,2021; 王虎琴等,2016; 轩诗壮等,2021; 张亚芬等,2022)。
香蕉Musaspp.是热带地区重要的经济作物,云南红河流域的香蕉面积和产量均占云南全省的80%,香蕉是该地区主要农业经济来源之一,高温高湿的香蕉园杂草生长旺盛,以禾本科和菊科杂草居多,这些杂草光合效率高,根系发达,营养生长能力强,能快速向生殖生长过渡,最终与作物争夺水分和养分(杜浩等,2020)。只佳增(2020)研究发现,云南山地香蕉园的杂草种类丰富,没有明显的单优势种,雨季和旱季的杂草在生态位宽度和重叠上有着显著的变化,雨季优势草以马唐、香附子CyperusrotundusL.等单子叶杂草为主,旱季优势草以白花鬼针草BidenspilosaL.、小飞蓬ErigeroncanadensisL.等阔叶杂草为主,入侵杂草以连片斑块状分布于香蕉园,演替成该地优势杂草。目前已开展了香蕉园杂草群落物种多样性及优势种群生态位的研究,但关于杂草种间关系的研究还未见报道,鉴于此,本文拟通过研究不同人为干扰(使用除草剂、人工挖除、机械割除)方式下香蕉园入侵杂草的种间关联格局,阐明不同干扰方式对香蕉农田生态系统的物种影响,预测入侵杂草群落的演替方向及稳定机制,为有效进行杂草生物防治提供理论参考。
1 材料与方法
1.1 研究区域概况
试验地位于云南省河口县云南省红河热带农业科学研究所基地内(22°34′27″N, 103°52′26″E),海拔98~176 m,属低山河谷丘陵地貌,为热带季风雨林温热型气候,年平均温度22.6 ℃,年平均降雨量1587.3 mm,降雨长达163 d,为高温高湿气候。香蕉园土壤为红河流域冲积的酸性黄色砂壤土,供试土壤理化性质pH值4.83、有机质11.09 g·kg-1、全氮0.71 g·kg-1、速效磷28.60 mg·kg-1、速效钾256.0 mg·kg-1(只佳增等,2020),是香蕉栽培较为理想的种植地,试验地栽培的香蕉为自主选育品种及巴西蕉,均为中秆品种。
1.2 研究方法
试验于2018—2021年进行,设置3个杂草管理方式(人工挖除、药剂除草、机械割除),每个除草方式3个重复,平均每种除草方式的样地为3 hm2,总面积为9 hm2。其中,人工挖除频率为每2个月1次,把杂草全部(包括部分土壤种子)铲除;药剂除草频率为每4个月1次,喷洒除草剂(32%滴酸·草甘膦,用量为13.5 kg·hm-2)使杂草枯死;机械割除每2个月1次,只割除地上部分,根部以下还存活,所有样地其他生产方式相同。每个干扰方式随机设立9个10 m×10 m的固定样方(每个重复设3个样方),样方内采用 “M” 5点取样法(赵丽娅等,2021)设置取样点,每样点面积为1 m2,每个样地45个采样点,共135个采样点。每年冬春季、夏秋季各调查1次,共采集3年试验样地杂草种类、密度、多度和盖度等基本数据。
1.3 数据分析
1.3.1 重要值及优势度 本研究共调查到杂草97种,其中入侵杂草25种,占杂草总数的25.8%,计算所有杂草的重要值(important value,IV)、优势度(dominance of species,D),对主要入侵杂草群落、种间进行关联度分析(张金屯,2018)
D=相对多度+相对盖度+相对频度
(1)
IV/%=(相对多度+相对盖度+相对频度)×100/3
(2)
1.3.2 群落总体联结性 群落的总体联结性体现群落中种群间总的关联性,用Schluter提出的方差比率(variance ratio,VR)检验种间总体的关联性(赵丽娅等,2021)。
(3)
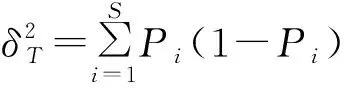
(4)
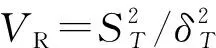
(5)

1.3.3 种间关联分析 为了排除物种分布的偶然性,只统计样方内出现5次以上的物种,进行种间关联分析,种间联结性基于建立2×2联列表,统计2个物种在样方内出现的次数,再用连续性Yates校正的χ2检验定性判定种间联结性(徐满厚等,2016)。
(6)
