取食臭椿不同部位对沟眶象和臭椿沟眶象成虫消化解毒酶的影响
2023-11-09马天驰温俊宝
马天驰,温俊宝
(北京林业大学林木有害生物防治北京市重点实验室,北京 100083)
沟眶象Eucryptorrhynchusscrobiculatus和臭椿沟眶象E.brandti同属鞘翅目象甲科沟眶象属EucryptorrhynchusHeller(赵养昌和陈元清,1980)。过去十几年,两种象甲严重危害宁夏园林绿化及农田防护林主要造林树种臭椿。国内目前主要是把沟眶象和臭椿沟眶象作为一种钻蛀性害虫来研究(胡婷和张蕊,2019;王谦,2019;张志伟等,2019)。在国外(主要是美国和欧洲),臭椿被认为是外来入侵种(Espenschied-Reillyetal.,2008; Terzietal.,2021),相关研究集中在把这两种象甲作为天敌昆虫来防治臭椿,而臭椿沟眶象是研究和利用的重点(Kowariketal.,2008;Herricketal.,2012)。
沟眶象幼虫取食臭椿树根,臭椿沟眶象幼虫取食树干木质部和韧皮部;臭椿沟眶象成虫主要取食臭椿主干,而沟眶象成虫主要取食臭椿的叶柄、1年生枝以及臭椿的其他幼嫩部位(Jietal.,2017)。国内外尚未对两种象虫共存于臭椿上的机制进行研究。
象甲成虫大多有补充营养习性,通常被认为对成虫产卵量和寿命有显著影响(Saitoetal.,2005;高旭等,2009;张锋,2013)。臭椿沟眶象成虫和沟眶象成虫在羽化后同样要靠取食臭椿补充营养来达到性成熟。补充营养同样会对沟眶象的产卵量和卵孵化率产生显著影响,取食臭椿的沟眶象产卵量和卵的孵化率要显著高于取食非寄主(清水、蜂蜜和苹果)的沟眶象成虫,而靠清水补充营养雌虫产的卵不能正常孵化。因此,可以认为补充营养会对沟眶象雌性成虫的生殖系统发育产生显著影响,营养来源关系到成虫的产卵量和卵的发育(汪永俊等,1984;高旭等,2009;张艳,2015)。
前人采用臭椿不同部位(叶柄、1年生枝、2~3年生枝和主干)对沟眶象和臭椿沟眶象进行补充营养,两种象甲的卵巢发育程度和产卵量都有明显差异。Guoetal.(2019)研究取食不同部位臭椿对臭椿沟眶象的卵巢发育程度和产卵影响,发现取食叶柄、1年生枝、2~3年生枝的臭椿沟眶象雌虫缺少功能性卵巢,无法产卵,而取食主干的雌虫会正常发育出功能性卵巢并成功产卵。沟眶象取食臭椿不同部位后,用叶柄饲养的卵巢发育程度最好,其次是主干,然后是1年生枝,而取食2~3年生枝的沟眶象不能完成卵巢发育,且无法产卵(郭文娟,未发表)。
植物的挥发物、物理性质、次生代谢产物和营养物质等外部因素,昆虫肠道微生物种类和数量,以及昆虫消化解毒酶活性等内部因素都对昆虫取食行为具有重要影响。用HPLC法和光谱法检测臭椿不同部位的次生代谢物质如黄酮类物质、没食子酸、β-谷甾醇和单宁,营养物质如糖、蛋白质、粗脂肪、纤维素以及一些金属元素等,证实这些次生代谢物和营养物质在臭椿不同部位的确存在差异。推测这些次生代谢物和营养物质对沟眶象和臭椿沟眶象雌成虫生殖系统发育造成了不同的影响,而两种象甲中后肠的消化解毒酶也可能因此发生变化。
使用解毒酶系进行解毒代谢是昆虫应对寄主植物防御的一种重要方式(Feltonetal.,2008)。谷胱甘肽硫转移酶(GST)、羧酸酯酶(CarE)是和细胞色素P450(CytP450)是昆虫体内三大解毒酶(宋维虎,2020),此前对沟眶象和臭椿沟眶象的研究没有涉及过这3种解毒酶。因此选择这3种解毒酶为研究目标。
