植硅体视角下浙江宁波鱼山遗址的生业考察
2023-10-25夏秀敏王力之雷少王结华郑晓蕖赵志军吴妍
夏秀敏 王力之 雷少 王结华 郑晓蕖 赵志军 吴妍

摘要:鱼山遗址是宁绍平原河姆渡文化时期以来的重要遗存,其考古发现为探讨东部沿海地带长时间尺度的水稻驯化进程和人地关系提供了良好素材。高精度的植硅体分析表明,河姆渡时期以来水稻扇型个体呈现了变小稳定的趋势,并逐渐向中间型和长柄型演化,暗示先民是以种植粳型化的水稻为主;水稻扇型植硅体鱼鳞状纹饰的变化揭示了在河姆渡文化晚期,水稻的驯化基本完成。受海平面变化的频繁影响,该地的水稻驯化具有一定的滞后性,驯化的周期也有所延长。结合植硅体分析和多种研究成果,推断在河姆渡文化时期,鱼山遗址的稻作生产获得了较大发展;在河姆渡文化晚期,水稻逐渐成为先民植食性资源的主体;在滨海多变的环境背景下,鱼山遗址形成了以水稻种植为主,采集和渔猎并行的多元化生业模式。良渚晚期,伴随着水稻驯化的完成和生产工具的进步,推测出现了系统的水田耕作系统。商周时期,依托暖干的环境,稻作生产向集约化发展,为古越国的兴起奠定了基础。
关键词:鱼山遗址 宁绍平原 植硅体 水稻 生业模式
Abstract: The Yushan Site is an important remain from the Hemudu culture in Ningshao Plain. Its archaeological excavation provides a good material for exploring the process of rice domestication over a long time scale and man-land relationship in the eastern coastal zone. The high-resolution phytolith analysis shows that the bulliform phytoliths have become smaller and more stable, and gradually evolved into intermediate and long-stalk types, which indicated that japonica rice was mainly cultivated by the ancestors since the Hemudu period. The change of fish-scale decorations of rice bulliform phytoliths revealed that full domestication of rice was not completed untill the late Hemudu period in this region. Due to the frequent influence of sea level changes, rice domestication had a certain lag, and the domestication period was also prolonged in this region. Combined with the phytolith analysis and recent research results, it can be concluded that rice farming of the Yushan Site had been developed greatly during the Hemudu period. Meanwhile, Rice gradually became the main plant resources of the ancestors in the late period of Hemudu culture. Under the background of the variable