式中,N为总样方数;a为A和B物种同时出现的样方数;b为只有B物种出现的样方数;c为只有A物种出现的样方数;d为A和B物种都没有出现的样方数。若χ2≥6.635,表示种间联结性极显著(P≤0.01);3.841≤χ2<6.635,种间联结性显著(0.01 χ2不能定量区分联结程度,采用联结系数(association coefficient,Ac)进一步检验种间的关联程度(李娟和张昱,2022)。 (7) (8) (9) 联结系数Ac值域为[-1,1],Ac越趋近-1,表示2个物种间负联结性越强;Ac越趋近1,表示2个物种间正联结性越强;Ac=0,表示2个物种间完全独立。 为减小Ac联结系数受d值影响产生误差,采用Ochiai指数(IO)测定种对间的正联结程度(徐满厚等,2016)。 (10) 0≤IO≤1,IO值与a值密切相关,表示种对的联结性程度和相伴随机出现的概率,指数越大,种对同时出现在样方中的概率越大,种对间联结程度越趋向于正联结。 1.3.4 种间相关性 种间相关性用Spearman秩相关系数r(i,k)定量分析,其原理是利用物种优势度数据(温鑫鸿等,2022)。 (11) 式中,N为样方数,Xij和Xkj为物种i和物种k在样方j中的秩。 如表1所示,香蕉园入侵杂草绝大部分为来源于美洲的物种,不同人为干扰方式下入侵杂草的重要值有一定的差异,人工挖除、药剂除草、机械割除方式下入侵杂草重要值IV占比分别为35.16%、50.35%、52.91%,人工挖除方式对入侵杂草分布影响较大,重要值占比最低。在不同干扰下的白花鬼针草IV为12.22%~18.24%,阔叶丰花草IV为6.41%~12.68%,为绝对优势杂草,小飞蓬、野茼蒿Crassocephalumcrepidioides(Benth.) S. Moore、鹅肠菜Myosotonaquaticum(L.) Scop.为主要优势杂草。 表1 不同干扰方式香蕉园入侵杂草重要值Table 1 Values of invasive weeds in banana fields at different interference modes 表2 不同群落种间的总体关联性Table 2 Overall associations among species in different communities 2.3.1χ2检验 通过χ2检验可以看出(表3、4、5),人工挖除方式下入侵杂草间共组成55个种对,其中3对显著联结,1对极显著联结;药剂除草方式下共组成105个种对,其中3对显著联结;机械割除方式下共组成66个种对,其中6对显著联结,3对极显著联结。 表3 人工挖除方式下入侵杂草群落的χ2统计量检验Table 3 Invasive weeds communities χ2 statistical test of artificial weeding 表4 药剂除草方式下入侵杂草群落的χ2统计量检验Table 4 Invasive weeds communities χ2 statistical test of chemical weeding 表5 机械割除方式下入侵杂草群落的χ2统计量检验Table 5 Invasive weeds communities χ2 statistical test of mechanical weeding 2.3.2 种间联结系数 将Ac值划分5个区间,其中,Ac值在[-1,-0.5]说明种对间负联结性较高;值在(-0.5,-0.2]说明种对间有负联结性,但联结不紧密;Ac值在(-0.2,0.2)说明种对间联结性较弱,呈相互独立趋势;Ac值在[0.2,0.5)说明种对间有正联结性,但联结不紧密;Ac值在[0.5,1]说明种对间正联结性较高。