镉胁迫下不同甘薯品种抗性分析
2023-11-09陈丰酆廖乐琴俞笛扬朱恩国陶亮奇王朝福凌腾芳康乐
陈丰酆,廖乐琴,俞笛扬,朱恩国,陶亮奇,王朝福,凌腾芳,康乐*
(1.西华师范大学 环境科学与工程学院,四川 南充 637000;2.韩国生命工学研究院,韩国 大田 34141)
金属镉是一种受到社会各界广泛关注的有毒重金属元素,过量摄入镉对人体和动植物体等都有着巨大的危害作用。人体研究表明镉对细胞内抗氧化酶活性具有抑制作用,促使细胞的氧化胁迫加剧,最终对人体的健康造成伤害[1-2]。镉进入人体的主要途径是通过食物链,长期食用含镉食物会使镉大量积存于肝或肾脏,对其造成危害,尤其是对肾脏危害最为明显,极少数严重的病人会出现骨骼病变,1931年日本出现的“骨痛病”就是由于过度摄入镉而产生。近些年在自然原因以及人为因素的双重影响下,部分地区耕作地内土壤中的镉含量逐渐增加,从而导致土壤毒性增加[3-5],使得其中的栽种的作物减产甚至绝收[6]。镉对植物的途径主要通过根系与叶片进入植物体内,可以破坏植物细胞内的线粒体与叶绿体结构,从而影响植物的光合作用和呼吸作用[7-8]。同时与人体研究结果类似的是高浓度镉也能抑制植物体内酶的合成,部分酶甚至会失活[8],与之同时植物细胞也会受到明显的氧化伤害,最终导致植物生长和发育受到抑制[9-10]。
甘薯又名红薯、番薯、地瓜,为旋花科植物,是中国四大粮食作物之一,在世界粮食地位中同样占有重要地位[11]。它具有丰富的营养价值和防癌作用,其体内丰富的胡萝卜素还能够有效地清除自由基,,而自由基是诱发癌症等疾病的原因之一[12-13]。Kumar等研究表明,在铬处理条件下,甘薯会通过增加体内POD、CAT、SOD等抗氧化酶活性来减少该种重金属对甘薯的伤害,在镍处理时,体内抗氧化酶活性也会增加,在高浓度镍的处理条件下,甘薯还可以通过减少氧化应激来减少镍对其伤害[14-15]。Lai等发现紫甘薯在受到铀(U)毒害时,减轻毒害的主要途径是通过改善自身能量供应和增加初级代谢物的合成,当U对红薯进行毒害时,会显著诱导甘薯防御相关基因的表达,包括抗氧化物质或酶等含量的增加来抵御重金属U的毒害[16]。
通常来说植物细胞内的活性氧是通过酶促系统(如SOD、CAT和POD等物质)和非酶促系统来进行清除的[17],它们相互协调,清除细胞体内多余的活性氧,从而减轻活性氧对植物的伤害[18-19]。但当植物长期处于重金属胁迫下,其体内的活性氧会大量增加[20],超过了植物本身的活性氧清除限度,使得植物细胞正常功能受到严重影响,最终引起作物衰老甚至死亡。因此,在重金属胁迫下,植物体内活性氧的清除能力可以影响到植物的抗重金属的胁迫的能力[21-22]。基于此,本研究以10种不同的甘薯植株材料探讨镉处理对其生理生化指标的影响,比较其体内叶绿素、H2O2、各种抗氧化酶含量的差异,揭示甘薯在镉胁迫下的调节机制,并进行甘薯品种的抗性分析。
1 材料与方法
1.1 实验材料
以南紫薯020、渝薯153、漯徐2号、冀薯4号、渝薯1号、南渝薯02、冀薯5号、淮薯3号、渝薯33、烟薯25为供试材料。将10种甘薯植株分别种入含镉苗床及其对照苗床,最后取其叶片作为实验材料。
1.2 实验方法
选取10种不同品种的甘薯植株,对照组用含改良霍格兰营养液的苗床进行水培培养,实验组则采用含450 μmol/L镉的改良霍格兰营养液进行培养,通过观察甘薯植株表现性状,当甘薯植株对照组与实验组出现明显差异时(如图1~2),将选取其植株第3片至第5片叶进行实验指标测定。