另一方面,营养物质吸收受损的生理成本通常与脂质、蛋白质和碳水化合物代谢速率的改变相关,从而导致食物转化为生物质的效率降低,如豌豆蚜Acyrthosiphonpisum的中肠消化酶活性和苜蓿MedicagoSativa抗性品种有关,苜蓿品种的抗性越强则豌豆蚜虫消化酶活性越弱(达丽婷等, 2015)。昆虫体内与脂质、蛋白质和碳水化合物有关的主要消化酶有纤维素酶、蛋白酶、脂肪酶、淀粉酶。前期已经对沟眶象相关纤维素酶做了研究(雍惠莉等,2007;杨贵军等,2008;陈雅媛,2016;任媛,2017),因此选择蛋白酶、脂肪酶和淀粉酶为3种主要的消化酶。
本研究选取臭椿4种不同部位(叶柄、1年生枝、2~3年生枝、主干)饲喂沟眶象和臭椿沟眶象成虫,并利用ELISA试剂盒测定两种象甲的3种解毒酶(谷胱甘肽转移酶、羧酸酯酶和细胞色素P450)和3种消化酶(胰蛋白酶、淀粉酶、脂肪酶)的活性,以探究取食臭椿不同部位对沟眶象和臭椿沟眶象成虫消化解毒酶的影响。
1 材料和方法
1.1 供试材料
臭椿沟眶象成虫的获得:2021年4月在宁夏灵武市梧桐树村(116°17′50.81″N,38°9′1.37″E)砍伐3棵约20年生受害臭椿,其中1棵没有发芽,2棵顶部有1年生新枝条;同期,在宁夏吴忠市巴浪湖(116°12′47.98″N,37°54′36.65″E)砍伐 3棵 30~40年生受害臭椿,其中2棵没有发芽,只有1棵顶部有当年新枝条;所有6棵受害臭椿上均有新鲜虫粪和流胶。将臭椿树锯成60~80 cm长的木段方便运输和培养,通过物流运回学校(116°20′49.63″N,40°0′34.01″E),在室内遮阳遮雨,自然温度湿度条件下用网罩罩住,每天检查 1次,收集当天羽化的臭椿沟眶象成虫,采用陈冲等(2013)的方法来区分臭椿沟眶象雌雄虫。
沟眶象成虫的获得:2021年5月24日在宁夏吴忠巴浪湖(116°12′47.98″N,37°54′36.65″E)采集了羽化不久的沟眶象成虫,雌雄各48头。
臭椿枝条的获得:每周从宁夏吴忠野外农田防护林中选择有臭椿沟眶象取食且未施用农药的臭椿,采集新鲜足量的不同部位(叶柄、1年生枝、2~3年生枝条、主干),快递寄回北京,放在冰箱中4℃冷藏,每2 d更换1次。臭椿不同部位的选择基于Zhangetal.(2017)的方法。
臭椿沟眶象成虫的饲养:臭椿沟眶象开始羽化后,当天羽化的雌/雄成虫每4头为1组,每4组分别放在有盖并打孔透气的塑料盒中用新鲜的臭椿不同部位(叶柄、1年生枝、2~3年生枝条、主干)进行饲养处理,每个处理6个生物学重复。臭椿沟眶象先饲养24 h,再饥饿处理24 h后解剖,如来不及解剖则先在超低温冰箱80 TSX(Thermo Fisher,America) 中-80℃冰冻。饲养光周期14 L∶10 D,温度25±1℃,相对湿度60%±5%。
沟眶象成虫的饲养:由于沟眶象在土中羽化,难以获得刚羽化成虫,而挖土采集老龄幼虫或蛹的难度太大,故尽量采集羽化出土不久的成虫进行试验。一般成虫背部红色鳞片范围越大,沟眶象成虫越接近刚羽化。试验用沟眶象成虫羽化后时间约0~5 d,雌/雄成虫每2头为1组,快递寄回北京林业大学森保研究室后每4组分别用新鲜臭椿不同部位(叶柄、1年生枝、2~3年生枝条、主干)在有盖并打孔透气的塑料盒中进行饲养处理。每个处理6个生物学重复。沟眶象先饲养24 h,再饥饿处理24 h后解剖,如果来不及解剖则在超低温冰箱中-80℃冰冻后解剖。饲养光周期为14 L∶10 D,温度25±1℃,相对湿度60%±5%。
1.2 试验方法
1.2.1酶活液的制取