由联结系数Ac值半矩阵图(图1)可知,人工挖除方式下,Ac值在(-0.2,0.2)的种对有43对(占总对数的78.2%),Ac值在[0.2,0.5)的种对有10对(18.2%),Ac值在[0.5,1]的种对有2对(3.6%);药剂除草方式下,Ac值在[-1,-0.5]的种对有19对(18.1%),Ac值在(-0.5,-0.2]的种对有17对(16.2%),Ac值在(-0.2,0.2)的种对有42对(40.0%),Ac值在[0.2,0.5)的种对有23对(21.9%),Ac值在[0.5,1]的种对有4对(3.8%);机械割除方式下,Ac值在[-1,-0.5]的种对有6对(9.1%),Ac值在(-0.5,-0.2]的种对有16对(24.2%),Ac值在(-0.2,0.2)的种对有35对(53.1%),Ac值在[0.2,0.5)的种对有7对(10.6%),Ac值在[0.5,1]的种对有2对(3.0%)。结果显示,人工挖除方式下入侵杂草间趋向于正联结,总体联结不紧密;药剂除草、机械割除方式下入侵杂草间负联结性强于正联结性。 图1 不同干扰方式下入侵杂草种间联结系数半矩阵图Fig.1 Semi matrix of interspecific association coefficient of invasive weeds at different interference modes 2.3.3 种间正联结程度IO值表示种对的正联结程度和共同出现的概率,将IO值划分3个区间,IO值为[0,0.5)说明种对间联结程度和共同出现的概率较低,IO值为[0.5,0.8)说明种对间联结程度和共同出现的概率较高,IO值为[0.8,1]说明种对间联结程度和共同出现的概率极高。由IO值半矩阵图(图2)可知,人工挖除方式下,IO值为[0,0.5)的种对有42对(占总对数的76.4%),IO值为[0.5,0.8)的种对有13对(23.6%);药剂除草方式下,IO值为[0,0.5)的种对有80对(76.2%),IO值为[0.5,0.8)的种对有25对(23.8%);机械割除方式下,IO值为[0,0.5)的种对有53对(80.3%),IO值为[0.5,0.8)的种对有12对(18.2%),IO值为[0.8,1]的种对有1对(1.5%)。结果表明,不同干扰方式下大部分入侵杂草种间正联结程度较弱,只有少部分种对间有较强的联结性,这与Ac值分析结果相似。 图2 不同干扰方式下入侵杂草种间正联结程度半矩阵图Fig.2 Semi matrix of interspecific interspecific association degree of invasive weeds at different interference modes 种间联结性是定性分析,而种间相关性是定量分析,后者具有更高的灵敏度(赵丽娅等,2021)。由秩相关分析(表6、7、8)可知,人工挖除方式下,有9个种对呈显著正相关,占16.4%,1个种对呈显著负相关,占1.8%;药剂除草方式下,有10个种对呈显著正相关,占9.5%,5个种对呈显著负相关,占4.8%;机械割除方式下,有6个种对呈显著正相关,占9.1%,3个种对呈显著负相关,占4.5%。在3种干扰方式下同时出现的28个种对中,有5个种对的相关显著性发生了改变。 表6 人工挖除方式下入侵杂草群落的秩相关系数半矩阵Table 6 Semi matrix of rank correlation coefficient of invasive weeds communities of artificial weeding 表7 药剂除草方式下入侵杂草群落的秩相关系数半矩阵Table 7 Semi matrix of rank correlation coefficient of invasive weeds communities of chemical weeding 表8 机械割除方式下入侵杂草群落的秩相关系数半矩阵Table 8 Semi matrix of rank correlation coefficient of invasive weeds communities of mechanical weeding 3种方式下重要值前三的入侵杂草都是白花鬼针草、阔叶丰花草、鹅肠菜,成为香蕉农田杂草的主要建群种。药剂除草和机械割除方式下入侵杂草优势度较人工挖除高,其中两耳草PaspalumconjugatumBerg.