乒乓球运动员动作预测时空整合的大脑网络连接特征
2023-11-04高思雨梁轶睿伦梓文周成林
高思雨,梁轶睿,伦梓文,周成林
乒乓球作为一项球速快、球飞行轨迹变化多的运动项目。运动员需要在短时间内对球飞行时的旋转轨迹及最终落点进行辨别、预测,选择合适的策略应对、调整拍面角度,进行有效的击球还击。动作预测是指根据少量不完整的信息对动作最终指向结果作出判断的过程,动作预判过程需要运动员具有在时间与空间维度上对动作的精准预测能力,才能完成高质量击球,提高获胜的可能性。
长期专项训练形成的知觉-认知技能可促进运动员预测能力提升[1-2],在时间维度上体现为专家运动员能够根据对手的早期运动信息进行后续动作及动作结果的预测。Farrow等采用时间阻断技术进行的研究发现,相比于乒乓球新手,专家级选手可更早利用有效信息对发球方向进行预测[3]。Runswick 等在一项棒球研究中发现,在越接近投球点时,击球手对球方向的预测就越依靠投球手的身体运动信息和球飞行信息[4],说明专家与新手运动员相比,在预测晚期阶段(球飞出前),更为依赖运动员的运动信息。而在这个过程中,动作模式识别能力是专家运动员预测加工的关键。研究发现,专家运动员对动作模式的记忆提取与识别更强,并且专家运动员对运动项目早期更强的动作模式识别能力有助于对动作结果进行预测[5-6]。
神经电生理的相关研究为运动经验如何促进动作识别提供了证据。Orgs 等通过对比专业舞者和非专业舞者在观看并评判专业舞蹈动作和日常生活动作视频时发现,专业舞者在观看舞蹈动作时,α节律的事件相关去同步活动显著大于非专业舞者[7]。α节律通常被认为与大脑皮层兴奋性有关,α节律同步性增加代表皮层兴奋性减弱,而α节律去同步性增加则代表皮层兴奋性增加。因此,在动作预测之前的动作观察阶段,专家运动员大脑皮层兴奋性的增加可能反映了对动作模式识别的更高唤醒程度。θ节律代表的是工作记忆的加工过程[8-10],与大脑对信息的编码[9]和记忆的加工、大脑工作负荷[11]呈正相关关系,并且θ节律在大脑的额叶、顶叶和前额叶之间功能的强交互作用形成了一个能够促进刺激加工、整合和信息分类的网络[12-13],能增强θ节律的注意力控制功能[14]。目前关于运动学信息如何与α节律、θ节律相互作用于动作预测的时间过程还缺乏深入的探究。
在空间维度上,共同编码理论提出,专家运动员对动作预测的知觉优势来自于运动与知觉表征之间的紧密联系,二者共享一套编码系统[15],并建立相应的“内部模型”。内部模型指运动指令与感觉结果间形成相互转化的对应关系。长期运动经验促进运动员建立完善动作内部模型,并在感知动作过程中进行调用,通过人体运动系统对运动信息进行模拟并匹配至潜在的动作结果从而完成预测。
动作观察网络是内部模型的重要神经网络。该神经网络仅仅通过观察动作就会被激活,在感知他人的动作中起着至关重要的作用[16]。动作观察网络涉及到以颞上沟后部(缩写为pSTS)、顶下小叶(缩写为IPL)与前运动皮层(缩写为PMC)为核心节点的众多脑区。PMC 与IPL 组成的额顶网络系统在动作意图理解中起到重要作用,并反映了对运动信息的高级加工[17-18]。而pSTS 主要作用是输入视觉信息,是运动信息表征的输入节点[19],不具备对运动经验的特异性贡献[20]。尽管大部分的研究强调了AON 在知觉预期过程中的重要性,但研究结果尚未一致。
因此,本研究拟采用EEG、fMRI 技术,探索运动信息对动作预测的时空特点,以及AON 的脑区激活与网络连接特征。
1 乒乓球运动员动作预测的大脑活动时间特征
1.1 研究目的与假设
