油脂氧化对水产动物影响的研究进展
2023-11-01李永安郑普强李军亮程开敏李新宇
■ 李永安 宋 飞 郑普强 李军亮 程开敏 李新宇*
(1.广东粤海饲料集团股份有限公司,广东湛江 524000;2.华南师范大学生命科学学院,现代水产养殖科学与工程研究院,广东省水产健康安全养殖重点试验室,广东广州 510631)
油脂是鱼类重要能量来源,在体内完全氧化时每克能产生37.66 kJ 的能量,是糖类和蛋白质的2.25 倍[1]。此外,油脂能提供大部分水产动物不能自身合成的n-3和n-6系列多不饱和脂肪酸[2]。鱼油(富含EPA和DHA)和豆油(18:2n-6)因其富含多不饱和脂肪酸,对鱼类的机体健康和生长性能都有着积极的作用,是水产饲料主要的油源[3]。另一方面,相关油脂中富含PUFA 导致其在使用和储存过程中极易被氧化,产生各种毒害物质(醛、酮、醇和过氧化氢等)[4],从而降低油脂的营养价值,对动物的生长和健康造成一定影响。因此,人们通常选择在油脂或者饲料加工过程中加入抗氧化剂以防止油脂氧化。
普遍认为,油脂氧化会对水产动物造成负面影响,在饲料生产和储存中应避免发生。关于氧化油脂的研究,大部分的结果显示饲料中使用氧化油脂(鱼油为主)会抑制鱼类的生长性能,并对机体造成损伤。另一方面,部分研究结果显示使用氧化油脂对鱼体的生长无影响,甚至能促进生长。因此,有必要对油脂在饲料中的氧化过程、结果和影响进行系统总结。文章通过对饲料使用氧化油脂的研究进行分析,探讨其中结果差异的原因,并对各种鱼类对氧化油脂的耐受过氧化值进行统计,以期为饲料行业解决氧化油脂带来的危害提供理论依据。
1 油脂氧化及抗氧化剂
1.1 油脂氧化过程
油脂的氧化受多方面因素影响,其中活性物质发挥了关键作用。活性物质包括自由基和非自由基两类:自由基是指具有不成对电子的原子或基团,通常不稳定且有很强的反应性,如氧自由基[包括羟基自由基(•OH)、过氧自由基(RO2•)、烷氧基自由基(RO•)等]、氮自由基[一氧化氮(NO)和二氧化氮(•NO2)];非自由基类则是由氧或氮自由基转化而来的,如过氧化氢(H2O2)、次氯酸(HClO)、过氧亚硝酸盐(ONOO-)等。油脂氧化从脂肪分解成游离脂肪酸和低级脂肪酸(C≤10)开始,而后与活性物质发生氧化反应:当活性物质从不饱和脂肪酸酰基链中提取一个氢原子时,就会产生一个以碳为中心的脂质自由基(L•),而脂质自由基极易与氧再次反应形成脂质过氧基(LOO•),脂质过氧基则通过对附近的不饱和脂肪酸中提取一个氢原子,进一步发生氧化连锁反应,从而导致油脂的加速腐败[5]。过程中产生的脂质过氧化氢(LOOH)很容易分解成脂质烷氧基自由基(LO•),烷氧基自由基又可被转化为二次氧化产物(包括丙二醛、甲酮、乙醇等)。油脂自氧化过程的机制详见图1。除了油脂的自氧化外,油脂的氧化也受到原料残渣和微生物产生的酶类影响,这一部分称为酶解过程。这一过程主要是将脂肪酸分解成游离脂肪酸或与其反应生成醛、酮、醇等物质,一般与自氧化同时发生。油脂氧化还受到光照的影响,这一类称为光氧化。
