放牧强度对高寒草甸优势植物叶片解剖结构的影响
2023-11-02林轸荣姜风岩石丽娜赵洪鑫李永元祁进贤党志英董全民周华坤邵新庆
肖 晴,林轸荣,姜风岩,石丽娜,赵洪鑫,李永元,祁进贤,党志英,董全民,周华坤,邵新庆*
(1.中国农业大学草业科学与技术学院,北京 100193;2.青海省饲草料技术推广站,青海 西宁 810000;3.青海省祁连县野牛沟林场,青海 海北藏族自治州 810499;4.青海省祁连县林场,青海 海北藏族自治州 810499;5.青海大学畜牧兽医科学院,青海省畜牧兽医科学院,青海省高寒草地适应性管理重点实验室,青海 西宁 810016;6.中国科学院西北高原生物研究所,青海省寒区恢复生态学重点实验室,青海 西宁 810001)
青藏高原是我国主要的牧区之一,草地类型丰富,高寒草地面积占我国草地总面积的44%[1]。近年来,由于气候变化和超载过牧等原因,青藏高原高寒草甸出现了不同程度的退化现象[2],严重干扰着草地生态系统的稳定性和生态平衡。放牧是草地利用的主要方式之一[3]。在长期的放牧过程中,放牧家畜和草地植物之间存在着复杂的平衡关系。放牧导致的环境变异使植物在长期放牧过程中产生多种适应性进化机制[4],以保护其与非生物环境和放牧家畜的协调共存。放牧会改变草原植物群落的宏观生理状态,如高度[5]、丰富度[6]和生物量[7]等,而且对植物个体生长和形态特征产生不可忽略的影响[8,9]。
叶片是植物个体在生长发育过程中的重要器官,其形态和结构上往往存在变异性和可塑性[10]。以放牧为代表的人类活动不仅会通过放牧家畜的采食直接影响植株的器官和生长进程,还会改变植物生境中的关键环境因子(如温度、土壤含水量、光照条件等),影响叶片的生长发育[11]。有关放牧强度对植物叶片的相关研究结果显示,放牧强度的增加会导致短花针茅的叶面积表现出“小化”[12],并降低高原早熟禾叶片的木质部厚度[13];过度放牧导致部分植株叶角质层厚度增厚[14],角质层蜡质的组分与含量发生适应性变化[15]。长期放牧甚至会影响基因表达的可塑性,影响植物生理代谢途径[16]。放牧压力下植物所表现出的植物叶肉细胞密度[17]等形态特征的变化,反映了植物在异质生境下的适应性,即植物通过对可利用资源的重新分配[18],优化分配格局[19]让部分器官可以获得优先发育权[20],最终达到稳定植物体正常生长发育进程的目的。因此,研究植物叶片在放牧干扰下的解剖结构变化,更能反映植物对环境变化的差异性和可塑性。矮生嵩草(Kobresiahumilis)属于莎草科嵩草属多年生密丛型嵩草,紫花针茅(Stipapurpurea)属于禾本科针茅属多年生密丛禾草,二者作为高寒草甸的主要优势植物,均具有良好的耐寒性、耐旱性和耐牧性等特性,是高寒草甸的主要建群种[21,22]。经过长期不同强度的放牧后,优势植物除了在形态[3]和繁殖策略[23]等方面适应放牧外,叶解剖结构是否也受放牧强度影响尚不可知。
本研究以不同放牧强度下连续6年在生长季内(6—10月)持续放牧的两种主要植物(即紫花针茅和矮生嵩草)为研究对象,通过分析不同放牧强度下叶片解剖结构的变化从微观角度揭示植物对放牧干扰的响应,探讨植物对放牧干扰产生的可塑性变化,这将有助于进一步揭示高寒草甸植物对放牧的适应机理,以期为植物微观结构对放牧适应机制的进一步研究提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于青海省北州海晏县西海镇(36°44′~37°39′N,100°23′~101°20′E),平均海拔3 000~3 100 m。该地属高原大陆性气候,降水多集中在5—9月年平均降雨量为400 mm,年均温为1.5℃,年日照时间为2 580~2 750 h。一年之中四季变化不明显,仅有冷暖季之别[24]。研究区土壤类型为高山草甸土,草地类型为高寒草甸。样地内植物群落主要优势种为矮生嵩草(Kobresiahumilis)和紫花针茅(Stipapurpurea),次优势种主要为干生薹草(Carexaridula)、星毛委陵菜(Potentillaacaulis)和草地早熟禾(Poapratensis)等。