探究乒乓球运动员如何整合不同时间进程的信息进行动作预测。通过EEG 技术记录其实验过程中的脑电变化,分析预测过程的时间特点。假设:运动信息是影响预测表现的主要变量参数。
1.2 研究设计
单因素被试内设计。自变量为运动信息量(少量为800 ms,大量为900 ms),因变量为视频观看过程中对发球者动作预测表现(正确率)、对发球者动作观看时的α同步值和θ同步值。
1.2.1 被试
使用软件“G-power”计算所需样本量,根据以往类似研究结果,效应量在0.13~0.50。设置置信水平为0.95,统计检验力为0.95,选择最低效应量=0.13 进行计算,最低样本量为16 人。最终本实验共招募21 名被试,均为上海体育大学在校学生,并获得乒乓球国家二级及以上运动员证书(以下以“专家”指代)。其中男性为10 人,女性为11 人。年龄为18~25 岁,平均年龄为20.41 岁。其中一名男性自我报告为左利手,其余被试均为右利手。
1.2.2 实验材料
实验刺激材料为本课题组自行录制的乒乓球发球视频,发球者为2 名发球较好的乒乓球运动员,使用右手直握拍法,站在球台左侧正手发球。发球动作分为左侧短球、左侧长球、右侧短球、右侧长球4类,各重复15 遍,由健将级乒乓球运动员挑选发球较好的10 组动作。最终得到40 个不同的发球视频。
1.2.3 实验程序
本实验为组块设计,正式实验共有6 个组,每组30 个试次。每个组块之间强制休息30 s。实验前提供20 个练习试次让被试理解实验要求和熟悉反应按键。练习试次和正式实验试次均不反馈被试判断的正确性。每个试次由一个1 000 ms 的注视点开始,随后是一段发球动作视频,4 种发球类型随机呈现,视频长度根据运动信息量分为800 ms(少量)和900 ms(大量)2种。视频结束后立刻切换到下一页面,防止被试根据最后一帧进行观察和预测。视频结束后为反应阶段,屏幕显示内容为“请进行落点预测”。被试根据自己的预测使用“F”“V”“J”“N”键进行预测反应,按键分别对应“左上”“左下”“右上”“右下”。屏幕上也会有相应的提示。被试必须熟记按键所包含的反应内容才允许进入正式实验。按键结束后是500 ms 的空屏,接着开始下一个试次。实验流程如图1 所示。

图1 实验流程
1.2.4 数据采集与分析
1.2.4.1 行为数据采集
根据乒乓球落点的运动信息量(少量、大量)对正确率进行配对样本t 检验,显著性阈值设为p<0.05。
1.2.4.2 脑电数据记录与处理
使用符合国际“10-10”系统的64 导活动电极脑电帽(actiCAP;Brain Products GmbH,Germany)采集脑电,并使用Brain Vision Recorder 2.0(Brain Products GmbH,Germany),采样频率为1 000 Hz。FCz 作为参考电极,GND 作为接地电极。FT9 和FT10 分别贴在被试左眼外侧1 cm 位置和右眼下方1 cm 位置采集水平眼电和垂直眼电。记录时不使用在线滤波。
脑电数据由开源工具包Letswave7(https://letswave.cn)和Matlab R2018b(MathWorks Inc.,MA,USA)进行分析。脑电数据处理步骤包括:1)在数据查看器中目视检查,对异常通道进行插值。2)使用Butterworth 滤波器设置通过范围为0.05~30 Hz。3)使用独立成分分析(ICA)分离脑电成分,设置成分数为40,人工剔除眼电成分。4)根据实验条件对数据进行分段。5)使用自动去伪迹剔除振幅大于±100 mμV 的试次。6)使用全脑平均的方法对全脑电极进行重参考,并还原FCz点。7)对每个实验条件下的脑电数据进行平均叠加。8)将注视点呈现前500 ms 到0 ms 作为基线对数据进行校正。