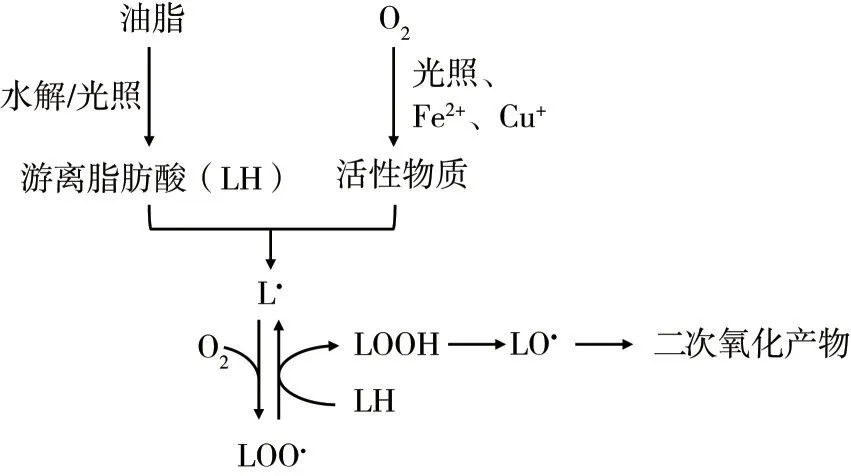
图1 油脂自氧化过程机制
水分含量是油脂氧化发生的一个关键因素。过高的水分极易使油脂发生水解,产生游离脂肪酸,而当游离脂肪酸达到一定比例时,就会加速其他脂肪的水解[6]。同样,紫外线也会导致游离脂肪酸的生成,同时也会促使氧气转化为活性物质[7]。此外,氧化过程也受金属离子(Cu+、Fe2+、Zn2+等)的催化影响。Sutton等[8]研究发现,铁和铜均可催化油脂过氧化,且铁对于油脂氧化的催化效果要优于铜,但两者在相同过氧化值下的抑制效果是否相同还有待研究。通过生产实践跟踪发现,常温下通常需要几个月才会发生油脂氧化,而在高温条件下几天就会酸败,说明温度对油脂氧化也有极其大的影响。同时,原料残渣和水分含量都会导致油脂中微生物的滋生,油脂本身也含有少量的油脂氧化酶,微生物生长繁殖过程中也会产生大量氧化油脂的酶类,加速油脂的变质[9]。因此,生产上对于油脂的储存要求是在阴凉避光处。
油脂的氧化程度一般由以下几个标准来进行评价(见表1):①过氧化值(POV),指油脂氧化过程中产生过氧化物的含量,但氧化后期过氧化物部分会被分解二级产物。②茴香胺值(p-AV),表示油脂氧化过程中产生二级产物(醛、酮、醌等)的总量,但这些物质在加热过程中则会被分解从而影响最后数值。③总氧化值(TOTOX;TOTOX=2POV+p-AV),基于过氧化值和茴香胺值计算所得,可相对客观体现油脂的氧化程度。④酸价(AV),是油脂中游离脂肪酸含量的标志,在一定程度上可以作为判断油脂质量的参考指标。油脂氧化是一个多步骤过程,由分解成游离脂肪酸开始,到氧化后产生氢过氧化物结束。因此,酸价在氧化过程中变化缓慢,如油脂氧化程度很高,但游离脂肪酸极少,酸价不高;另一方面,酸价高只能说明油脂中游离脂肪酸多,氧化程度不一定高。⑤碘值(IV),利用碘和双键的加成反应检测油脂中不饱和程度的高低,能在一定程度上反映油脂的氧化程度。⑥硫代巴比妥酸值(TBARS)或丙二醛(MDA)含量,丙二醛作为不饱和脂肪酸过氧化的降解二级产物中的主要醛类物质,在实际生产上多用来评估油脂的氧化程度。但是,不同种类的油脂氧化产生的丙二醛含量差异较大,例如植物油氧化后产生的丙二醛含量远小于动物油[10]。在科学研究中,研究者大多不用丙二醛作为油脂氧化指标。硫代巴比妥酸也能与饱和醛、单烯醛和甘油醛反应,因此并不能特异检测丙二醛的含量。⑦挥发性物质含量,主要检测油脂二级氧化后产生易挥发的醛类化合物(如丙醛、己醛等)。

表1 评价油脂氧化程度的指标
1.2 油脂氧化成分变化
油脂的氧化将导致其成分发生变化。研究表明,8种植物油(棕榈油、花生油、山茶油、菜籽油、葵花油、玉米油、紫苏油和大豆油)在30 d的加速氧化过程中,其酸价变化并不明显;除紫苏油和菜籽油外,其余6 种植物油的过氧化值在30 d 后检测均显著高于初始值;紫苏油、菜籽油和大豆油产生的丙二醛较其余植物油高;8种植物油在氧化过程中,生育酚都有不同程度的降低[11]。王永庆等[10]的研究中发现,植物油氧化后其主要脂肪酸(饱和脂肪酸、单不饱和脂肪酸、多不饱和脂肪酸)的相对含量并无变化。