乌头碱调控miR-181d-5p/DDX3轴对宫颈癌HeLa细胞增殖、凋亡的影响
2023-11-08赵丹丹张素娥苗立业王岩
赵丹丹 张素娥 苗立业 王岩
摘要:目的 探討乌头碱通过调控微小RNA-181d-5p(miR-181d-5p)/DEAD-box RNA解旋酶3(DDX3)轴对宫颈癌HeLa细胞增殖、凋亡的影响。方法 使用不同剂量乌头碱处理宫颈癌HeLa细胞,四甲基偶氮唑盐法检测细胞增殖确定给药剂量。将HeLa细胞分为对照组(正常培养,不进行处理),乌头碱低剂量组(4 mg/L)、中剂量组(8 mg/L)、高剂量组(16 mg/L),顺铂组(5 mg/L),乌头碱高剂量+miR-NC组[16 mg/L乌头碱+转染miR-181d-5p siRNA阴性对照(miR-NC)质粒]及乌头碱高剂量+miR-181d-5p低表达组[16 mg/L乌头碱+转染miR-181d-5p小干扰RNA(siRNA)质粒]。平板克隆形成实验及流式细胞仪分别检测各组HeLa细胞克隆形成数与凋亡情况;实时荧光定量PCR检测HeLa细胞中miR-181d-5p和DDX3 mRNA表达水平;蛋白免疫印迹法检测HeLa细胞中DDX3、细胞周期蛋白D1(Cyclin D1)、增殖细胞核抗原(PCNA)、B淋巴细胞瘤-2(Bcl-2)、Bcl-2相关X蛋白(Bax)、胱天蛋白酶-3(Caspase-3)蛋白表达水平;双萤光素酶报告基因实验验证miR-181d-5p与DDX3的靶向关系。结果 以0.5~64 mg/L的乌头碱分别处理HeLa细胞24 h、48 h、72 h后,对HeLa细胞均有不同程度的抑制作用,选择剂量为4 mg/L、8 mg/L、16 mg/L的乌头碱用于后续实验。与对照组比较,乌头碱低、中、高剂量组及顺铂组HeLa细胞克隆形成数、DDX3 mRNA和蛋白、Cyclin D1、PCNA、Bcl-2蛋白表达水平依次降低(P<0.05),凋亡率、miR-181d-5p、Bax、Caspase-3蛋白表达水平依次升高(P<0.05);与乌头碱高剂量组和乌头碱高剂量+miR-NC组比较,乌头碱高剂量+miR-181d-5p低表达组HeLa细胞克隆形成数、DDX3 mRNA和蛋白、Cyclin D1、PCNA、Bcl-2蛋白表达水平升高(P<0.05),凋亡率、miR-181d-5p、Bax、Caspase-3蛋白表达水平降低(P<0.05);双萤光素酶报告基因检测证实miR-181d-5p与DDX3存在靶向关系。结论 乌头碱可调控miR-181d-5p/DDX3轴,促进miR-181d-5p表达,抑制DDX3表达,进而抑制宫颈癌HeLa细胞增殖,促进细胞凋亡。
关键词:乌头碱;微小RNA-181d-5p;DEAD-box RNA解旋酶3;宫颈癌HeLa细胞
中图分类号:R737.33文献标志码:ADOI:10.11958/20221825
Effects of aconitine regulating miR-181d-5p/DDX3 axis on proliferation and apoptosis of
HeLa cells of cervical cancer
ZHAO Dandan ZHANG Sue MIAO Liye WANG Yan
1 Department of Obstetrics, 2 Department of Gynecology, the Fourth Hospital of Shijiazhuang, Shijiazhuang 050032, China
Abstract: Objective To investigate effects of aconitine on proliferation and apoptosis of cervical cancer HeLa cells by regulating the microRNA-181d-5p (miR-181d-5p)/DEAD-box RNA helicase 3 (DDX3) axis. Methods Cervical cancer HeLa cells were treated with different doses of aconitine, and cell