1.2 放牧试验设计
为控制本底和空间异质差异,本研究选择地势相对平坦、环境相对均匀的同一块草地布设样地。该放牧试验采用完全随机区组设计,共设置不放牧(Control,CK)、轻度放牧(Light grazing,LG)、中度放牧(Moderate grazing,MG)和重度放牧(Heavy grazing,HG)4个处理,每个处理设置3个重复。轻度放牧为3只·hm-2,牧草利用率为30%~35%,中度放牧为4只·hm-2,牧草利用率为50%~55%,重度放牧为5只·hm-2,牧草利用率为65%~70%。试验中放牧家畜选择体况相近的1岁左右公藏羊,体重为(30±2) kg,每只藏羊为1个羊单位。各处理的家畜数量和小区面积见表1。长期的放牧试验平台自2014年延续至今,于每年的6月份开始放牧,10月底结束放牧。放牧期间不进行补饲,在样地内固定位置放置饮水槽,每隔2天加水1次,确保藏羊饮水充足[25]。

表1 放牧强度试验设计Table 1 Experimental design of grazing intensity
1.3 取样与测定
1.3.1试验材料获取 2021年8月下旬,分别在不同放牧强度处理小区内采集2种优势种矮生嵩草(Kobresiahumilis)和紫花针茅(Stipapurpurea)叶片,每个小区内的2种植物分别随机选取5丛植株,记为5个重复。选择自上而下第1个完全展开的健康叶片,用卡尺测定植物叶片长度,记为植物叶长。取样时,用剪刀剪取叶片靠近叶基部(标记为“B”)以及靠近叶尖端(标记为“T”)0.5~1.0 cm的叶段,分别记录该叶段中点距叶尖的距离。将剪取的植物叶片立即放入FAA固定液(福尔马林-70%酒精-醋酸混合液,体积比为5∶90∶5)中进行固定。将FAA固定液固定24 h后的植物叶片依次移入浓度为70%,85%,95%和100%酒精中(各2 h)进行脱水,并依次用100%纯酒精和二甲苯混合液(1∶1,2 h)和纯二甲苯(2 h)将叶片材料透明化,再用熔点为52℃~54℃的石蜡进行包埋。用连续切片机(Leica RM2235,Leica Inc.,Wetzler,Germany)切片,经苏木精伊红染色后制成永久切片。永久切片中叶横切面用连接数码相机(SPOT Insight,Diagnostic Instruments Inc.,Stirling Heights,MI,USA)的显微镜(Olympus CX21,Olympus Inc.,Tokyo,Japan)观察并拍照获取图像,用Image-Pro Plus软件进行显微结构测定[26]。
1.3.2指标测定与方法 选取3~4个连续且完整的叶切片主脉处的叶片厚度均值记为该叶段切片的叶厚(Leaf thickness,μm)。选取3~4个连续且完整的叶切片主脉处同一部位的近轴面角质层、近轴面表皮细胞、远轴面角质层厚和远轴面表皮细胞的厚度均值记为该叶段切片的近轴面角质层厚(Adaxial cuticular thickness,μm)、近轴面表皮细胞厚(Adaxial epidermis thickness,μm)、远轴面角质层厚(Abaxial cuticular thickness,μm)和远轴面表皮细胞厚(Abaxial epidermis thickness,μm)。将同一个切片所测得的叶厚减去角质层厚以及表皮细胞厚所得到的厚度记为该切片的叶肉细胞厚,选取3~4个连续且完整的叶切片的叶肉细胞厚均值,记为该叶段切片的叶肉细胞厚(Mesophyll cell thickness,μm)。