氧化条件对不同鱼油氧化的影响各异。光氧化对鳕鱼肝油化学特性的影响远大于其对凤尾鱼油,而热氧化条件对两种鱼肝油有相似的影响;对于鳕鱼肝油而言,相同时间内光氧化对其化学特征的影响远高于热氧化;对于鳀鱼油,热氧化对其化学特征的影响远高于光氧化。由此可见,即使在同等氧化条件下,不同油脂的氧化结果有所不同;该实验还发现,鱼油被高度氧化后,其过氧化值、茴香胺值、总氧化值、挥发物含量明显增加,并且会消耗自身存在的生育酚而加速变质,结果见表2[12]。与此同时,其长链不饱和脂肪酸会经过二次氧化产生各种挥发物(醇、醛、酮),但鱼油中主要脂肪酸种类(饱和脂肪酸、单不饱和脂肪酸、多不饱和脂肪酸)的相对含量在氧化过程中并没有变化。
1.3 机体抗氧化机制
油脂氧化会产生大量的氢过氧化物和二次氧化产物,会对鱼类的生长和健康产生不利影响。机体内含有大量非酶抗氧化物质或抗氧化酶以抵抗油脂氧化,大致分为以下几大类:①抗氧化酶类,包括谷胱甘肽(GSH)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)等,具有将体内形成过氧化物转换为毒害较低或无害的物质的功效;②维生素类,如维生素A(VA)、维生素E(VE)、维生素C(VC)和类胡萝卜素等,本身具有抗氧化性或具体调节相关抗氧化机制功能;③矿物质类,如铁、铜、锌、锰、硒等,以上元素作为辅酶因子或酶组成部分,也在机体抗氧化体系中扮演重要角色。
动物体内的抗氧化作用机理详见图2,主要可分为清除活性物质和阻断氧化连锁反应。油脂氧化过程是活性物质与不饱和脂肪酸双键发生加成反应的过程,因此活性物质清除是抗氧化的重要方法之一。例如,在SOD、GSH 和CAT等催化下,机体可以通过清除活性物质在初期阻断油脂氧化的发生。氧自由基则在SOD、CAT和GSH 的作用下还原成水和氧气。很多抗氧化物质(如虾青素)能向自由基提供电子或吸引自由基的未配对电子,有效地猝灭氧化性极强的单线态活性氧以及体内其他自由基。NO 是一自由基气体,在体内极不稳定,但该物质在体内可以与氧合血红蛋白快速反应,生成稳定产物NO3-(硝酸根)[13]。同时,NO 也与GSH 反应生成相对稳定的亚硝基硫醇[14]。油脂氧化会在LOO•形成后发生连锁反应,指数增加油脂氧化的速度,而VE 可以通过与LOO•反应生成LOOH中断该连锁反应,阻断油脂氧化。VC可以还原维生素E 自由基(VE•)为VE,间接协助阻断油脂的氧化。VE•和VC•都不是反应性物质[15],可被GSH 重新转化为VE 和VC。GSH 在机体抗氧化体系中扮演重要角色,参与多条反应路径。GSH 在与自由基反应后则会生成GS•(氧化自由基),但两个GS•可以反应生成氧化谷胱甘肽(GS-SG),并经过NADPH 依赖性谷胱甘肽还原酶(GSH-R)还原成GSH[16]。
2 氧化油脂对鱼类的影响
2.1 生长性能和饲料利用