proliferation was detected by tetramethylazolium salt method to determine the dose. HeLa cells were divided into the control group (normal culture, no treatment), the aconitine low dose group (4 mg/L), the aconitine medium dose group (8 mg/L), the aconitine high dose group (16 mg/L), the cisplatin group (5 mg/L), the aconitine high dose + miR-NC group [16 mg/L aconitine + transfected with miR-181d-5p siRNA negative control (miR-NC) plasmid] and the aconitine high dose + miR-181d-5p low expression group [16 mg/L aconitine + transfected miR-181d-5p small interfering RNA (siRNA) plasmid]. Plate clone formation experiment and flow cytometry were used to detect the clonal formation number and apoptosis of HeLa cells in each group. Expression levels of miR-181d-5p and DDX3 messenger RNA (mRNA) in HeLa cells were detected by real-time fluorescent quantitative PCR. Western blot assay was used to detect expression levels of DDX3, Cyclin D1, proliferating cell nuclear antigen (PCNA), B cell lymphocytoma-2 (Bcl-2), Bcl-2 related X protein (Bax) and Caspase-3 protein in HeLa cells. Dual luciferase reporter gene experiment was used to verify the targeting relationship between miR-181d-5p and DDX3. Results HeLa cells were treated with 0.5-64 mg/L aconitine for 24 h, 48 h and 72 h, respectively, and HeLa cells were inhibited to varying degrees. Aconitine doses of 4 mg/L, 8 mg/L and 16 mg/L were selected for follow-up experiments. Compared with the control group, HeLa cell clonal formation number, DDX3 mRNA and protein, Cyclin D1, PCNA and Bcl-2 protein expression levels were decreased successively in the aconitine low, medium and high dose groups and the cisplatin groups (P<0.05), and the apoptosis rate, expression levels of miR-181d-5p, Bax and Caspase-3 protein were increased successively (P<0.05). Compared