每个叶切片的最大导管直径为视野中主脉木质部处的两个大导管的直径均值,最小导管直径为视野中主脉木质部处2~3个小导管的直径均值。选取3~4个连续且完整的叶切片的最大导管直径与最小导管直径的均值记为该叶段切片的最大导管直径(Maximum xylem vessel diameter,μm)与最小导管直径(Minimum xylem vessel diameter,μm)。选取3~4个连续且完整的叶切片主脉处的木质部以及韧皮部的面积均值记为该叶段切片的木质部面积(Xylem transection area,μm2)与韧皮部面积(Phloem transection area,μm2)。

图1 测定指标示意图Fig.1 Schematic diagram of the measured indicators注:1,近轴面角质层;2,近轴面表皮细胞;3,远轴面角质层;4,远轴面表皮细胞;5,叶肉细胞组织厚;6,叶厚;7,木质部;8,韧皮部Note:1,Adaxial cuticle;2,Adaxial epidermal cells;3,Abaxial cuticle;4,Abaxial epidermal cells;5,Mesophyll cell;6,Leaf thickness;7,Xylem;8,Phloem
1.4 数据处理
采用Microsoft excel 2016整理野外调查和室内测定所得数据。在数据分析前使用R软件中的Stats包的shapiro.test函数进行Shapiro-Wilk正态分布检验,使用car包的leveneTest函数实现Levene方差齐性检验。采用单因素Kruskal-Wallis非参数检验对不同放牧强度下的同一取样部位进行组间比较,当总体组间有统计学差异时,进一步使用FSA包的dunnTest函数进行Dunn test多重比较,P值校正方法选择bonferroni法。同一放牧强度下的不同取样部位间差异用wilcox秩和检验,采用Stats包的wilcox.test函数进行两组间非参数差异分析。使用rcompanion包的scheirerRayHare函数对取样部位和放牧强度进行双因素Kruskal-Wallis非参数检验。显著性水平设置为P<0.05。以上统计分析均在R 4.2.0(R Core Team,2021)中完成,图表中数据为平均值±标准误。
2 结果与分析
2.1 放牧强度对叶形态结构的影响
中度放牧下,矮生嵩草的叶长较不放牧处理下显著减小了26.44%(图2C,P<0.05);重度放牧下,紫花针茅和矮生嵩草的叶长较不放牧处理均表现出不同程度的减小,分别降低20.90%和22.89%,达到显著水平(图2A,C,P<0.05)。
2.2 放牧强度对叶解剖结构的影响
2.2.1放牧强度对叶光合组织的影响 随着放牧强度的增加,紫花针茅叶基部的叶肉细胞厚逐渐降低;而矮生嵩草叶基部的叶肉细胞厚随着放牧强度的增大而逐渐增厚(图3)。除重度放牧外,紫花针茅的叶肉细胞厚在不同取样部位之间均存在显著差异(P<0.05)。矮生嵩草的叶肉细胞厚在取样部位与放牧强度下均无显著差异。
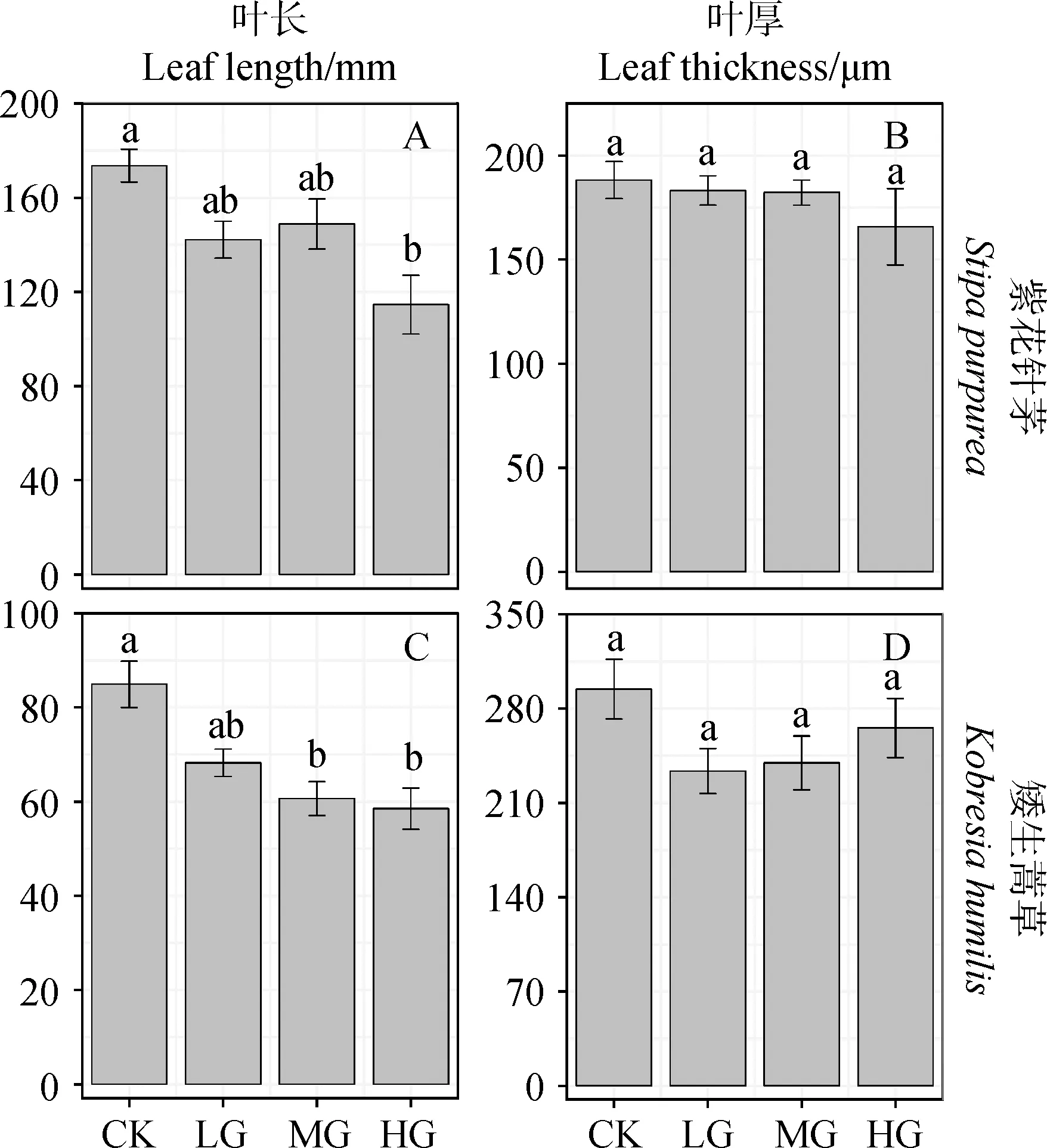