研究表明,油脂氧化产生的腐败气味和有毒物质会影响养殖鱼类的食欲和消化性能,导致生长性能下降[17]。饲料油脂氧化对生长和饲料利用的危害性已经在多个物种中被证实,如拉氏鱥(Rhynchocypris lagowski)[18]、南亚野鲮(Labeo rohitaHamilton, 1822)[19]、北极红点鲑(Salvelinus alpinus)[20]、斑点叉尾鮰(Ictalurus punctatus)[21]等。随着氧化油脂添加量的升高,杂交石斑鱼(♀ Epi-nephelus fuscoguttatus×♂ Epinephelus lanceolatus)生长性能受到了明显抑制,饲料系数则表现为升高[22]。氧化油脂显著降低罗非鱼(Oreochromis niloticus)生长性能和消化酶活性[3]。也有研究表明,氧化油脂并不会影响虹鳟(Oncorhynchus mykiss)[23-24]、塞内加尔鳎鱼苗(Solea senegalensis)[25]、庸鲽(Hippoglossus hippoglossus)[26]的生长性能。长吻鮠(Leiocassis longirostris)[27]和 施 氏 鲟(Acipenser schrenckii)[28]生长性能虽然也不受饲料氧化油脂的影响,但其体色随着油脂的氧化程度而变暗。研究发现,氧化油脂短期对大西洋鳕鱼(Gadus morhuaLinnaeus,1758)生长无显著影响,但该实验整体的死亡率过高,可能会影响数据的准确性[29]。饲料添加氧化油脂对加州鲈(Micropterus salmoides)生长反而有促进作用[30-31]。由此可见,饲料添加氧化油脂对生长和饲料利用的影响在不同实验和鱼类中有一定差异。
通过比较相关试验的配方(见表3)也发现,不同实验料中的VC和VE含量差异较大,可能是影响实验结果的重要因素之一。部分试验饲料添加了大量的VC和VE,从而将油脂氧化产物消耗后仍有剩余,可能导致油脂氧化作用不明显。另一方面,不同物种对油脂氧化的敏感性也存在一定差异。氧化油脂会抑制大菱鲆(Scophthalmus maximus)和庸鲽的生长,而对金头鲷(Sparus aurata)的生长起到促进作用[32]。此外,大部分实验均以过氧化值为单一指标评价饲料氧化程度,该方法是否足够准确客观也需要进一步确定。关于油脂氧化对鱼类影响的相关实验的设计条件存在较大差异,油脂氧化对鱼类的影响需要一个标准来进行评估。
2.2 免疫和抗氧化能力
油脂氧化产生的自由基会攻击鱼体的免疫和抗氧化系统。因此,饲料油脂氧化对鱼体的免疫和抗氧化能力都会造成一定影响。研究发现,饲料中添加氧化油脂对虹鳟(初重46.9 g)的生长性能无影响,但增加了血清中丙二醛含量,对其抗氧化能力造成负面影响[23]。另一项研究发现,氧化油脂对虹鳟(初重62 g)的生长性能和抗氧化能力无显著影响[24]。以上两个研究结果差异可能受饲料过氧化值、VC 和VE 水平差异的影响,同时也有可能受实验鱼规格不同而影响,生长后期的鱼对氧化应激的承受能力可能更高。饲料添加氧化油脂反而引起加州鲈抗氧化酶活性的升高,可能是因为脂质过氧化自由基激发了加州鲈的抗氧化防御体系,引起了抗氧化酶活性的上升[30]。