图1 CK处理
1.3 测量方法
叶绿素含量的测定方法选取乙醇提取-紫外分光光度法进行测定[23],H2O2、SOD、GSH、CAT、POD的测定使用来自中国苏州格瑞思生物科技有限公司的市售检测试剂盒,并使用紫外分光光度计测量吸光度,实验均重复测定3次。
2 结果与分析
2.1 镉处理对甘薯叶片内叶绿素含量的影响
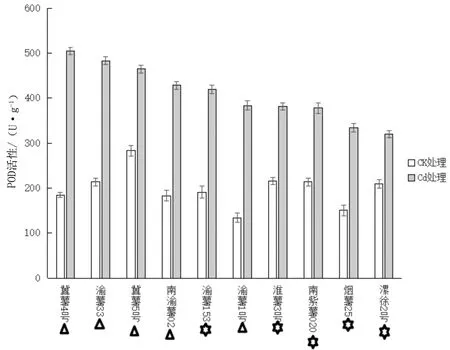
叶绿素是植物光合系统中重要组分之一, 其在叶片中的含量能直接影响到植物叶片光合作用的效率,进而影响到植物生长和发育。 通常测定叶绿素含量可以获得植物生长状态,因而长势良好的植物会含有更高含量的叶绿素。由图3可知,对照组漯徐2号、冀薯5号、南渝薯02、烟薯25、南紫薯020五类甘薯叶片叶绿素含量分别是实验组甘薯叶片叶绿素含量的2.06倍、2.21倍、2.09倍、2.83倍、2.5倍,差异达到极显著水平(P<0.01),对照组渝薯33、渝薯153、淮薯3号、冀薯4号、渝薯1号五种甘薯叶片叶绿素含量分别是实验组甘薯叶片叶绿素含量的1.49,1.29,1.46,1.86,1.79倍,差异达到显著水平(0.01 图2 Cd处理 表示镉胁迫组与对照组相比差异显著(0.01 H2O2是生物体内常见的活性氧之一,它能损伤和修饰生物大分子例如蛋白质分子和脱氧核糖核酸(DNA)。高浓度的过氧化氢能攻击这些分子产生细胞毒害。而低浓度的过氧化氢细胞内信号分子,在生物和非生物胁迫应激中以及植物生长发育中起重要的调节作用。由图4可见,南紫薯020、烟薯25、淮薯3号、漯徐2号、南渝薯02、漯徐2号、渝薯1号、渝薯33、冀薯4号、渝薯153甘薯叶片实验组H2O2含量分别是对照组甘薯叶片H2O2含量的5.33,5.2,5.67,4.72,5.22,3.91,4,4.68,4.23,4.27倍,差异达到极显著水平(P<0.01);镉处理条件下,各品种H2O2从少到多依次为渝薯153、冀薯4号、渝薯33、渝薯1号、冀薯5号、南渝薯02、漯徐2号、淮薯3号、烟薯25、南紫薯020。结果表明随着镉胁迫程度加深,甘薯叶片中H2O2含量会不断增加,使甘薯细胞受到毒害,从而抑制植株生长。 表示镉胁迫组与对照组相比差异极显著(P<0.01)。图4 镉处理对10种甘薯过氧化氢(H2O2)含量的影响 SOD是生物体内一种不可或缺的抗氧化酶,在生物体内普遍存在,主要负责清除·O2-发生歧化反应从而生成H2O2,它具有抗衰老和提高机体对疾病抵抗力等功能,还能帮助机体适应不同的外界环境,以减少植物体受到的氧化胁迫。由图5可知,冀薯4号、淮薯3号、烟薯25、南渝薯02、渝薯1号、漯徐2号六种甘薯叶片实验组SOD含量分别是对照组叶片SOD含量的2.29,2.79,2.66,2.02,2.42,2.16倍,差异达到极显著水平(P<0.01),渝薯33、渝薯153、冀薯5号、南紫薯020四种甘薯叶片实验组SOD含量分别是对照组叶片SOD含量的1.84,1.93,1.71,1.37倍,差异达到显著水平(0.01 △表示镉胁迫组与对照组相比差异显著(0.01 GSH是细胞内主要的抗氧化巯基物质之一,具有清除代谢产生的多余活性氧自由基等功能,以减少活性氧对细胞造成的伤害,它还可以在抗氧化、蛋白质巯基保护和氨基酸跨膜运输等发挥重要作用,因此,GSH可以作为细胞氧化还原状态的主要动态指标之一,检测其含量,细胞所处的氧化还原状态能够很好地被反映出来。