with the aconitine high dose group and the aconitine high dose + miR-NC group, HeLa cell clonal formation number, DDX3 mRNA and protein, Cyclin D1, PCNA and Bcl-2 protein expression levels were increased in the aconitine high dose + miR-181d-5p low expression group (P<0.05), and the apoptosis rate, expression levels of miR-181d-5p, Bax and Caspase-3 protein were decreased (P<0.05). Dual luciferase reporter gene detection confirmed the targeting relationship between miR-181d-5p and DDX3. Conclusion Aconitine can regulate the miR-181d-5p/DDX3 axis, promote the expression of miR-181d-5p, inhibit the expression of DDX3, and then inhibit the proliferation of cervical cancer HeLa cells and promote cell apoptosis.
Key words: aconitine; microRNA-181d-5p; DEAD-box RNA helicase 3; cervical cancer HeLa cells
宫颈癌是女性生殖系统常见的恶性肿瘤,其发病率呈年轻化趋势,早期宫颈癌以手术治疗为主,晚期及复发性宫颈癌以放化疗为主,耐药导致晚期患者预后较差[1]。寻找疗效高、毒副作用小的药物对于宫颈癌的治疗具有重要意义[2]。乌头碱是乌头、附子等中草药的有效成分,具有镇静、抗炎、镇痛作用[3]。研究表明乌头碱亦具有抗肿瘤作用,可抑制肝癌细胞的增殖、迁移和侵袭[4-5],但其对宫颈癌的作用尚不清楚。微小RNA-181d-5p(miR-181d-5p)为microRNA一种,在肾细胞癌、非小细胞肺癌中表达下调,上调miR-181d-5p表达可抑制肿瘤细胞增殖,促进细胞凋亡[6-7]。StarBase数据库预测显示,miR-181d-5p可靶向调控DEAD-box RNA解旋酶3(DEAD-box RNA helicase 3,DDX3)表达。DDX3是具有ATP酶依赖性的DEAD盒RNA解旋酶,在基因转录、剪接与翻译过程中发挥调控作用。研究显示,DDX3在宫颈癌中表达上调,其过表达可促进宫颈癌细胞的增殖、迁移和侵袭[8]。miR-181d-5p是否靶向调控DDX3影响宫颈癌细胞增殖与凋亡尚未可知。本研究通过体外培养宫颈癌HeLa细胞,探索乌头碱对HeLa细胞增殖、凋亡及对miR-181d-5p/DDX3轴的影响,探索乌头碱在宫颈癌中的作用机制。
1 材料与方法
1.1 主要试剂与仪器 人宫颈癌HeLa细胞购自合肥合生生物工程有限公司。乌头碱(原料药,纯度≥98%)、顺铂(原料药,纯度≥99.5%)、DMEM/F12培养基、Lipofectamine 3000试剂盒、TRIzol试剂、ECL显色液均购自成都曼斯特生物科技有限公司;miR-181d-5p小干扰RNA(siRNA)质粒、miR-181d-5p siRNA阴性对照(miR-NC)质粒、DDX3野生型(DDX3-WT)质粒、DDX3突变型(DDX3-MUT)质粒、miR-181d-5p模拟物(mimics)质粒、miR-181d-5p mimics阴性对照质粒、miR-181d-5p、U6引物均由广州锐博生物有限公司合成;四甲基偶氮唑盐(MTT)、萤光素酶检测试剂盒、逆转录试剂盒均购自美国Sigma-Aldrich公司;PCR试剂盒(SYBR qPCR mix)、AnnexinV-FITC/PI凋亡检测试剂盒、蛋白提取试剂盒、羊抗鼠二抗均购自北京启维益成科技有限公司;鼠源DDX3、细胞周期蛋白D1(Cyclin D1)、增殖细胞核抗原(PCNA)、B淋巴细胞瘤-2(Bcl-2)、Bcl-2相关X蛋白(Bax)、胱天蛋白酶-3(Caspase-3)、β-肌动蛋白(β-actin)单克隆抗体均购自英国Abcam公司。