根据以往研究,选取θ频段(5~8 Hz)和α频段(9~12 Hz)进行分析。θ频段选取FCz 电极点进行分析,α频段选取Pz、P3、P4、P5、P6、P7、P8、POz、PO3、PO4、PO7、PO8、Oz、O1、O2电极点进行分析。以50 ms 作为一个时间窗,从视频开始到球与球拍接触为止,共18 个时间窗(T1、T2、T3、T4、T5、T6、T7、T8、T9、T10、T11、T12、T13、T14、T15、T16、T17、T18)。由于本研究重点关注球与球拍接触前的脑电特征,因此,选取T10~T16作为感兴趣时间窗,对α和θ同步化平均值进行统计检验。
1.3 实验结果
1.3.1 行为结果
对乒乓球运动员分别在少量运动信息和多量运动信息条件下的反应正确率进行配对样本t 检验,结果表明,少量运动信息下的正确率与大量运动信息下的正确率相比,差异显著(t=-3.163,p=0.03<0.05),结果见表1。表明运动信息量是影响预测准确性的因素。

表1 不同运动信息条件下落点预测正确率比较
1.3.2 脑电结果
2 种运动信息量的EEG 时频情况如图2 所示,横轴代表时间段(0 点表示视频开始,纵轴代表频率)。
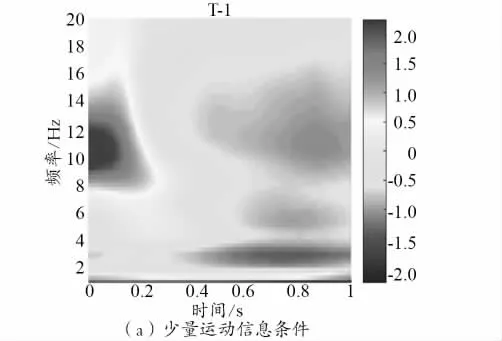
图2 被试在2 种运动信息量条件下观看动作视频的时频情况
2 种运动信息量条件下的α节律同步化活动值和θ节律同步化活动值在感兴趣时间窗内的平均值和标准差见表2 和表3。
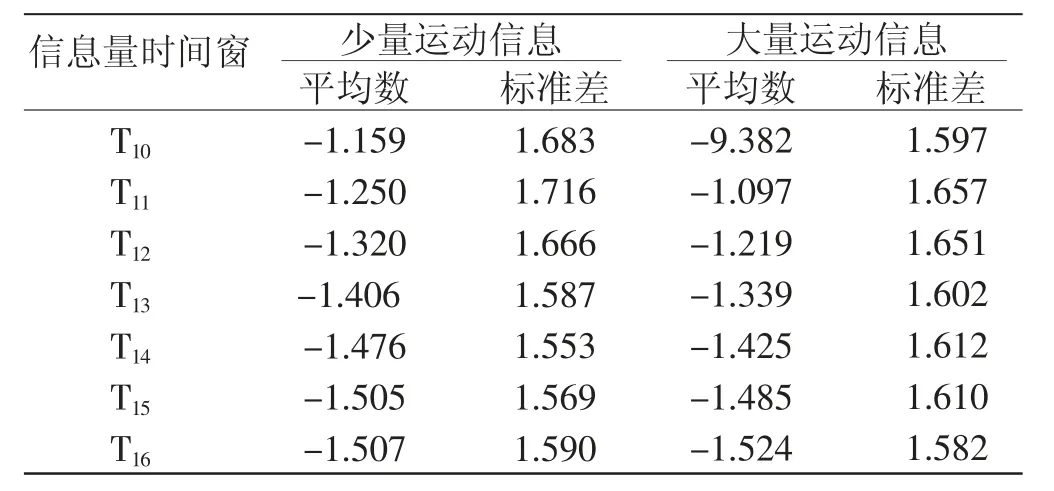
表2 不同运动信息量条件下的顶枕叶-α 值描述统计结果

表3 不同运动信息量条件下的FCz-θ 值描述统计结果
1.3.2.1α节律同步化活动
对α节律同步化活动进行2(运动信息量)×7(时间窗)两因素重复测量方差分析。经Mauchly’s 球形假设检验,结果显示不通过。又使用Greenhouse &Geisser 方法校正,结果发现,运动信息量主效应不显著;时间窗主效应显著;运动信息量×时间窗交互作用不显著0.048]。使用Bonferroni 法对时间窗进行事后多重比较,发现T10与T13(p=0.015)、T10与T14(p=0.011)、T10与T15(p=0.013)、T10与T16(p=0.012)差异显著;T11与T13(p=0.031)差异显著。其他时间窗口间(10 与T11、T10与T12、T11与T12、T11与T14、T11与T15、T11与T16、T12与T13、T12与T14、T12与T15、T12与T16、T13与T14、T13与T15、T13与T16、T14与T15、T14与T16、T15与T16)差异不显著。α节律同步化活动在时间窗内波幅变化趋势如图3 所示。
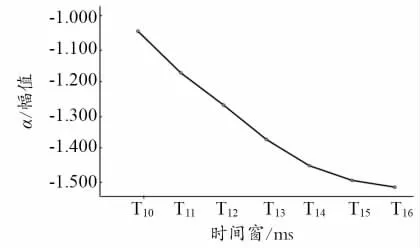
图3 α 频率同步化值随时间窗的变化
1.3.2.2θ节律同步化活动