沟眶象和臭椿沟眶象解剖时,对超净工作台进行紫外灭菌0.5 h,所有材料和仪器预先进行灭菌。分别用70%乙醇浸泡2 min,再用磷酸盐缓冲盐水PBS清洗两种象甲(Axelssonetal.,2017)。在超净工作台上先剪掉沟眶象和臭椿沟眶象所有足,用手分离其鞘翅,用眼科手术剪剪去其后翅,再在其背面胸腹连接处横着剪开,沿着其背部断口中缝剪至其肛门,在显微镜下观察,用3根昆虫针分别固定其腹部靠近腹中线的位置和体壁裂口近胸端左右侧,使其体壁完全展开,并固定在冰盒上的蜡板上,在体式显微镜Olympus 311477(Tokyo)下取出腹部的中后肠加入1.5 mL离心管中,在电子天平上称取中后肠重量并放置在PBS(cytiva,America)缓冲液中,其中中后肠质量∶PBS体积=1∶9质体比,再使用手持电动组织研磨枪OSE-Y40(TIANGEN,Beijing)匀浆后,在低温离心机Fresco 21(Thermo Fisher,America)中4℃低温离心16 000 r/min,取出上清液,将大管中溶液再分装5个1 mL离心管(生物学重复5次),和剩下的大管一起放在-80℃超低温冰箱中保存备用。
解剖后样品标记如下。EbFT:取食主干的臭椿沟眶象雌虫;EbMT:取食主干的臭椿沟眶象雄虫;EbFB:取食2~3年生枝条的臭椿沟眶象雌虫;EbMB:取食2~3年生枝条的臭椿沟眶象雄虫;EbFA:取食1年生枝条的臭椿沟眶象雌虫;EbMA:取食1年生枝条的臭椿沟眶象雄虫;EbFP:取食叶柄的臭椿沟眶象雌虫;EbMP:取食叶柄的臭椿沟眶象雄虫;EsFT:取食主干的沟眶象雌虫;EsFT:取食主干的沟眶象雄虫;EsFB:取食2~3年生枝条的沟眶象雌虫;EsMB:取食2~3年生枝条的沟眶象雄虫;EsFA:取食1年生枝条的沟眶象雌虫;EsMA:取食1年生枝条的沟眶象雄虫;EsFP:取食叶柄的沟眶象雌虫;EsMP:取食叶柄的沟眶象雄虫。
1.2.2酶活性测定
使用酶联生物的6种酶的ELISA试剂盒(昆虫谷胱甘肽转移酶(GST)ELISA试剂盒ml062705、昆虫羧酸酯酶(CES)ELISA试剂盒ml036265、昆虫细胞色素P450(CYP450)ELISA试剂盒ml036261、昆虫胰蛋白酶(trypsin)ELISA试剂盒ml036260、昆虫淀粉酶(AMY)ELISA试剂盒ml062684、昆虫脂肪酶(Lipase)ELISA试剂盒ml062686)进行酶活性测定。
按试剂盒说明书加样和检测,具体操作如下:①加样:设置标准品孔、样本孔和空白孔,标准品孔中加6种不同浓度的标准品,各设置5个生物学重复,技术重复3次。在待测样品孔中加样品稀释液40 μL与待测样品10 μL,空白孔之加入稀释液。分别在标准品孔和样本孔各自加入酶标试剂100 μL。②恒温培育:使用封板膜封好后置37℃恒温培育1 h,并将20倍洗涤液用双蒸水稀释后待用。③清洗酶标板:揭掉封板膜,弃去所有液体,待甩干后往每孔加满洗涤液,静置30 s后弃去,重复洗涤5次,拍干酶标板。④显色反应:每个孔先加入显色剂A 50 μL,再加入显色剂B 50 μL并震荡混匀,使用锡纸包住酶标板后,在37℃恒温条件下避光显色,15 min后加入反应终止液50 μL。⑤上机检测:在酶标仪SpectraMax 190(Molecular Devices,America)上测定酶活性,使用空白孔调零,在450 nm波长下测定酶标板的吸光度(OD值)。
1.3 数据分析
使用SPSS v26检查3个技术重复中所测定OD值是否存在可疑数值,剔除可疑值。使用标准物的浓度与OD值在Orgin 2021b软件中计算出标准回归曲线,再将样品的OD值代入方程式,计算出样品浓度。