酶标仪(型号SYNERGY H1)、显微镜(型号MVX10)、实时荧光定量PCR(qPCR)仪(型号SLAN-96P)均购自西安淳风生物科技有限公司;流式细胞仪(型号Fortessa X-20)、凝胶成像系统(型号E5001-SDB)均购自廊坊纵世科技有限公司。
1.2 方法
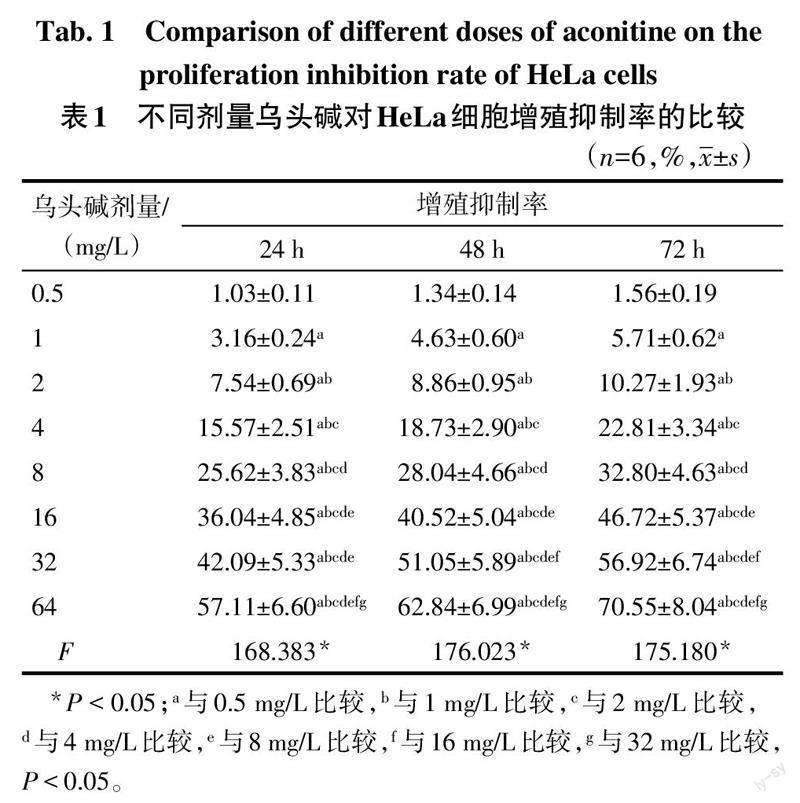
1.2.1 MTT法检测不同剂量乌头碱对HeLa细胞增殖的影响 宫颈癌HeLa细胞接种于DMEM/F12培养基中(含有10%胎牛血清及1%链霉素与青霉素)培养。取对数生长期细胞,按照5×104个/孔接种于96孔板中,分别使用剂量为0.5、1、2、4、8、16、32、64 mg/L的乌头碱(用DMEM/F12培养液稀释)处理HeLa细胞,分别培养24、48和72 h后,每孔加入10 μL的MTT溶液,孵育4 h再加入150 μL二甲基亚砜溶液终止反应,在酶标仪490 nm波長处检测吸光度(A)值,另设只添加MTT溶液、二甲基亚砜的HeLa细胞作为空白组。细胞增殖抑制率(%)=(1-各剂量乌头碱组A值/空白组A值)×100%。
1.2.2 HeLa细胞培养及分组 取对数生长期的HeLa细胞,按照1.2×106个/孔接种于6孔板中,使用DMEM/F12培养基进行培养,分为对照组(正常培养,不进行处理)、乌头碱低剂量组、乌头碱中剂量组、乌头碱高剂量组、顺铂组、乌头碱高剂量+miR-NC组、乌头碱高剂量+miR-181d-5p低表达组。乌头碱低、中、高剂量组和顺铂组分别向含有HeLa细胞的DMEM/F12培养基中加入剂量为4、8、16 mg/L乌头碱和5 mg/L顺铂[9];乌头碱高剂量+miR-NC组、乌头碱高剂量+miR-181d-5p低表达组分别采用Lipofectamine 3000试剂盒转染miR-NC质粒、miR-181d-5p siRNA质粒,严格按照试剂盒说明书操作,然后再向培养基中加入剂量为16 mg/L的乌头碱。每组重复6次。
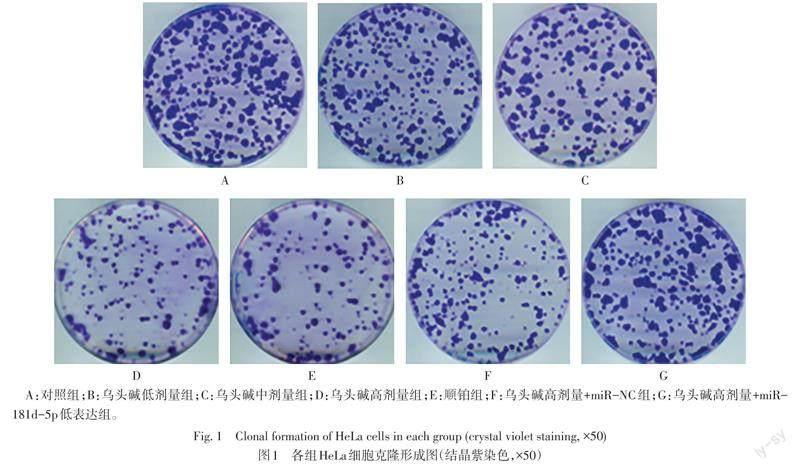
1.2.3 平板克隆形成实验 取对数生长期的HeLa细胞,以每孔800个细胞接种于6孔板中,置于37 ℃、5%CO2培养箱中培养14 d,每隔2~3 d换液并观察细胞形态,培养皿中出现肉眼可见的克隆时,使用4%多聚甲醛固定15 min,0.1%结晶紫染色30 min,观察细胞克隆形成数。
1.2.4 流式细胞仪检测HeLa细胞凋亡 细胞培养48 h后,胰酶消化细胞,PBS重悬,每孔分别加入5 μL的AnnexinV-FITC和PI,避光孵育20 min,流式细胞仪检测细胞凋亡情况,细胞凋亡率为早期凋亡与晚期凋亡细胞占总细胞的百分比。
1.2.5 qPCR检测HeLa细胞中miR-181d-5p和DDX3 mRNA的表达 细胞培养48 h后,TRIzol试剂提取细胞总RNA,逆转录试剂盒合成cDNA后,采用qPCR法检测各组HeLa细胞中miR-181d-5p和DDX3 mRNA相对表达量。miR-181d-5p上游引物:5'-GCAAACATTCATTGTTGTCGGT-3',下游引物:5'-CCAGTGCAGGGTCCGAGGT,产物大小22 bp;DDX3上游引物:5'-ATCCTGGACTCAGCAGTCAGATGC-3',下游引物:5'-TGCGTAATGGCATGGTAACTTAAT-3',产物大小20 bp;U6上游引物:5'-CGCTTCGGCAGCACATATACTA-3',下游引物:5'-GGAACGCTTCACGAATTTGC-3',产物大小15 bp。PCR反应体系25 μL,其中SYBR qPCR Mix 12.5 ?L,cDNA 模板1 ?L,上下游引物各0.5 ?L,加ddH2O至25 μL。反应条件:95 ℃ 60 s;95 ℃ 15 s,60 ℃ 15 s,72 ℃ 45 s,40个循环。以U6为内参,采用2-??Ct法计算miR-181d-5p和DDX3 mRNA相对表达量。