对θ节律同步化活动进行2(运动信息量)×7(时间窗)两因素重复测量方差分析。经Mauchly’s 球形假设检验,结果显示不通过,又使用Greenhouse &Geisser 方法校正,结果发现,运动信息量主效应不显著;时间窗主效应显著;运动信息量×时间窗交互作用不显著0.011]。使用Bonferroni 法对时间窗进行事后多重比较发现,T10与T11(p=0.002)、T12(p=0.002)、T13(p=0.001)、T14(p<0.001)、T15(p<0.001)、T16(p<0.001)之间差异显著;T11与T12(p=0.002)、T13(p=0.001)、T14(p<0.001)、T15(p<0.001)、T16(p<0.001)差异均显著;T12与T13(p=0.001)、T14(p<0.001)、T15(p<0.001)、T16(p=0.001)差异均显著;T13与T14(p=0.001)、T15(p=0.002)、T16(p=0.006)差异均显著;T14与T15(p=0.012)差异显著。其他时间窗口之间(T14与T16、T15与T16)差异不显著。θ节律同步化活动在时间窗内波幅变化趋势如图4 所示。
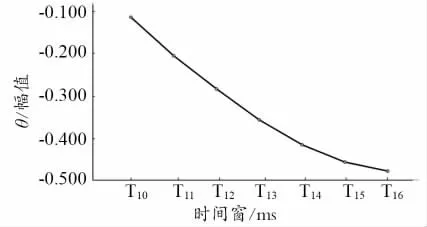
图4 θ 频率同步化值随时间窗的变化
以上结果表明,α和θ节律的波幅变化随时间窗口的变化整体呈现下降趋势,并且时间窗的成对比较分析显示,α和θ节律的波幅变化在视频播放的前期时间点与视频播放的后期时间点之间存在差异,说明运动员随着视频观看时间的累积,α和θ节律的波幅变化更加显著。
2 乒乓球运动员动作预测的大脑活动空间特征
2.1 研究目的与假设
1)探究运动经验是否导致AON 主要脑区的激活及AON 在动作预测中的作用。2)研究假设:运动员在进行预测任务时AON 的激活加强,功能连接增强。
2.2 研究设计
采用3(遮挡条件:早期、中期、晚期)×2(组别:专家组、新手组)两因素混合设计。其中组别为被试间变量,遮挡条件为被试内变量,因变量为反应正确率和fMRI 任务中激活脑区。
2.2.1 被试
共招募了41 名上海体育大学乒乓球运动员(以下以“专家”指代)与普通大学生(以下以“新手”指代)。其中有3 名被试因在功能性磁共振(fMRI)扫描过程中头部移动过大被排除,另外1 名被试在磁共振扫描过程中因身体不适终止了实验,最终37 名被试的数据纳入分析,其中:专家组为18 人(国家一级乒乓球运动员和国家二级乒乓球运动员)、新手组为19 人(无运动训练经验的普通大学生)。所有被试裸眼视力或矫正视力正常,均为右利手,无神经系统疾病和精神障碍史。被试在完成实验后会获得一定的实验报酬。
2.2.2 实验材料
实验刺激材料为自行录制的乒乓球运动员发球视频,发球者为2 名发球较好的乒乓球运动员(男生1人,女生1 人)。男生身着蓝色T 恤,女生身着红色T恤,并且在实验开始前告知被试,方便被试轻松辨认。发球者使用反手发球,球的落点位置在左右球台各半、远近球台各半。截断时间分为3 种条件。1)早期遮挡。视频最后一帧为球与球拍接触前一帧(视频长度800 ms)。2)中期遮挡。视频最后一帧为球与球拍接触(视频长度900 ms)。3)晚期遮挡。视频最后一帧为球与球拍接触后一帧(视频长度1 000 ms)。
2.2.3 实验程序
在正式实验开始前,被试需要签署知情同意书并且被告知有中途退出的权力。在进入磁共振扫描室前,告知被试注意事项和实验任务,并在磁共振扫描室外进行练习,确保被试完全理解实验任务,练习完成后,让被试进入磁共振扫描仪进行正式实验。本研究使用软件“E-prime”制作实验程序并呈现视频刺激。扫描过程中被试可以通过安装在头部线圈上的反射镜观看视频刺激,通过与磁共振扫描仪兼容的反应盒进行按键反应。反应盒一共有5 个键,对应右手的5个手指,本实验中被试只需要用到食指(对应1 键)和中指(对应2 键)。实验程序首先呈现准备界面,扫描开始后首先呈现8 s 的倒计时,然后开始正式实验任务。