由图6所见,南渝薯02、渝薯33、淮薯3号、漯徐2号四种甘薯实验组GSH含量分别是对照组甘薯GSH含量的2.1,2.93,2.18,2倍,差异达到极显著水平(P<0.01),渝薯153、冀薯4号、渝薯1号、冀薯5号、南紫薯020、烟薯25六种甘薯实验组GSH含量分别是对照组GSH含量的1.63,1.35,1.36,1.33,1.83,1.83倍,差异达到显著水平(0.01 △表示镉胁迫组与对照组相比差异显著(0.01P CAT具有催化过氧化氢分解为分子氧和水的活性,能使细胞免受过氧化氢毒害的功能,普遍存在于动植物组织,其活性可以一定程度反映生物体的代谢强度及抗寒、抗病能力。由图7可知,渝薯1号实验组甘薯CAT含量是对照组CAT含量的2.02倍,差异达到极显著水平(P<0.01),渝薯153、渝薯33、冀薯4号、南紫薯020、冀薯5号、淮薯3号、南渝薯02、烟薯25、漯徐2号九种甘薯实验组CAT含量分别是对照组CAT含量的1.65,1.83,1.65,1.25,1.42,1.43,1.35,1.36,1.52倍,差异达到显著水平(0.01 △表示镉胁迫组与对照组相比差异显著(0.01 POD广泛存在于生物中,是一种重要的氧化还原酶,其具有参与光呼吸作用、可催化过氧化氢生成水和氧气来消除过氧化氢毒性作用的功能。由图8可知,冀薯4号、渝薯33、南渝薯02、渝薯153、渝薯1号、烟薯25六种甘薯实验组POD含量分别是对照组甘薯含量的2.73,2.26,2.34,2.2,2.87, 2.22倍,差异达到极显著水平(P<0.01),冀薯5号、淮薯3号、南紫薯020、漯徐2号四种甘薯实验组POD含量分别是对照组甘薯含量的1.64,1.77,1.77,1.53倍,差异达到显著水平(0.01 △表示镉胁迫组与对照组相比差异显著(0.01 叶绿素作为叶片中光合系统的最重要的基本组成之一,其含量多少直接决定光合作用强度。植物在镉处理的条件下将直接导致其叶绿素含量降低,使光合作用减弱。本研究中,在镉胁迫下,渝薯33、渝薯153两种甘薯叶绿素含量相对较高,因此其光合作用的效率相较其他八种甘薯更高,淮薯3号、冀薯4号、漯徐2号、渝薯1号四种甘薯叶绿素含量在10种甘薯中相对均衡,冀薯5号、南渝薯02、烟薯25、南紫薯020四种甘薯叶绿素含量最少,其光合作用也最弱;同时,10种甘薯叶片都表现出变黄、萎蔫的症状,相比于对照组,叶绿素含量都极大的降低,这与以往研究结果类似。黄等研究发现,当环境中的镉达到一定含量时,会导致稻苗叶子停止生长,最终枯黄[24]。孙等研究发现随着镉处理的时间变长,黄瓜幼苗中叶绿素含量逐渐降低[25]。刘等发现随着镉处理浓度的增加,金银花叶片逐渐失绿,其叶绿素的含量呈现逐渐下降的趋势[26]。这种现象出现的原因可能是镉影响了叶绿素合成所需要的酶,从而导致叶绿素含量下降,叶片发黄。镉处理会导致植物叶片内叶绿素含量下降,影响植物体内的光合作用及其他生理活动,包括植物对光能的吸收受到影响、植物光合系统所需物质的合成过程也会遭到破坏等。 镉胁迫条件下,植物体内将产生大量的活性氧,这些多余的活性氧将引起生物大分子受到损伤甚至细胞的死亡。本研究中,在镉胁迫条件下,过氧化氢含量最低的是渝薯153、冀薯4号两种甘薯,因此,其受镉胁迫的毒害相对较低,其次是渝薯33、渝薯1号、漯徐2号、南渝薯02四种甘薯,含量最高的则是冀薯5号、淮薯3号、烟薯25、南紫薯020四种甘薯,这四种甘薯受镉胁迫毒害最严重;同时,镉处理条件下甘薯体内过氧化氢的含量会大量增加,这与以往研究类似。于等研究发现镉处理条件下黄瓜胚轴与胚根中的过氧化氢含量显著提高[27]。