图2 叶长和叶厚对放牧强度的响应Fig.2 Response of leaf length and leaf thickness to grazing intensities注:图中为不同物种叶长(A,C;mm)、叶厚(B,D;μm)对放牧强度的响应;CK,对照;LG,轻度放牧;MG,中度放牧;HG,重度放牧;小写字母分别表示在不同放牧强度之间的显著性差异Note:The panels display the response of Leaf length (Panel A,C;mm) and Leaf thickness (Panel B,D;μm) to grazing intensities.CK,Control;LG,Light grazing;MG,Moderate grazing;HG,Heavy grazing.The different lowercase letters indicate a significant difference between the different grazing intensities

图3 叶肉细胞厚对放牧强度的响应Fig.3 Response of mesophyll tissue cell thickness to grazing intensities注:图中为不同物种叶肉细胞厚(A,B;μm)对放牧强度的响应;B,叶片基部位置;T,叶片尖端位置;CK,对照;LG,轻度放牧;MG,中度放牧;HG,重度放牧;大写字母表示同一放牧强度下不同取样部位之间的显著性差异,小写字母分别表示在同一取样部位下不同放牧强度之间的显著性差异。下同Note:The panels display the response of Mesophyll tissue cell thickness (panel A,B;μm0 to grazing intensities.B,Base of the leaf;M,Middle of the leaf;T,Tip of the leaf.CK,Control;LG,Light grazing;MG,Moderate grazing;HG,Heavy grazing.The different uppercase letters indicate a significant difference between different sampling sites under the same grazing intensity,and the different lowercase letters a significant difference between different grazing intensities in the same sampling site.The same as below
2.2.2放牧强度对叶保护组织的影响 不放牧处理下,紫花针茅叶尖端的近轴面角质层厚、近轴面表皮细胞厚以及远轴面角质层厚均显著大于叶基部(图4A-C,P<0.05),随着放牧强度的增大,不同解剖结构在不同取样部位间的厚度关系受到干扰:仅重度放牧处理下,叶尖端的近轴面角质层厚显著大于叶基部(图4A,P<0.05);放牧干扰下两部位间虽仍表现为叶尖端的近轴面表皮细胞厚高于基部,但差异缩小(图4B);仅中度放牧下叶尖端的远轴面角质层厚显著大于叶基部(图4C,P<0.05)。
对矮生嵩草的叶保护组织部分解剖结构进行分析发现,放牧强度和取样部位对其近轴面角质层厚、近轴面表皮细胞厚以及远轴面表皮细胞厚存在显著影响。与不放牧处理相比,矮生嵩草叶尖端的远轴面表皮细胞厚在中度放牧处理下减小了18.61%,且差异显著(图4H,P<0.05);其近轴面角质层厚在重度放牧处理下表现出显著增厚(图4E,P<0.05)。矮生嵩草叶尖端的近轴面表皮细胞厚在中度、重度放牧下较轻度放牧处理表现为显著增大(图4F,P<0.05)。不同取样部位下的近轴面角质层厚在不放牧和轻度放牧下均存在显著差异(图4E,P<0.05);除轻度放牧处理外,其余处理条件下矮生嵩草不同取样部位之间的近轴面表皮细胞厚均存在显著差异,并且在中度放牧下二者差异达到最大,其顶端叶段高出基端叶段83.99%(图4F,P<0.05);仅在中度放牧下,叶基端的远轴面表皮细胞厚显著大于叶尖端(图4H,P<0.05)。