首先呈现任务类型(落点判断和性别判断)5000ms,然后呈现“+”注视点1 000 ms,注视点消失后呈现视频刺激800 ms,视频播放完后,屏幕立即变成黑屏,被试有4 500~6 000 ms(试次间抖动变化)的时间用右手的食指和中指按键作出反应。要求被试在不牺牲正确率的情况下尽快作出反应。在落点判断任务中,判断球落在左边按1 键,落在右边按2 键;在性别判断任务中,判断为男性按1 键,判断为女性按2 键(如图5所示)。实验过程中不呈现结果反馈。

图5 实验流程
实验程序的设计采用组块模式,正式实验中每个被试进行4 组实验(4 个序列),每组实验中有4 个组块,分别对应4 种视频刺激(落点判断任务:早期遮挡、中期遮挡、晚期遮挡;性别判断任务),每个组块有16 个视频刺激(男女被试各半),共256 个视频刺激。4个组块的顺序在被试内进行平衡。每组实验完成后,被试可以选择休息,然后再开始下一组实验。
2.2.4 磁共振扫描程序与参数设定
本研究使用上海体育大学脑成像实验中心的西门子“3T MAGNETOMPrisma”磁共振成像系统进行数据采集,实验过程使用了20 通道头部线圈。磁共振扫描序列包括了定位像、场图、3 组功能像扫描和一个结构像扫描,扫描时长大约为40~50 min。
功能态脑成像数据的采集使用平面回波成像序列(缩写为EPI)。具体参数如下:扫描视野=224×224 mm2,重复时间=2 000 ms,回波时间=30 ms,翻转角=90°,体素大小=3.5×3.5×3.5 mm3,层厚=3.5 mm,层数为33 层,每个任务序列包含218 张全脑扫描图像。
结构像数据的采集使用快速梯度回波序列(缩写为MPRAGE),具体参数如下:扫描视野=256×256 mm2,重复时间=2 530 ms,回波时间=2.98 ms,翻转角=7°,体素大小=1×1×1 mm3,层厚=1 mm,层数为192 层。
2.2.5 磁共振数据激活分析
磁共振数据分析使用基于“Matlab2018a”的SPM12(http://www.fil.ion.ucl.ac.uk/spm)工具包完成,具体分析步骤如下。
1)预处理。具体包括:数据格式转换;删去功能像前8 s 的数据(前4 张图片)以保证扫描性能稳定性;时间层矫正;头动矫正,剔除头动大于3 mm 或转动超过3°的被试数据;结构像配准;空间标准化,将功能像配准到加拿大蒙特利尔神经学研究所(缩写为MNI)标准化空间;空间平滑,将标准化图像采用半高全宽(缩写为FWHM)调整为8×8×8 mm3的平滑核完成高斯卷积。
2)个体分析。通过一般线性模型(GLM)对被试的整个脑区激活情况进行统计分析,界定知觉预期任务中的一般认知脑区。根据实验条件设置每个试次的开始时间和持续时间,并剔除反应错误的试次,6 个头动参数作为协变量,将这些变量与标准血液动力学响应函数(缩写为HRF)进行卷积。该实验中每个扫描序列的矩阵有10 列,分别为落点判断任务的早期遮挡、中期遮挡、晚期遮挡、性别判断任务及6 列头动参数。基于上述设计矩阵构造出多组线性对照作为实验结果的条件:早期遮挡条件与性别判断任务;中期遮挡条件与性别判断任务;晚期遮挡条件与性别判断任务。此外,还将3 种遮挡条件的落点判断任务进行合并分析。
3)组分析。分为2 个部分:首先对2 个组的被试分别进行单样本t检验,探究预期任务是否会激活AON的相关脑区;其次对2 组被试进行双样本t检验,探究组别效应。