高氧化油脂能激活塞内加尔鳎鱼苗的抗氧化防御体系[25]。以上结果说明,饲料中油脂氧化后产生的自由基会攻击鱼体的抗氧化防御体系,激发酶活性的升高,但机体防御体系不足以解决自由基时,就会造成机体油脂氧化产物丙二醛的积累,导致机体受到氧化损伤,降低机体抗氧化能力。在饲料中添加氧化油脂不会对鱼类的死亡率造成影响,仅发现大西洋鳕鱼的研究中整体死亡率过半,但对照组与试验组并未有差异[29]。饲料中的氧化油脂会降低鱼类免疫力[33]。饲料氧化鱼油水平增加减少了杂交石斑鱼肠道菌群的多样性,并改变了肠道菌群的结构,从而进一步影响健康[22]。
2.3 组织健康
虽然饲料添加氧化油脂对鱼类的生长影响在不同实验中有一定差别,但对于组织的氧化压力却是随着氧化程度增加而普遍明显增加。对南亚野鲮[19]的研究表明,油脂氧化程度的增加降低了肝脏和肌肉中VE 的含量,并增加了肝脏的氧化压力,从而影响组织健康。例如,对石斑鱼研究表明,氧化鱼油会导致肝脏中的营养成分、氧化应激、炎症、形态异常和转录组的改变[34]。类似的结果在北极红点鲑[20]、罗非鱼[28]、虹鳟[24]中被发现。虽然氧化油脂对庸鲽[26]的生长性能无影响,但高水平的过氧化值则会影响肝脏和肌肉的丙二醛含量。研究也发现,高氧化程度的油脂会使鱼体肝组织出现脂肪空泡和核偏移[30],并引起肝脏中炎症细胞增多[28]。此外,油脂氧化同时会显著影响鱼体肠道的杯状细胞数量、绒毛长度和肌肉厚度[21],对消化吸收能力产生影响。最新研究表明,氧化鱼油导致杂交石斑鱼脾脏氧化应激,导致脾脏组织损伤[35]。由此可见,油脂氧化会损害鱼类多个组织器官。
2.4 信号通路和代谢
研究表明,氧化油脂可通过调节相关信号通路而影响机体。实验发现氧化油脂通过影响Keap1-Nrf2-ARE信号通路,对拉氏鱥生长性能和抗氧化能力起到显著的负面影响[18]。短期添加氧化油脂会影响罗氏沼虾(Macrobrachium rosenbergii)IMD 和Toll 信 号通路,导致其抗氧化能力降低,体内丙二醛含量升高,从而降低了罗氏沼虾的生长性能[36]。氧化油脂对鱼体的脂质代谢也会产生负面影响[21,37]。由此可见,氧化油脂对鱼类的影响是多方面的:一方面是氧化油脂产生的过氧化自由基通过影响抗氧化系统信号通路(Keap1-Nrf2-ARE 信号通路),影响鱼类抗氧化能力,对组织造成氧化压力,从而影响生长;另一方面则是通过鱼体内丙二醛的蓄积,激活免疫系统信号通路(IMD/Toll 信号通路),对生长性能产生负面影响。此外,油脂氧化后变质将影响鱼体对脂肪代谢的相关基因表达,导致鱼体血脂和胆固醇异常,从而影响鱼体健康。
3 抗氧化功能性物质
3.1 氨基酸