1.2.6 蛋白免疫印迹法检测HeLa细胞中DDX3、Cyclin D1、PCNA、Bax、Bcl-2、Caspase-3蛋白的表达 细胞培养48 h后,提取各组细胞总蛋白,BCA法测定蛋白浓度,取30 μg蛋白加上样缓冲液煮沸变性后,电泳分离蛋白,湿转法转膜,加入5%的牛血清白蛋白封闭1 h,分别加入鼠源DDX3(1∶1 030)、Cyclin D1(1∶1 030)、PCNA(1∶1 030)、Bax(1∶1 460)、Bcl-2(1∶1 460)、Caspase-3(1∶1 460)、β-actin(1∶1 000)抗体,4 ℃孵育过夜,PBST漂洗3次,加羊抗鼠二抗室温孵育2 h,ECL显色液显色,凝胶成像系统显影,以β-actin为内参,Image J软件分析目的蛋白相对表达量。
1.2.7 双萤光素酶报告基因实验 通过靶基因在线预测网站StarBase(https://starbase.sysu.edu.cn/)预测miR-181d-5p与DDX3是否存在靶向结合位点。将HeLa细胞按照5×105个/孔接种于24孔板,培养24 h后,分别将DDX3-WT和DDX3-MUT质粒与miR-181d-5p mimics、miR-181d-5p mimics阴性对照质粒共转染HeLa细胞24 h,檢测各组细胞相对萤光素酶活性。
1.3 统计学方法 采用SPSS 25.0进行数据分析,符合正态分布的计量资料以x±s表示,2组间比较采用独立样本t检验;多组间比较采用单因素方差分析,组间多重比较行SNK-q检验。P<0.05为差异有统计学意义。
2 结果
2.1 不同剂量乌头碱对HeLa细胞增殖的影响 使用0.5~64 mg/L的乌头碱分别处理HeLa细胞24 h、48 h、72 h,随乌头碱剂量的增高,其对HeLa细胞的增殖抑制作用呈增强趋势(P<0.05),见表1。为保证一定的细胞存活数量以及蛋白质提取量,选择4 mg/L、8 mg/L、16 mg/L的乌头碱用于后续实验。
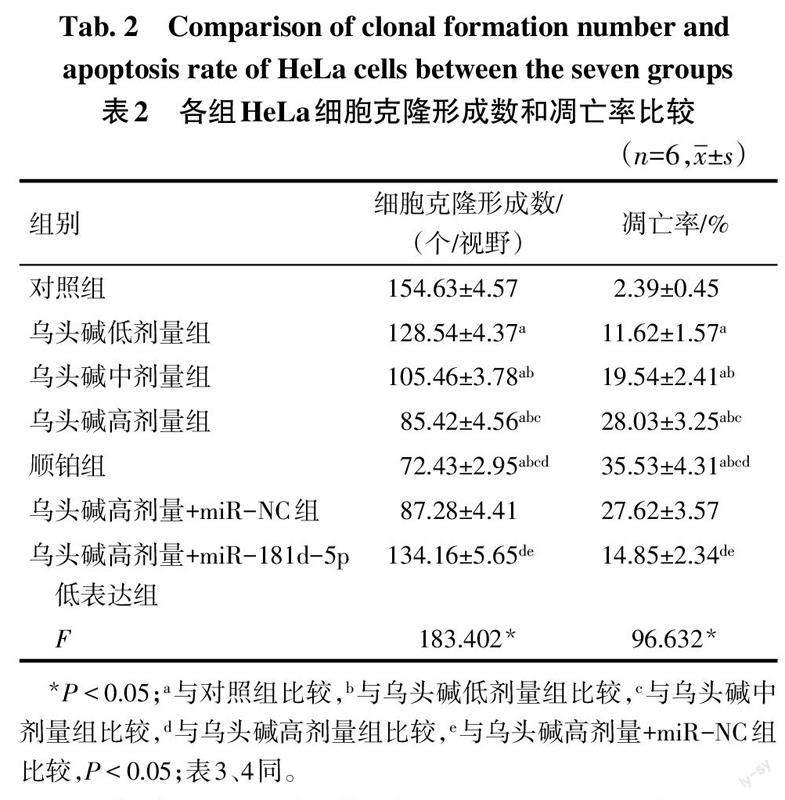
2.2 各组HeLa细胞克隆形成数和凋亡率比较 与对照组比较,乌头碱低、中、高剂量组及顺铂组HeLa细胞克隆形成数依次降低,凋亡率依次升高(P<0.05);与乌头碱高剂量组和乌头碱高剂量+miR-NC组比较,乌头碱高剂量+miR-181d-5p低表达组HeLa细胞克隆形成数增多,凋亡率降低(P<0.05)。见表2,图1、2。
2.3 各组HeLa细胞中miR-181d-5p和DDX3 mRNA表达水平比较 与对照组比较,乌头碱低、中、高剂量组及顺铂组HeLa细胞中miR-181d-5p表达水平依次升高,DDX3 mRNA表达水平依次降低(P<0.05);与乌头碱高剂量组和乌头碱高剂量+miR-NC组比较,乌头碱高剂量+miR-181d-5p低表达组HeLa细胞中miR-181d-5p表达水平降低,DDX3 mRNA表达水平升高(P<0.05)。见表3。
2.4 各组HeLa细胞中DDX3、Cyclin D1、PCNA、Bax、Bcl-2、Caspase-3蛋白表达水平比较 与对照组比较,乌头碱低、中、高剂量组及顺铂组HeLa细胞中DDX3、Cyclin D1、PCNA、Bcl-2蛋白表达水平依次降低,Bax、Caspase-3蛋白表达水平依次升高(P<0.05);与乌头碱高剂量组和乌头碱高剂量+miR-NC组比较,乌头碱高剂量+miR-181d-5p低表达组HeLa细胞中DDX3、Cyclin D1、PCNA、Bcl-2蛋白表达水平升高,Bax、Caspase-3蛋白表达水平降低(P<0.05)。见图3、表4。
2.5 miR-181d-5p与DDX3的靶向关系 StarBase数据库预测显示,miR-181d-5p与DDX3 mRNA 3'UTR区有结合位点,见图4。双萤光素酶报告基因检测结果显示,与共转染DDX3-WT和miR-181d-5p mimics阴性对照组比较,共转染DDX3-WT和miR-181d-5p mimics组相对萤光素酶活性降低(P<0.05),见图5。证明miR-181d-5p与DDX3存在靶向关系。