4)ROI分析。ROI 选取了双侧额下回(缩写为IFG)、双侧顶下小叶(缩写为IPL)、左侧额上回(缩写为SFG)、左侧颞上回(缩写为STG)、双侧颞上沟(缩写为STS)和双侧背侧前运动皮层(缩写为dPMC)。ROI 的分析用于探究专业知识是否会导致AON 主要脑区的激活差异及AON 在动作预测中的作用。
5)功能连接分析。同样基于选定的ROI,对选定的ROI 进行了ROI-to-ROI 的功能连接的分析,将选定的ROI 进行两两相关分析,相关系数显著,说明这2 个ROI 之间存在有效连接,然后对2 组被试共同存在的有效连接进行相关系数的差异性检验,探究2 组被试的功能连接差异脑区,验证运动经验是否会导致AON 相关脑区的功能的适应性改变。
2.3 实验结果
2.3.1 行为结果
反应时采用3(遮挡条件:早期、中期、晚期)×2(组别:专家组、新手组)的两因素重复测量方差分析进行检验。
行为数据如表4 所示。落点判断任务的结果分析采用3(遮挡条件)×2(组别)的重复测量方差分析,结果发现组别主效应显著[F(1,35)=35.848,p<0.001,η2=0.506],专家组的正确率显著高于新手组;遮挡条件主效应显著[F(2,70)=126.723,p<0.001,η2=0.784],晚期遮挡条件的正确率高于早期遮挡和中期遮挡条件;组别与遮挡条件的交互作用显著[F(2,70)=9.249,p<0.001,η2=0.209]。性别判断任务2 组差异不显著,且均高于90%。
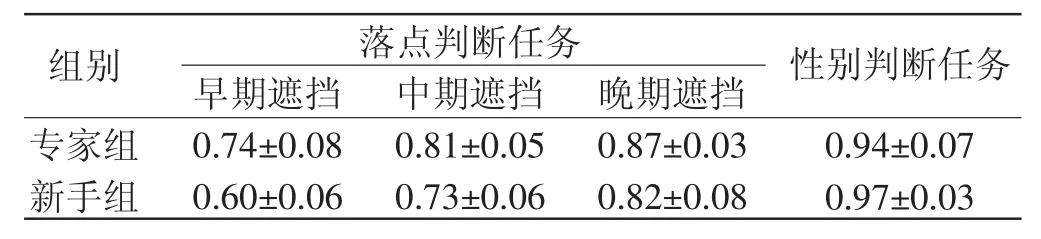
表4 2 组任务判断正确率(平均数±标准差)
进一步简单效应分析表明:在早期遮挡条件下,专家组的正确率显著高于新手组(p<0.001,95%CI[0.099,0.191]);在中期遮挡条件下,专家组的正确率显著高于新手组(p<0.001,95%CI[0.052,0.127]);在晚期遮挡条件下2 组反应正确率差异无统计学意义。专家组在早期遮挡条件下的正确率显著低于中期遮挡条件下(p<0.001,95%CI[-0.116,-0.032])和晚期遮挡条件下的正确率(p<0.001,95%CI[-0.165,-0.087]),而中期遮挡条件和晚期遮挡条件下的正确率差异无统计学意义;新手组在早期遮挡条件下的正确率显著低于中期遮挡条件下(p<0.001,95%CI[-0.170,-0.088])和晚期遮挡条件下的正确率(p<0.001,95%CI[-0.257,-0.181]),中期遮挡条件下的正确率显著低于晚期遮挡条件下的正确率(p<0.001,95%CI[-0.125,-0.055])。结果如图6 所示。
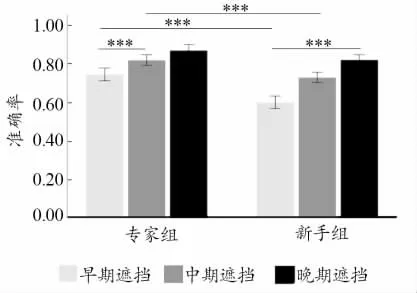
图6 2 组被试在不同遮挡条件下的正确率
2.3.2 AON 参与动作预测的脑区激活研究结果
2.3.2.1 预测任务激活脑区特点
在动作预测任务中,2 组被试的激活脑区结果显示,2 组被试均激活了左侧枕中回(缩写为MOG)、右侧颞中回(缩写为MTG)及右侧额中回(缩写为MFG),与新手组相比,专家组还存在右侧顶下小叶(缩写为IPL)的激活,而新手组则额外存在左侧中央后回(缩写为PoCG)的激活,见表5 并如图7 所示。
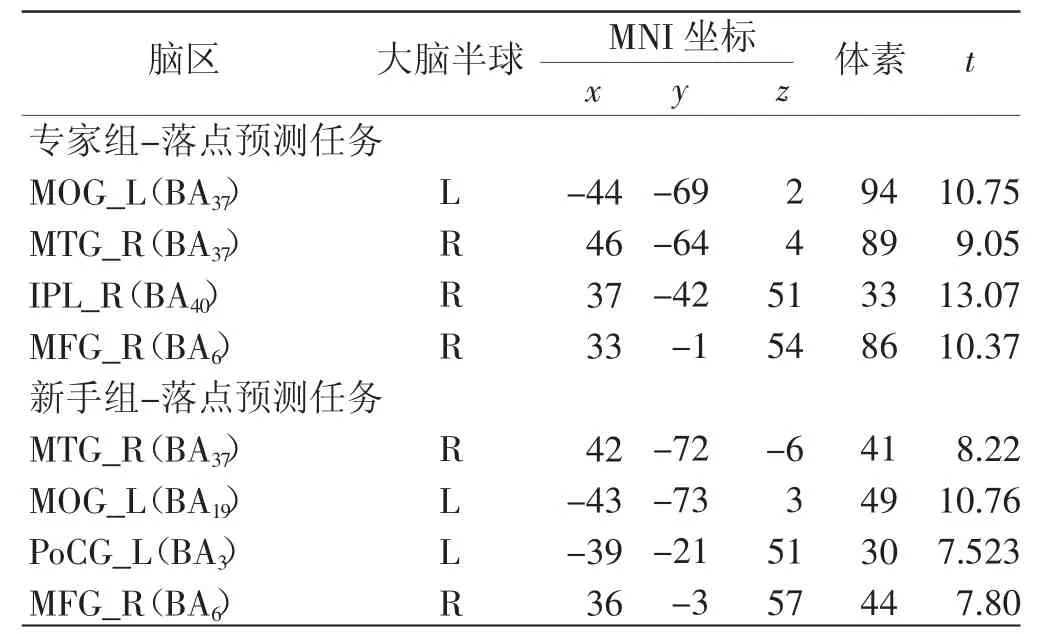
表5 各组落点预测任务激活脑区

图7 落点预测任务脑区激活情况
2.3.2.2 运动经验影响大脑激活的结果