图4 近轴面角质层厚、近轴面表皮细胞厚、远轴面角质层厚和远轴面表皮细胞厚对放牧强度的响应Fig.4 Response of adaxial cuticular thickness,adaxial epidermal cells thickness,abaxial cuticular thickness and abaxial epidermal cells thickness to grazing intensities
2.2.3放牧强度对维管组织的影响 除最小导管直径外,紫花针茅叶尖端和叶基部间的叶片最大导管直径、韧皮部横截面积和木质部横截面积均存在显著性差异(图5A-D,P<0.05)。矮生嵩草叶基部最大导管直径在中度和重度放牧处理下较不放牧处理显著分别降低了19.44%和17.16%(图5E,P<0.05),但对叶尖端的最大导管直径无显著影响(图5E)。在不放牧、轻度放牧和中度放牧处理下,矮生嵩草叶基部的最小导管直径显著高于顶端叶段,分别高出42.74%,40.42%和26.65%;重度放牧处理下矮生嵩草叶尖端与叶基部间的最小导管直径则无显著差异(图5F)。矮生嵩草叶基部和叶尖端间的韧皮部横截面积以及木质部横截面积在不同放牧强度处理下均表现出显著差异(P<0.05)。
2.3 取样部位和放牧强度对不同物种叶解剖结构的影响
取样部位和放牧强度对矮生嵩草的近轴面表皮细胞厚均具有显著影响(P<0.05),且取样部位对近轴面表皮细胞厚的影响大于放牧强度(表2)。取样部位对矮生嵩草的近轴面角质层厚、木质部横截面积、韧皮部横截面积、最大导管直径以及最小导管直径均具有极显著影响(P<0.001)。放牧强度与取样部位的交互作用对矮生嵩草的远轴面表皮细胞厚有显著影响(P<0.05)。取样部位对紫花针茅叶肉细胞厚、近轴面角质层厚、近轴面表皮细胞厚、远轴面角质层厚、木质部横截面积、韧皮部横截面积及最大导管直径存在极显著影响(P<0.001)。

图5 最大导管直径、最小导管直径、韧皮部横截面积和木质部横截面积对放牧强度的响应Fig.5 Response of maximum xylem vessel diameter,minimum xylem vessel diameter,phloem transection area,and xylem transection area to grazing intensities

表2 取样部位和放牧强度对矮生嵩草和紫花针茅叶解剖结构的影响Table 2 Effects of different sampling sites and grazing intensities on the leaf anatomical traits of Kobresia humilis and Stipa purpurea
3 讨论
3.1 放牧强度增大促使高寒草甸矮生嵩草和紫花针茅的叶形态结构趋向“小型化”
植物叶片直接暴露于环境之中,往往对环境变化非常敏感,植物对环境变化的响应往往最先由叶片表现出来[27],叶形态解剖结构则能够很好地反映植物对环境变化的适应能力[28]。本研究中,中度放牧与重度放牧下均出现叶长的显著减小,这说明在放牧干扰下,家畜对植物的过度采食会导致叶片变短,这与李江文等[12,29]的研究结果一致。叶厚可以作为植物抗逆性的反映指标,通常情况下抗逆性较强的植物叶厚较厚[30]。本研究中两种植物的叶厚在不同的放牧强度均无显著性差异,表明紫花针茅和矮生嵩草适应放牧的能力较强,与其耐牧性强的特性相吻合。
3.2 放牧下矮生嵩草和紫花针茅的叶解剖结构产生适应性变化