所有酶的酶活性单位统一为U/L,数值用平均值 ± 标准差表示,不同小写字母表示其所在组的酶活性存在显著性差异。使用Orgin 2021b中的Paired comparison Plot v3.60 app作图,并采用单因子方差分析(ANOVA)以明确取食臭椿不同部位(叶柄、1年生枝、2~3年生枝、主干)对沟眶象和臭椿沟眶象成虫3种解毒酶(谷胱甘肽转移酶、羧酸酯酶和细胞色素P450)和3种消化酶(胰蛋白酶、淀粉酶、脂肪酶)活性的影响,处理间差异显著性分析采用Turkey检验(P< 0.05)。
2 结果与分析
2.1 取食臭椿不同部位对两种象甲成虫细胞色素P450活性的影响
臭椿沟眶象雌虫取食2~3年生枝条CytP450 酶活性最低,其次是取食主干,取食1年生枝组和叶柄组最高且无显著性差异;雄虫取食1年生枝CytP450 酶活性最低,其次是取食叶柄,取食主干较高,取食2~3年生枝条最高,1年生枝组和叶柄组,主干组和2~3年生枝组之间差异不显著。沟眶象雌虫取食4种不同部位后CytP450酶活性无显著性差异;雄虫取食叶柄CytP450酶活性最低,其次是取食主干组和2~3年生枝组,取食1年生枝组最高(图1)。
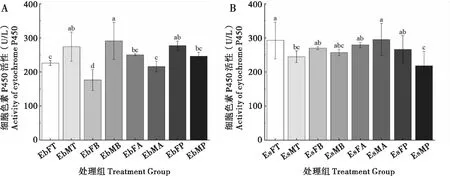
图1 两种象甲细胞色素P450酶活性
2.2 取食臭椿不同部位对两种象甲成虫谷胱甘肽转移酶活性的影响
臭椿沟眶象雌虫取食4种不同部位后GST酶活性无显著性差异;雄虫取食主干、1年生枝和2~3年生枝间GST酶活性较低,两者间差异不显著,取食叶柄最高。沟眶象取食叶柄雌虫GST酶活性最低,其次是取食2~3年,取食主干和1年生枝最高,但无显著差异;雄虫取食叶柄GST酶活性最低,其次是取食主干和2~3年生枝,取食1年生枝最高(图2)。
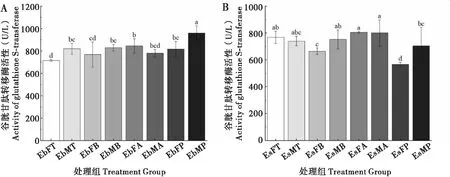
图2 两种象甲谷胱甘肽转移酶活性
2.3 取食臭椿不同部位对两种象甲成虫羧酸酯酶活性的影响
臭椿沟眶象雌虫取食主干和2~3年生枝CarE酶活性显著低于取食1年生枝和叶柄;雄虫取食2~3年生枝CarE酶活性最低,取食其它3个部位较高且无显著差异。沟眶象雌虫取食叶柄CarE酶活性最低,其次是取食主干,取食2~3年生枝和1年生枝组最高且无显著差异;雄虫取食叶柄CarE酶活性最低,其次是取食主干和2~3年生枝,取食1年生枝最高(图3)。
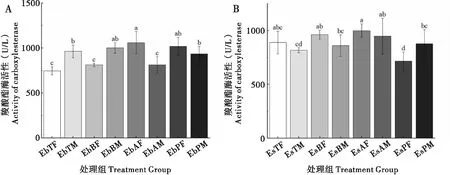
图3 两种象甲羧酸酯酶活性
2.4 取食臭椿不同部位对两种象甲成虫脂肪酶活性的影响
臭椿沟眶象雌虫取食1年生枝和主干LIP酶活性较低且无显著性差异,取食叶柄其次,取食主干最高;雄虫取食主干LIP酶活性最低,取食2~3年生枝其次,取食1年生枝和叶柄最高且无显著性差异。沟眶象雌虫取食叶柄LIP酶活性最低,取食2~3年生枝组较低,取食主干较高,取食1年生枝最高;雄虫取食叶柄和主干LIP酶活性最低且无显著差异,取食2~3年生枝较高,取食1年生枝最高(图4)。
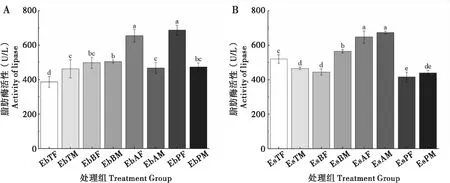
图4 两种象甲脂肪酶活性
2.5 取食臭椿不同部位对两种象甲成虫淀粉酶活性的影响
臭椿沟眶象雌虫取食主干和2~3年生枝AMY酶活性较低且差异不显著,取食1年生枝和叶柄较高且无显著性差异;雄虫1年生枝AMY酶活性最低,其次取食主干,取食2~3年生枝和叶柄最高且无显著性差异。沟眶象雌虫取食叶柄、主干和2~3年生枝AMY酶活性最低且无显著差异,取食1年生枝较高;雄虫取食叶柄和主干AMY酶活性最低且差异不显著,取食2~3年生枝和1年生枝最高,但两者差异不显著(图5)。
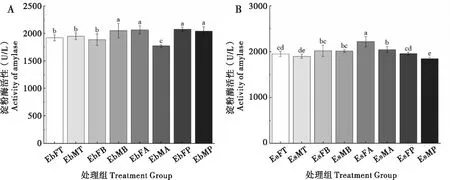
图5 两种象甲淀粉酶活性