在早期遮挡条件下,2 组被试未见显著差异;在中期遮挡条件下,2 组被试激活差异显著的脑区较多,表现为与新手组相比,专家组在MOG_L、IPL_R、PCG_L,SFG_L 和MFG 表现出更强的激活;在晚期遮挡条件下,与新手组相比,专家组在PCG_L、STG_R 有着更强的激活水平。
2.3.3 ROI 分析
基于以上结果进行分析可知,在中期遮挡条件下,2 组激活水平存在差异的脑区最多(见表6)。因此,在ROI 分析中只选取了中期遮挡条件进一步进行讨论。
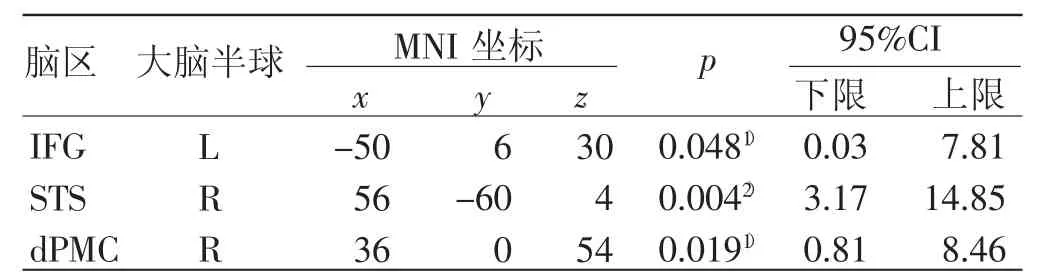
表6 脑区激活差异ROI 分析结果
对ROI 主要脑节点的激活分析发现,专家组在3个脑区(即IFG_L、STS_R、dPMC_R)的激活水平均显著高于新手组。
2.3.4 功能连接分析
从功能连接分析结果(如图8 所示)可以发现,专家组经ROI 分析的脑区间普遍存在着有效连接,且连接强度大,而新手组经ROI 分析的脑区间的有效连接较少,且连接强度较弱。

图8 2 组功能连接强度热力情况
专家组的IFG_L 与双侧IPL、STS_R,双侧IFG 与dPMC_L,双侧IPL 与STS、dMPC,SFG_L 和STG_L 之间有着更强的功能连接,而新手组则不存在以上脑区间的功能连接(见表7 和表8)。
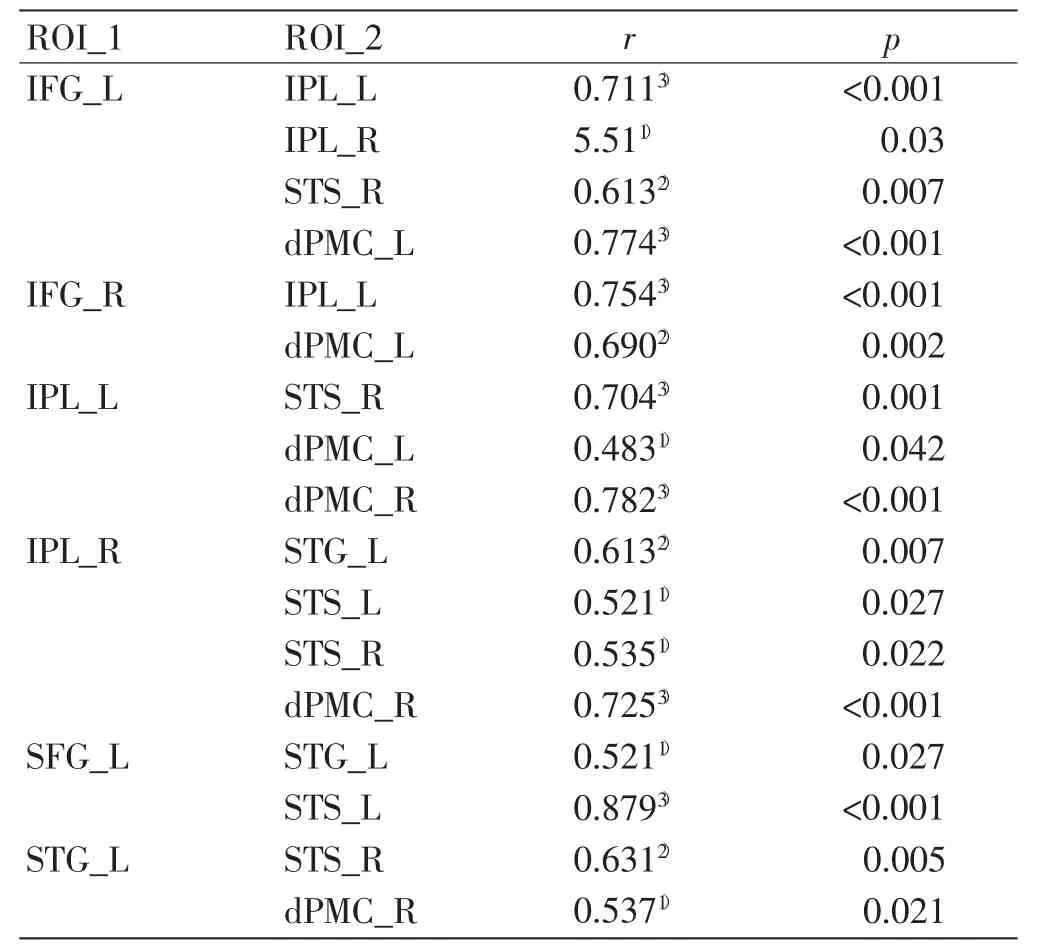
表7 专家组相关性显著脑区

表8 新手组相关性显著脑区
对2 组相关性均显著的脑区进行了相关系数的差异性检验,结果发现与新手组相比,专家组的SFG_L 与dPMC_R、IFG_L 功能连接更强,见表9。

表9 2 组相关系数均显著脑区比较
3 讨论
3.1 乒乓球运动员的动作预测的大脑活动时间与空间特征
乒乓球运动员动作预测的时间特征结果显示,α节律活动值随运动员动作预测的时间进程下降,而α节律的去同步化活动与大脑皮层唤醒度呈负相关关系。由α 同步值的变化曲线可知,在视频播放前期(动作刚开始进行时),被试的大脑平层兴奋性相对较弱,随着动作的进行,α 去同步化增大,被试大脑皮层兴奋性增强,说明在对手做动作前期,被试的关注度并不高,而随着动作的进行,特别是在球与球拍接触前的时间点,被试大脑皮层兴奋性处于最高点,投入的认知资源最多。因此,在动作预测之前的动作观察阶段,大脑皮层兴奋性的增加可能反映了运动员对动作模式识别的自下而上的注意力加工方式。
θ节律活动值随运动员动作预测的时间进程降低。而θ节律的去同步化与记忆加工和认知控制需求呈正相关。由θ同步值的变化曲线可知,在视频播放前期(动作刚开始进行时),被试对专项动作的信息编码和记忆加工需求最高,但随着运动信息的增加,θ节律活动的降低可能反映了运动员在处理动作信息时认知需求的降低。对此结果,结合前人研究作出2 方面推测。第一,由于刺激材料为仅1 000 ms 的短视频刺激材料,被试对看清发球动作和球飞行轨迹的刺激需求更高,这时θ对认知控制的高级功能不再占据主导,反而根据视觉输入的强度而将自上而下的认知控制调整为由刺激主导的自下而上的动作加工[10];第二,被试在逐渐适应了任务要求后,唤起了对专项运动相关记忆的加工,而被试在长期训练之下,其长时记忆系统存储有关发球动作特征模型,根据这些概括化的动作模型,只需少量的动作特征信息就可完成识别任务,因此表现出其注意负荷降低,动作特征信息检索、编码和匹配有效性提高[21]。