3 讨论
乌头碱是乌头类生物碱,具有抗心力衰竭、抗炎、免疫抑制及镇痛等多种药理学作用,具有广阔的临床应用前景[10]。在抗肿瘤方面,乌头碱可通过多种途径(如核因子-κB通路、雌激素受体)抑制肿瘤细胞的增殖与迁移,促进细胞凋亡[11-12]。本研究使用0.5~64 mg/L的乌头碱分别处理HeLa细胞24 h、48 h、72 h后,结果显示,乌头碱对HeLa细胞增殖均有不同程度的抑制作用。Wang等[13]研究表明,乌头碱可诱导卵巢癌A2780细胞凋亡和DNA损伤,抑制卵巢癌生长。另有研究显示,乌头碱通过下调核因子-κB表达以剂量和时间依赖性方式抑制胰腺癌细胞生长,促进细胞凋亡,且在胰腺癌的异种移植小鼠模型中,乌头碱可抑制肿瘤生长并增加细胞凋亡[14]。本研究结果显示,使用4、8、16 mg/L的乌头碱处理HeLa细胞后,HeLa细胞克隆形成数、Cyclin D1、PCNA、Bcl-2蛋白表达水平依次降低,凋亡率、Bax、Caspase-3蛋白表达水平依次升高,表明乌头碱可抑制宫颈癌HeLa细胞增殖,促进细胞凋亡,与在其他肿瘤中研究[13-14]结果类似,提示乌头碱在宫颈癌中表现为抗肿瘤作用,但乌头碱对HeLa细胞增殖、凋亡影响的具体作用机制尚不清楚。
有研究显示miR-181d-5p在不同肿瘤中发挥促癌或抑癌的作用,如在结肠癌细胞中表达上调,抑制其表达可抑制结肠癌细胞的增殖、迁移、侵袭能力[15]。而miR-181d-5p在肺腺癌、胶质瘤中表达下调,发挥抑癌作用,上调其表达可抑制肿瘤细胞增殖,促进细胞凋亡[16-17]。本研究结果表明,乌头碱可能具有上调miR-181d-5p表达的作用,与乌头碱高剂量组和乌头碱高剂量+miR-NC组比较,乌头碱高剂量+miR-181d-5p低表达组HeLa细胞克隆形成数、Cyclin D1、PCNA、Bcl-2蛋白表达水平升高,凋亡率、miR-181d-5p、Bax、Caspase-3蛋白表达水平降低,表明抑制miR-181d-5p表达可逆转乌头碱对HeLa细胞增殖的抑制作用和凋亡的促进作用,提示miR-181d-5p在宫颈癌中可能发挥抑癌基因的作用,这与李祥双等[18]研究结果类似。以上研究提示乌头碱可能通过上调miR-181d-5p表达从而抑制HeLa细胞增殖,促进细胞凋亡,这与miR-181d-5p在宫颈癌细胞中的作用有一定关系。
miRNA通过靶向调控下游基因的表达而发挥生物学功能。本研究通过双萤光素酶报告基因检测显示,miR-181d-5p与DDX3存在靶向关系。DDX3是依赖ATP的RNA解旋酶,可参与RNA转录、翻译和降解等过程,DDX3还可参与细胞周期与细胞凋亡的调控,在肿瘤的发生发展中发挥重要作用,高表达的DDX3与卵巢癌侵袭和乳腺癌患者不良预后有关[19-20]。研究显示,环状RNA 0035266通过miR-181d-5p/DDX3分子轴促进4-甲基亚硝胺基-1-3-吡啶基-1-丁酮+脂多糖诱导的人支气管上皮细胞中的炎症反应[21]。本研究使用4、8、16 mg/L乌头碱处理宫颈癌细胞后,DDX3 mRNA和蛋白表达水平依次降低;而乌头碱高剂量+miR-181d-5p低表达组HeLa细胞中DDX3 mRNA和蛋白表达水平高于乌头碱高剂量组和乌头碱高剂量+miR-NC组,进一步证实miR-181d-5p与DDX3的靶向调控关系。以上结果表明乌头碱通过上调miR-181d-5p表达,抑制DDX3表达,抑制HeLa细胞增殖,诱导细胞凋亡,这对于宫颈癌的治疗有一定积极意义。
综上所述,乌头碱可抑制宫颈癌HeLa细胞增殖,促进细胞凋亡,可能是通过调控miR-181d-5p/DDX3軸发挥作用。本研究仅对乌头碱的作用机制进行了初步探究,研究结果尚未能在动物模型中予以验证,因此其涉及的相关分子通路仍有待今后进一步研究。
参考文献
[1] 彭巧华,吕卫国. 2022年第1版《NCCN子宫颈癌临床实践指南》解读[J]. 实用肿瘤杂志,2022,37(3):205-214. PENG Q H,LYU W G. Interpretation of NCCN guidelines for cervical cancer,version 1. 2022[J]. Journal of Practical Oncology,2022,37(3):205-214. doi:10.13267/j.cnki.syzlzz.2022.034.