角质层及其蜡质是将水分散失限制在不受植物短期控制的安全值屏障,在控制CO2内流和水分外流中发挥关键作用[31],在抵御环境胁迫中起着重要作用。矮生嵩草叶尖端在中度放牧和重度放牧下较不放牧处理均表现为近轴面角质层增厚,并在重度放牧下达到显著水平,叶基部的远轴面角质层厚随着放牧强度的逐步加强而渐渐增厚。这可能是矮生嵩草应对放牧干扰下的一种避牧策略,植物叶片角质层厚度与纤维素的含量呈正相关关系[32],而纤维素含量的增加会降低植物的适口性[27],从而减少被家畜啃食的概率。在重度放牧下,矮生嵩草的近轴面表皮细胞厚较不放牧处理出现增厚现象,可能是由于过度放牧下草地出现一定程度的退化,致使地表裸露面积增大、土壤水分蒸发加快,矮生嵩草为应对水分缺失的环境而表现出表皮细胞的增厚。对紫花针茅而言,除中度放牧下叶基部的近轴面角质层厚较不放牧处理表现为小幅度增大外,其余放牧条件下的角质层厚均出现减小现象。除此之外,紫花针茅的表皮细胞厚也表现出相似情况,说明紫花针茅的叶保护组织对放牧胁迫产生一定适应性变化,其抗旱性小幅度减弱,且角质层与表皮细胞的变化出现一致性。
导管作为植物水分疏导系统,导管直径的微小变化对植物水分运输效率都至关重要[33]。紫花针茅的最大导管直径和最小导管直径在重度放牧下较不放牧处理均减小,矮生嵩草叶基部的最大导管直径在中度放牧和重度放牧较不放牧处理减小。这可能是因为放牧导致叶长的减小,叶长在一定程度上表示水分在叶间的运输距离,而导管直径与运输路径长度之间存在正相关关系,表现为显著的异速生长关系[34]。异速生长指生物体某两个性状的相对生长速率不相同的现象,反映了生物体不同性状间的相关性[35]。本研究中不同放牧强度下紫花针茅的最大导管直径在不同取样部位之间均表现出差异,而最小导管直径也表现出基端粗、顶端较细的特点,进一步证实了叶片导管与运输距离之间的幂指数关系[34]。
3.3 矮生嵩草和紫花针茅对放牧的响应存在敏感性差异
不同物种的叶片形态结构对环境的响应敏感性存在极大差异[36]。将植物叶形态结构的响应程度作为衡量植物对放牧敏感性的标准,在植株个体普遍矮小、植物表型不易区分的草原地区更具科学性[37]。本研究中,重度放牧显著改变了紫花针茅和矮生嵩草的叶长,说明两种植物的叶长对放牧表现出较强的敏感性,这与Mclntir等[38]的研究结果一致。除此之外,放牧改变了矮生嵩草的近轴面表皮细胞厚,降低了最大导管直径,但增加了近轴面角质层厚,这表明除叶长外,放牧压力下近轴面角质层厚、近轴面表皮细胞厚以及最大导管直径也属于矮生嵩草的敏感性指标[39]。在放牧干扰下,矮生嵩草的叶长以及最大导管直径均减小,近轴面角质层厚和近轴面表皮细胞厚增加,体现了叶长与最大导管直径、近轴面角质层厚和近轴面表皮细胞厚之间的协同变化。非对称响应,即部分指标在同一放牧干扰下的相异变化,而本研究中叶长与最大导管直径、近轴面角质层厚和近轴面表皮细胞厚表现出的不同的变化趋势,便是矮生嵩草非对称性响应机制的体现,这点与冯斌等[40]的研究结果一致。除叶长外,紫花针茅的各项叶片结构均不受放牧影响,属于惰性性状[40]。这可能是由于在放牧干扰下,紫花针茅为保证自身叶片光合作用不受影响,对体内营养物质进行了分配优化,更倾向于维持地上部分的正常生理结构及功能[41],从而表现出叶结构上的相对稳定。在放牧干扰下,矮生嵩草的叶长、近轴面角质层厚、近轴面表皮细胞厚以及最大导管直径均表现为敏感性指标;而紫花针茅的各项叶解剖结构指标中仅叶长对放牧强度表现出较强的敏感性。对比认为,矮生嵩草和紫花针茅对放牧的敏感性不同,矮生嵩草叶片对放牧刺激的响应更为敏感。
4 结论
放牧压力下高寒草甸优势植物为适应环境变化产生多种适应性进化机制以保护其与非生物环境和放牧家畜的协调共存。植物应对放牧干扰会通过改变叶形态结构以及保护组织等重要叶结构以适应放牧压力,表现出较强的结构可塑性。其中,矮生嵩草对放牧刺激的响应较紫花针茅更加灵敏。矮生嵩草各项敏感性指标对放牧表现出非对称响应以适应放牧压力,而紫花针茅叶解剖结构相对稳定。