从乒乓球运动员的动作预测的空间特征分析结果发现,通过对比乒乓球专家和新手2 组的激活差异脑区发现,乒乓球专家组在IPL、IFG、MFG、STS、MOG及dPMC 有着更强的激活水平,这都是AON 的主要脑区,再次证明AON 的确可以作为动作预测的神经基础。通过ROI 分析也发现,专家组在AON 的主要节点,例如IFG、IPL、STS/pMTG、dPMC 有着更强的激活状态;通过功能连接分析发现,乒乓球专家组的AON各脑区节点之间的功能连接普遍更强,额顶脑区和颞叶相关脑区之间均有显著连接,而乒乓球新手组仅在STG 与IFG、左侧IPL 表现出更强的连接。因此,可以证明AON 的激活强度和功能连接强度受到运动经验的调节,也就是说运动经验导致了AON 相关脑区功能的适应性改变。
3.2 乒乓球运动员动作预测的大脑活动时空整合特征
在对专家动作预测的时空特征整合的研究中发现:α节律、θ节律和AON 中的较初级脑区和较高级脑区本身具有的功能之间存在交互关系。
首先,对AON 的关键组成部分[22](即额顶系统)功能的大量探讨已经发现,运动员的预测优势可能存在于动作编码和匹配,这依赖于运动员在长期训练中形成的大量且准确的动作表征,他们在看到发球动作时就能够根据已有经验进行准确的动作模拟,为动作观察提供预测推理[23],从而作出正确预测。从阻断任务范式的研究中发现,专家相较于新手更依赖顶叶功能[24-25],专家由于视觉信息有限而采取了更优化的注意力策略[26],而顶叶θ已被证明能够提高注意力资源的招募效率[10],更好地整合自上而下和自下而上的加工[12]。本研究中θ节律的去同步化结果趋势可能就反映了针对目标朝向而调整的注意功能。
从前人关于枕颞脑区的研究中发现,枕颞脑区共同参与对运动信息的处理,例如对手的身体信息等[27]。在当前研究中发现,专家组在pSTS/pMTG 和MOG 都有显著激活。功能连接的分析结果显示,运动员的双测STS 和额顶的主要脑区如IPL、IFG 及SFG 存在有效连接。脑区间功能连通性的增强得益于乒乓球运动员个体长期的运动训练,从而形成了更高效的视觉信息整合策略,例如在一般视觉-空间任务中,乒乓球运动员的顶-枕、顶-额α节律相干性更高,反映了运动员的一般注意唤醒与任务特定注意加工相关[28]。同时,长期运动训练还会导致认知加工熟练化和自动化[29],表现为高级认知控制皮层的能量消耗降低,节约中枢认知资源,提高了运动员的行为绩效[21,30]。例如在专项动作识别任务中,乒乓球选手的左侧颞-额θ节律相干性更低[31]。有研究发现,当额外增加的观察负荷导致动作观察能利用的注意资源显著减少时,IFG节点附近的大脑活动就会减少[32-33],这说明当注意力资源的分配阻碍了对动作观察的编码时,自上而下的认知控制减弱(θ节律活动减少),对注意力资源的分配可能会更多倾斜给动作刺激本身的加工(α节律活动增多)。
综合EEG 和fMRI 的研究结果推测,当乒乓球运动员根据运动信息动作观察并作出动作预测时,从时间特征上,乒乓球运动员首先激活了对动作观察的高度唤醒(α节律去同步化活动),然后运动经验优化了他们对认知资源的控制与分配(θ节律同步化活动)。在乒乓球运动员识别到动作模式并从中提取出有效预测线索的认知加工过程中,AON 网络得到激活,开始对输入的刺激信息进行感知加工,并最终输出为对动作结果的预判。
4 结论
运动信息是影响动作预测表现的主要变量之一。随着运动信息量的增加,乒乓球运动员的动作预测正确率及注意力唤醒度提高,认知控制投入减少。运动训练经验是知觉预期过程中AON 重要节点脑区激活和功能连接的主要因素。α、θ节律与AON 重要节点脑区在功能上的整合,是动作预测在时空特征上反映大脑网络连接的基础。