2.6 取食臭椿不同部位对两种象甲成虫胰蛋白酶活性的影响
臭椿沟眶象雌虫取食主干和2~3年生枝TRP酶活性较低,无显著性差异;而取食1年生枝和叶柄较高,无明显地差异;雄虫取食1年生枝TRP酶活性最低,取食其他3个部位较高。沟眶象雌虫取食主干TRP酶活性最低,其次是取食2~3年生枝,而取食叶柄相对较高,取食1年生枝最高;雄虫取食4种不同部位后TRP酶活性无显著性差异(图6)。
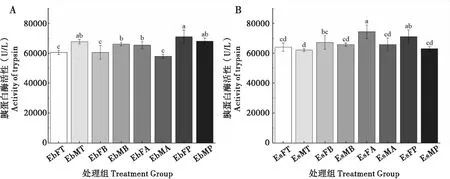
图6 两种象甲胰蛋白酶活性
3 结论与讨论
臭椿沟眶象雌虫取食主干后,解毒酶中CytP450酶活性较低,GST酶活性变化不显著,CarE酶活性最低,消化酶中LIP、AMY和TRP均为酶活性最低;而沟眶象雌虫取食2~3年生枝后,解毒酶中CytP450酶活性变化不显著,GST酶活性较低,CarE酶活性最高,消化酶中LIP和TRP酶活性较高,AMY活性最低。
考虑到Guoetal.(2019)的结论是只有取食主干的臭椿沟眶象雌成虫能正常产卵,以及只有取食2~3年生枝的沟眶象雌成虫不能正常产卵。如果两种象甲雌成虫取食臭椿不同部位(主干、2~3年生枝、1年生枝和叶柄)后的不能正常产卵和本研究中某一种消化酶或者解毒酶有关的话,则臭椿沟眶象取食主干组的雌成虫该种消化酶活性应该显著较高,或者该种解毒酶活性较低;而沟眶象取食2~3年生枝组的雌成虫该种消化酶活性应该显著较低,或该种解毒酶活性应该显著较高。因此臭椿沟眶象雌成虫只有取食主干能顺利产卵和取食主干CarE酶活性较低有关,沟眶象雌成虫只有取食2~3年生枝不能产卵和CarE酶活性较高以及AMY酶活性较低有关。
营养是影响昆虫雌虫交配和生殖的重要因素,成虫的补充营养行为会对雌虫的潜在生殖能力和实际的生殖能力起着重要作用(王竑晟,2003)。因为寄主植物的生长量以及营养质量能够影响昆虫的内分泌和生长,从而导致昆虫个体遗传差异以及昆虫种群间遗传分化(Geisert and Meinke, 2013)。由此可见,寄主不同部位的营养若存在差异,对于取食该寄主不同部位的昆虫,例如沟眶象和臭椿沟眶象这两个近缘种,可能使这两种象甲雌成虫补充营养后的生殖能力产生差异。
植物为了抵御植食性昆虫的取食,通过形态学以及化学分子调控等多种方式,对植食性昆虫产生防御机制(王亚军等,2017)。植物次生代谢物是通常不被昆虫作为营养物质利用,而一般起到植物防御昆虫取食的作用,影响昆虫对寄主的选择、利用和摄食(姜虹,2017;李明等,2017)。植物次生代谢物同样会影响昆虫的生长发育,如植物提取物槲皮素处理瓜实蝇Bactroceracucurbitae的卵和幼虫后,会抑制其卵的孵化以及幼虫的生长发育 (Sharma and Sohal, 2013)。昆虫使用化学感器识别和感受植物次生代谢物,在昆虫对寄主的选择中发挥了重要的作用(Pickettsetal., 1992;戴建青等,2010)。虽然没有直接证据表明同一寄主植物不同部位次生代谢物对昆虫成虫的生长发育会产生影响,但是有研究表明寄主植物的次生代谢物确实能对昆虫成虫的生长发育造成影响,臭椿不同位置的次生代谢物差异也可能是两种象甲野外取食分布差异的原因。
酶活性的变化能在短时间之内反应寄主昆虫体内的情况,通过对酶活性的分析有利于了解不同取食对寄主昆虫产生的影响。本研究也证明,即使是同一种植物,取食的部位不同仍然会对昆虫的消化解毒酶产生显著性的影响。
本研究明确了寄主植物不同部位对这两种象甲成虫中后肠消化解毒酶活性的影响,分析了两种象甲刚羽化雌成虫补充营养造成生殖系统发育差异的原因,对后续这两种象甲消化解毒酶相关研究有一定参考价值。