氨基酸作为抗氧化酶合成的物质,对油脂氧化也有一定的抑制作用。一些功能性氨基酸(精氨酸、组氨酸、脯氨酸、牛磺酸等)和小肽(如谷胱甘肽)已被证实具有抗氧化功能。牛磺酸作为一种非蛋白氨基酸,具有广泛的生理功能,包括渗透调节、抗氧化和抗炎功能[38-39]。牛磺酸可通过促进脂质代谢的转录因子而减少肝脏中的脂质沉积,同时影响Nrf2-Keap1信号通路缓解氧化油脂诱导的肝脏氧化损伤[21]。此外,由谷氨酸、半胱氨酸以及甘氨酸组成的谷胱甘肽,不仅能协助机体维持正常的免疫功能,本身也具有强抗氧化能力。作为一种特殊氨基酸,L-肉碱通过改变拉氏鱥脂肪合成代谢相关基因表达量来影响肝脏和肌肉中脂肪酸含量,从而缓解氧化油脂带来的影响[40]。氨基酸作为抗氧化功能物质,已有较系统的总结[41]。
3.2 维生素
许多维生素均有抑制自由基产生的作用,对氧化油脂带来的危害也有缓解作用。VC 和VE 的添加均能改善真鲷饲喂氧化油脂带来的危害[42-43]。同样的结果在南亚野鲮[19]、虹鳟[23]、庸鲽[26]和大西洋鳕鱼[29]中均有发现。饲料中的VE 能通过与脂质自由基反应,清除自由基,抑制油脂继续氧化。另一方面,饲料中缺乏VE 则会降低肝脏中过氧化氢酶、谷胱甘肽过氧化氢酶和谷胱甘肽还原酶的活性[44]。除了VE 和VC外,VA、VK2和烟酸也具有抑制自由基产生的作用。其中,类胡萝卜素是一种较为有效的抗氧化剂,包括α-胡萝卜素、β-胡萝卜素和γ-胡萝卜素、虾青素等[45]。其中,叶黄素、玉米黄质、β胡萝卜素的抗氧化活力是VE的10倍,而虾青素抗氧化活性是VE的100倍[46]。β-胡萝卜素能够与过氧自由基反应生成稳定的自由基,从而抑制过氧自由基的链传播反应。同时,作为NADP+/NADPH,NAD+/NADH 和FAD/FADH2组成部分的烟酰胺和核黄素在抗氧化应激系统中也发挥着至关重要的作用[45,47]。
3.3 矿物质
矿物质的抗氧化能力主要是通过影响酶的活性来进行,这在营养学和生物化学中被广泛证实。镁作为葡萄糖-6-磷酸脱氢酶和6-磷酸葡萄糖酸脱氢酶的辅助因子,参与了戊糖循环催化NADPH+生成NADPH。因此,提高镁离子的浓度能够降低人体外单核细胞中GS-SG 和MDA 的含量[48],饲料中补充镁能够提升草鱼体内谷胱甘肽的含量,提高草鱼的抗氧化能力[49]。抗氧化酶SOD 按含有不同金属辅基而分为三大类,分别为Cu/Zn-SOD、Mn-SOD、Fe-SOD。铜、锌和锰分别是Cu-SOD、Zn-SOD 和Mn-SOD 酶中不可或缺的金属离子。金属离子作为酶类中的辅助因子,其在饲料中含量高低会影响相应酶活性。因此,饲料中缺乏这些矿物质会显著降低Cu-SOD、Zn-SOD 和Mn-SOD 的活性,并引发过氧化物损失和线粒体紊乱[45]。饲料中补充适量铜和锌能提高鱼体的抗氧化能力[50]。硒作为相关酶的必要辅助因子,在机体抗氧化能力中发挥着重要作用。该物质不仅能显著提升机体谷胱甘肽过氧化氢酶的活性[50],也能减少过量的脂质在肌肉中沉积[51]。如在中国首次发现的克山病病因就是土壤中缺乏硒,显著降低谷胱甘肽过氧化物酶90%的活性[52],从而影响了机体正常健康。
4 饲料油脂氧化控制措施
4.1 合成抗氧化剂