[2] ZHAO M,GU R Y,DING S R,et al. Risk factors of cervical cancer among ethnic minorities in Yunnan Province,China:a case-control study[J]. Eur J Cancer Prev,2022,31(3):287-292. doi:10.1097/CEJ.0000000000000704.
[3] LI L,ZHANG L,LIAO T,et al. Advances on pharmacology and toxicology of aconitine[J]. Fundam Clin Pharmacol,2022,36(4):601-611. doi:10.1111/fcp.12761.
[4] SONG N,MA J,ZHANG X,et al. Lappaconitine hydrochloride induces apoptosis and S phase cell cycle arrest through MAPK signaling pathway in human liver cancer HepG2 cells[J]. Pharmacogn Mag,2021,17(74):334-341.doi:10.4103/pm.pm_251_20.
[5] 熊慧生,蒋参,高瑞,等. 乌头碱对肝癌MHCC97细胞生长、侵袭和迁移的调控作用及机制研究[J]. 中国免疫学杂志,2018,34(5):688-691. XIONG H S,JIANG C,GAO R,et al. Regulatory effects and mechanism of aconitine on proliferation,invasion and migration of hepatoma carcinoma cell MHCC97[J]. Chinese Journal of Immunology,2018,34(5):688-691. doi:10.3969/j.issn.1000-484X.2018.05.009.
[6] DING M,ZHAO X,CHEN X,et al. Cancer-associated fibroblasts promote the stemness and progression of renal cell carcinoma via exosomal miR-181d-5p[J]. Cell Death Discov,2022,8(1):439. doi:10.1038/s41420-022-01219-7.
[7] 夏炳辉,张振山,姚天宇,等. miR-181-5p下调人非小细胞肺癌细胞KLF6表达并抑制其增殖、迁移和侵袭[J]. 生物技术,2022,32(5):587-591. XIA B H,ZHANG Z S,YAO T Y,et al. miR-181-5p down-regulates the expression of human non-small cell lung cancer cell KLF6 and inhibits its proliferation,migration and invasion[J]. Biotechnology,2022,32(5):587-591. doi:10.16519/j.cnki.1004-311x.2022.05.0093.
[8] 郭芳芳,赵瑞皎,李杜娟,等. DDX3表达上调在人宫颈癌细胞增殖中的作用分析[J]. 中华病理学杂志,2021,50(2):119-124. GUO F F,ZHAO R J,LI D J,et al. Role of up-regulated DDX3 in the proliferation of human cervical cancer cells[J]. Chin J Pathol,2021,50(2):119-124. doi:10.3760/cma.j.cn112151-20200519-00394.