等禾本科杂草在人工挖除方式下的重要值较高,而飞机草Chromolaenaodorata(Linnaeus) R. M. King &H. Robinson、无刺含羞草Mimosadiplotrichavar.inermis(Adelb.) Verdc.等阔叶杂草在机械割除方式下有较高的重要值,这是由于干扰程度大的环境下以演替更新快的禾本科和一年生杂草为主(赵丽娅等,2021)。3种杂草管理方式群落种间总体呈显著正联结趋势,机械割除方式下入侵杂草总体种间关联性更强,群落更趋向稳定,这可能与机械割除方式只割除地上部分,根部以下还存活,未破坏土壤种子库,对环境压力较药剂除草和人工挖除小有关,可见,干扰程度是影响杂草群落分布格局的重要因素之一(张婧等,2017)。 种间关联性可以反映群落种间的相互作用关系,种间竞争与互利同时发生,并且二者的作用强度及平衡状态与环境条件密切相关(姜倪皓等,2022)。采用3种检验方法(χ2检验、联结系数Ac、Ochiai指数IO)分析各种对间的关联性得到结果基本一致,达到显著和极显著的种对较少,大部分入侵杂草种间联结性较弱,只有少部分种对间有较强的联结性。本研究中相同物种在3种干扰方式下,种间关联性质、关联程度有很大的差异,种间关联性在一定程度上反映了种间的相互关系及其与环境因子的关系,而种间的关系也一定程度上受环境因子的影响(何淑嫱等,2019; 赵丽娅等,2021)。研究显示,人工挖除方式下的群落中显著正联结的为两耳草等少数一年生杂草,而药剂除草和机械割除方式下显著正联结的以一年生阔叶杂草居多,速生型杂草间表现出一定的协同性,能够促进彼此的生长。在机械割除方式下,种间联结作用更显著,杂草群落呈正向演替趋势,推测该管理方式下干扰因素相对前二者较小,增强了气候因子及生境因子的效应,是导致种对间正负联结作用加强的主要原因(方楷等,2012)。此外,各检验方法存在一定的差异,如χ2检验是由物种在样方中是否出现得来,在一定程度上弱化了种间关联,不可避免地会损失信息量(崔玉华等,2021; 郭佳琦等,2021; 姜倪皓等,2022);在联结系数Ac检验中,当a=0时,Ac夸大了2个物种均不出现的联结性,导致负联结种对要多于正联结种对(吴艳玲等,2016)。阔叶丰花草在不同干扰方式下与种间都有显著的关联性,说明阔叶丰花草对环境的敏感性较低,具有极强的适应性,有在农田杂草群落稳定共存中发挥重要作用的潜力,这与纪红等(2021)的研究结果相似。种间关系在一定程度上取决于植物所处外界环境的压力强度,然而,本研究只探讨了干扰方式对入侵杂草种间关联的影响,干扰对种间关系的影响应综合考虑温度、光照、水分和土壤等环境因子的变化(赵丽娅等,2021),还有待进一步研究。 Spearman秩相关分析中,人工挖除、药剂除草、机械割除3种干扰方式下的入侵杂草显著相关的种对比例均较低,分别为18.2%、14.3 %、13.6 %,其中绝对优势入侵杂草白花鬼针草与阔叶丰花草之间也没有显著相关性,说明二者的种间关系比较松散,优势入侵种间的独立性较强,这与农田生态系统人为干扰程度大,先锋植物占有各自优势的生态位,加之农田杂草演替更新快,处于演替初期的群落种间的关联性较弱,甚至表现出较强的负关联(姜倪皓等,2022; 申旭芳等,2021)。此外取样尺度及取样量也会影响种间关系的分析结果,在本研究取样过程中,不同群落内种对间的正负联结数相差不大,正负联结种对比例趋近于1,说明本研究取样尺度合理有效(何光熊等,2023)。 综上所述,不同干扰方式下的香蕉园入侵杂草总体表现出正关联,但是大部分入侵杂草种间联结性较弱,只有少数种间有较强的联结性,种间关系表现较为松散。机械割除方式下的杂草群落更趋于正向演替,可见,适度的减小干扰程度有利于群落稳定。阔叶丰花草在杂草群落稳定共存中发挥重要作用,在香蕉园杂草生物防治中具有利用潜力。


2 结果与分析
2.1 主要入侵杂草群落物种概况

2.2 不同群落总体联结性分析


2.3 种间联结性分析




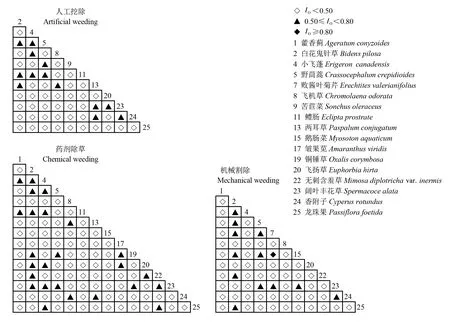
2.4 种间相关性分析



3 讨论
3.1 不同干扰方式下香蕉园入侵杂草组成及群落总体关联情况
3.2 不同干扰方式下香蕉园入侵杂草的种间关联格局