在生产上,人们为了抑制油脂氧化往往会在油脂或饲料中添加合成或提取的天然抗氧化剂来抑制油脂的氧化。众多抗氧化物质根据作用机理可分为三类:清除活性物质、阻断连锁反应和间接抗氧化剂。常用化学合成抗氧化剂包括乙氧基喹啉(EQ)、丁基羟基茴香醚(BHA)、没食子酸丙酯(PG)和特丁基对苯二酚(TBHQ)等。这些抗氧化剂主要通过提供氢离子与过氧自由基反应来阻断氧化的链增长反应,从而终止自由基链式反应。研究表明,EQ 的添加量小于150 mg/kg 较为安全,但添加量为300 mg/kg 时抗氧化保护能力最强[53]。在水产饲料的生产中,人们常添加EQ 或二丁基羟基甲苯(BHT)用以防止饲料氧化,但其剂量需要严格把控。EQ 本身具有一定副作用。此外,该物质是对氨基苯乙醚和丙酮在催化剂的存在下加热脱水缩合制得,对氨基苯乙醚是一种剧毒物质。因此,对氨基苯乙醚残留量过高的EQ是禁止在动物饲料中添加使用。后续研究发现,EQ在机体内会被代谢成脱甲基-EQ、醌亚胺-EQ和EQ二聚体(EQDM),其中EQDM的半衰期远高于EQ本身,因此在机体内的含量会远高于EQ的添加量[54-55]。研究也发现,鱼类禁食期间并不能将体内残留的EQ及其代谢物完全排出体内,存在较高安全风险[55]。同样的,BHT以对甲酚、异丁醇为原料,以浓硫酸作为催化剂,氧化铝作为脱水剂而合成,其过多摄入对于动物而言具有毒性作用。因此,合成抗氧化剂的添加量需被谨慎评估。
4.2 天然抗氧化剂
相比于合成的抗氧化剂,植物中存在的抗氧化剂安全性更高、抗氧化能力强、副作用更少。研究表明,植物中提取的化合物(包括酚类、多酚类的化合物),如类黄酮和儿茶素,都具有很强的抗氧化活性,能够有效清除自由基[15,56]。添加茶多酚和茶多酚棕榈酸酯组成的复合抗氧化剂能够很好的保护南极磷虾油不被氧化[57],同时茶多酚有增强红细胞的抗氧化应激能力[58],并且能够有效清除超氧化物和羟基自由基[59]。同样的,在饲料中补充茶多酚能够提高鱼体的抗氧化能力,并且降低血清中丙二醛的浓度[60]。另一方面,植物中的酚类化合物虽然有抗氧化的功能,但也会与氨基酸(如蛋氨酸)相结合,影响鱼类对蛋白质的吸收利用,降低生长性能。因此,在饲料中补充酚类化合物时需要注意其与其他营养物质的相互影响。
5 结论及展望
综上所述,氧化油脂会产生大量的脂质过氧化自由基和醛、酮、醇等有毒有害物质,对鱼体的抗氧化防御系统、免疫系统和营养代谢系统造成一定的影响。油脂氧化也对鱼类机体肝脏、肌肉、肠道等组织器官造成氧化压力,引起损伤或影响品质。此外,油脂氧化也影响肠道上皮细胞杯状细胞数目、绒毛长度和肌层厚度,从而影响鱼体对营养素的消化吸收,导致鱼体生长受阻。大量研究表明,饲料中补充VC、VE 或其他抗氧化剂均能改善油脂氧化带来的危害。合成的抗氧化剂由于会在鱼体积累聚集且部分化合物对人体有害,需要谨慎评估其添加量。未来需要进一步大力探索和开发可代谢、安全性高、抗氧化效率高的天然抗氧化剂。虽然油脂氧化在鱼类中进行了大量研究,但仍然缺乏一套标准研究方法和条件,导致相关数据相互参考和对比性不强;油脂的众多氧化评价指标会随着油脂种类和氧化条件和程度不同而发生改变,需要建立标准化的研究方法及评价条件用以氧化油脂的相关研究。此外,有关油脂氧化对鱼类所造成相关影响的机理仍然了解较少,需要进一步探索。