[9] 李萍,姬白嫣,魏娟,等. 藏红花素对人宫颈癌Hela细胞顺铂耐药性的影响[J]. 新乡医学院学报,2021,38(5):401-405. LI P,JI B Y,WEI J,et al. Effect of crocin on cisplatin resistance of human cervical cancer Hela cells[J]. Journal of Xinxiang Medical University,2021,38(5):401-405. doi:10.7683/xxyxyxb.2021.05.001.
[10] GAO Y,FAN H,NIE A,et al. Aconitine:A review of its pharmacokinetics,pharmacology,toxicology and detoxification[J]. J Ethnopharmacol,2022,293:115270. doi:10.1016/j.jep.2022.115270.
[11] LUAN S,GAO Y,LIANG X,et al. Aconitine linoleate,a natural lipo-diterpenoid alkaloid,stimulates anti-proliferative activity reversing doxorubicin resistance in MCF-7/ADR breast cancer cells as a selective topoisomerase Ⅱα inhibitor[J]. Naunyn Schmiedebergs Arch Pharmacol,2022,395(1):65-76. doi:10.1007/s00210-021-02172-5.
[12] 周小剑,李玉华,唐湘,等. 乌头碱抑制食管癌EC-1细胞增殖与侵袭并诱导其凋亡作用研究[J]. 中国医药生物技术,2019,14(4):335-340. ZHOU X J,LI Y H,TANG X,et al. Aconitine inhibits the proliferation and invasion while induces the apoptosis of esophageal cancer EC-1 cells[J]. Chinese Medicinal Biotechnology,2019,14(4):335-340. doi:10.3969/j.issn.1673-713X.2019.04.008.
[13] WANG X,LIN Y,ZHENG Y. Antitumor effects of aconitine in A2780 cells via estrogen receptor β-mediated apoptosis,DNA damage and migration[J]. Mol Med Rep,2020,22(3):2318-2328. doi:10.3892/mmr.2020.11322.
[14] JI B L,XIA L P,ZHOU F X,et al. Aconitine induces cell apoptosis in human pancreatic cancer via NF-κB signaling pathway[J]. Eur Rev Med Pharmacol Sci,2016,20(23):4955-4964.
[15] 柯东平,朱金祥,毛俊倩,等. miR-181d-5p对人结肠癌细胞凋亡的影响及机制[J]. 贵州医科大学学报,2019,44(7):814-820. KE D P,ZHU J X,MAO J Q,et al. Effect and mechanism of microRNA-181d-5p on apoptosis of human colon cancer cells[J]. Journal of Guizhou Medical University,2019,44 (7):814-820. doi:10.19367/j.cnki.1000-2707.2019.07.014.
[16] DONG X,LIU Y,DENG X,et al. C1GALT1,Negatively regulated by miR-181d-5p,promotes tumor progression via upregulating RAC1 in lung adenocarcinoma[J]. Front Cell Dev Biol,2021,9:707970. doi:10.3389/fcell.2021.707970.
[17] KHALIL S,FABBRI E,SANTANGELO A,et al. miRNA array screening reveals cooperative MGMT-regulation between miR-181d-5p and miR-409-3p in glioblastoma[J]. Oncotarget,2016,7(19):28195-28206. doi:10.18632/oncotarget.8618.
[18] 李祥双,于海霞,叶燕珍,等. miR-181d-5p对宫颈癌细胞放射敏感性的影响及机制[J]. 山东医药,2022,62(22):42-46. LI X S,YU H X,YE Y Z,et al. Influence and mechanism of miR-181d-5p on radiosensitivity of cervical cancer cells[J]. Shandong Medical Journal,2022,62(22):42-46. doi:10.3969/j.issn.1002-266X.2022.22.010.
[19] NI J,CHEN L,LING L,et al. MicroRNA-196a promotes cell proliferation and inhibits apoptosis in human ovarian cancer by directly targeting DDX3 and regulating the PTEN/PI3K/AKT signaling pathway[J]. Mol Med Rep,2020,22(2):1277-1284. doi:10.3892/mmr.2020.11236.
[20] VAN DER POL C C,MOELANS C B,MANSON Q F,et al. Cytoplasmic DDX3 as prognosticator in male breast cancer[J]. Virchows Arch,2021,479(4):647-655. doi:10.1007/s00428-021-03107-4.
[21] HUA Q,LIU Y,LI M,et al. Tobacco-related exposure upregulates Circ_0035266 to exacerbate inflammatory responses in human bronchial epithelial cells[J]. Toxicol Sci,2021,179(1):70-83. doi:10.1093/toxsci/kfaa163.
(2022-11-14收稿 2023-03-14修回)
(本文編辑 陈丽洁)