于研究发现随着镉处理浓度的增加和处理时间的延长,小白菜幼苗叶片中过氧化氢的含量呈现上升趋势[28]。邱等研究得到在镉胁迫下,小麦幼苗生长受到抑制,体内过氧化氢的产生速率显著增加[29]。过氧化氢的增长在某种程度上说明了重金属镉对植物造成的伤害,镉对甘薯植株的毒害随着胁迫时间的延长更严重,最终导致甘薯细胞的稳定性和功能性遭到破坏。 植物具有抗氧化系统会帮助植物抵抗环境胁迫诱导的氧化损伤[30],其在植物对抗外部环境压力时可以发挥重要作用[31]。本实验研究了抗氧化系统在甘薯植株遭受环境胁迫时的保护机制,由图5、图6、图7、图8可知,在镉胁迫下,SOD表现活性较高的是冀薯4号、淮薯3号、渝薯33三种甘薯,南渝薯02、渝薯1号等七种甘薯SOD活性较低;GSH表现活性较高的是渝薯153、冀薯4号等五种甘薯,冀薯5号、淮薯3号等五种甘薯GSH活性相对较低;CAT表现活性较高的是渝薯153、渝薯1号等五种甘薯,表现活性较低的则是冀薯5号、淮薯3号等五种甘薯;POD表现活性较高的五种甘薯是冀薯4号、渝薯33等,表现活性较低的是渝薯1号、淮薯3号等五种甘薯。镉处理导致活性氧含量增加,从而引起SOD、POD等抗氧化酶活性增加,而抗氧化酶活性的增加对于甘薯抵抗镉胁迫具有重要作用。这与以往研究结果类似。曹等研究发现,玉米随着镉浓度的增加其中的SOD等抗氧化酶的活性增加[32]。刘等发现,随着镉浓度的增加,龙葵中的SOD、POD等抗氧化酶的活性增大[33]。赵研究发现,镉胁迫条件下,大豆体内SOD、POD、CAT的活性显著增加[34]。本实验中,镉处理条件下,虽然各类甘薯品种体内的抗氧化酶的活性相比于其对照组都有所增加,但由于品种的不同,抗氧化酶活性的增加也不相同,其中抗氧化酶含量越多,说明其甘薯在镉处理下的抗性也越强,这是由于抗氧化酶的活性直接决定植物在镉胁迫中的耐性所致。总之,在甘薯植株遭受镉胁迫时,体内的抗氧化酶系统会发挥作用,通过抗氧化酶活性的增加,减少重金属镉对甘薯植株造成的伤害。 通过本实验可以知道,在甘薯遭遇镉胁迫,导致其体内活性氧含量上升,会引起甘薯激发自我保护机制,从而使得甘薯体内抗氧化酶活性增加,例如超氧化物歧化酶(SOD)、还原型谷胱甘肽(GSH)等,抗氧化酶活性的提升可以催化活性氧分解成氧气、水等物质,降低甘薯细胞内的氧化胁迫,减轻甘薯细胞受到的毒害,增强甘薯细胞对镉的耐受性,但随着镉毒害加深,超过甘薯所能承受的最大程度,植株叶片便会变黄,最终枯萎。同时,本实验也表明,不同甘薯品种对镉的耐受性不同,部分甘薯体内的抗氧化酶在面对镉胁迫时相比于其他甘薯植株表现的活性更高,叶绿素含量更高,受过氧化氢毒害程度更低。因此,综合叶绿素、过氧化氢毒害、四种抗氧化酶活性分析,渝153、冀薯4号、渝薯33三种甘薯在镉胁迫中受过氧化氢毒害最低,在叶绿素含量、抗氧化酶活性的表现中都相对较好,其对重金属镉抗性潜力最大,淮薯3号、冀薯5号、南渝薯02、渝薯1号四种甘薯在镉胁迫作用下表现次之,南紫薯020、漯徐2号、烟薯25三种甘薯综合叶绿素含量、受过氧化氢毒害程度以及抗氧化酶的活性来观察,这三种甘薯是对镉毒害最为敏感的甘薯。
2.2 镉处理对甘薯叶片内过氧化氢(H2O2)含量的影响
2.3 镉处理对甘薯叶片内超氧化物歧化酶(SOD)活性的影响

2.4 镉处理对甘薯叶片内还原型谷胱甘肽(GSH)含量的影响
2.5 镉处理对甘薯叶片内过氧化氢酶(CAT)活性的影响
2.6 镉处理对甘薯叶片内过氧化物酶(POD)活性的影响
3 小结和讨论