coastal environment, it has been formed a diversified subsistence strategy containing rice cultivation, gathering and fishing and hunting. During the Liangzhu period, with the completion of rice domestication and the progress of production tools, it can be speculated that systematic paddy field farming occured, but the diversification of resource utilization still continued. During the Shang and Zhou dynasties, relying on the warm and dry environment, rice agriculture turned to intensive development, which laid the foundation for the rising of the ancient Yue State.
Keywords: Yushan Site Ningshao Plain Phytolith Rice Subsistence economy
一、 前 言
寧绍平原是浙江省东北部、杭州湾南岸的一片东西向的海岸平原,因其优越的地理位置和气候条件,成为我国新石器文化的重要分布区和稻作农业的起源地。其中,该地区以20世纪70年代余姚河姆渡遗址的发掘和大量稻属遗存的发现最为引人瞩目,对于探讨中国水稻栽培的起源及其在世界稻作农业史上的地位具有重大的意义。近二十年来,伴随着上山、小黄山、荷花山、湖西等新石器时代早、中期遗址稻属遗存的发现,进一步确立了水稻在距今1万年左右于长江下游地区起源的观点和认识。受区域环境和文化传播等方面的影响,水稻在长江下游各地区传入的时间、路径等存在着一定的差异,水稻驯化的速率也不尽相同,稻作生产的发展水平亦不均衡。
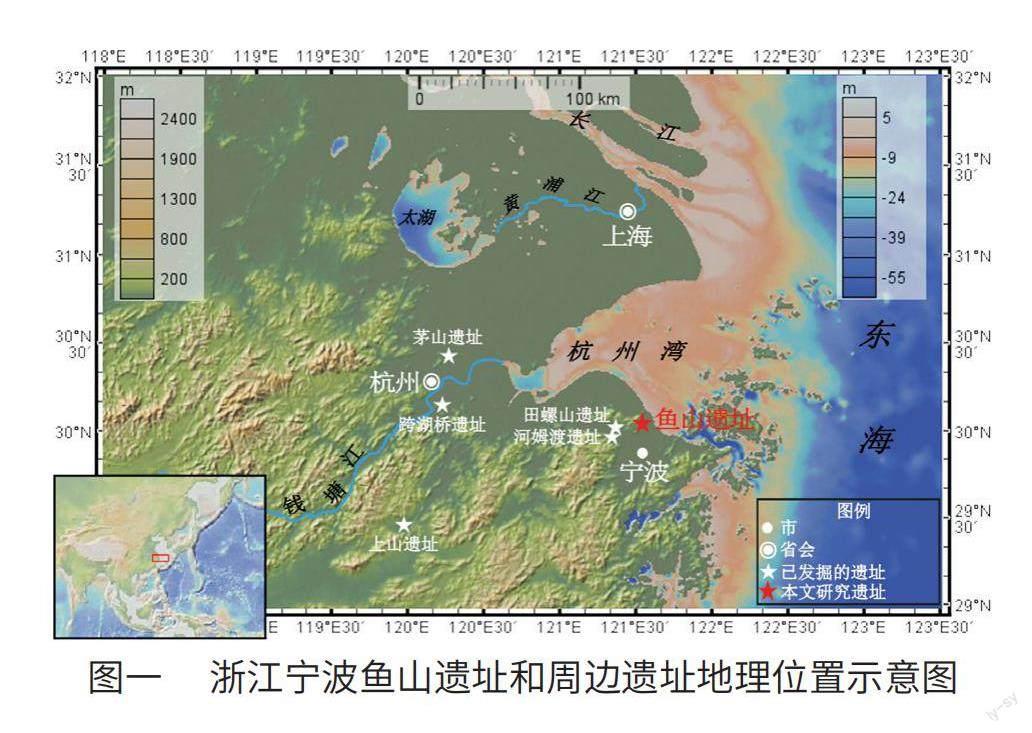
鱼山遗址位于宁绍平原东部,是目前发现的距离现代海岸线最近的河姆渡文化早期遗址。全新世以来,沿海地区曾发生多次海平面波动,并在不同程度上影响了周边地区的文化发展轨迹和生业模式。有研究表明,河姆渡文化时期,稻作农业仍处于较为初期的发展阶段,水稻正处于驯化的过程中,河姆渡仍是一个“以坚果采集和渔猎为主要生计模式的早期社会形态”。以河姆渡、马家浜、崧泽文化等为代表的典型遗址的动植物考古分析表明,该阶段长江下游地区的经济模式包括以水稻种植与湿地、园圃管理并行的植物资源和以狩猎、捕鱼、采集螺蚌为主、家养动物为辅的动物资源。植硅体分析揭示了长江下游多个遗址的水稻驯化进程大约持续了5000年左右,并在良渚文化时期驯化完成。同比之下,处在宁绍平原东部沿海地带鱼山先民的生业模式和水稻驯化进程是否遵循长江下游地区的演变趋势亦或有不同之处等问题尚未得到深入分析。
鱼山遗址大植物遗存发现的早期炭化水稻和杂草资源为探讨该地的水稻驯化和稻作生产提供了直接参考证据。基于植硅体原地沉积、产量大、耐腐蚀、易保存的优良特性及其在水稻属性鉴定、水稻驯化动态过程、稻田亩产恢复等方面的定性和定量的研究,本文依托高精度的植硅体分析,同时借助多方面的研究成果,尝试重建该地长时间尺度下水稻的驯化和生业模式变化。
二 、材料与方法
(一)遗址背景和剖面特征
鱼山遗址位于浙江省宁波市镇海区九龙湖镇河头村,鱼山山脚的东南麓,东距海岸线直线距离约7.3公里,地理坐标为30°02′N,121°33′E(图一)。2013年至2015年间,宁波市文物考古研究所(现宁波市文化遗产管理研究院)先期对遗址进行大规模勘探,确认遗址分布面积约16500平方米。后联合吉林大学、南京大学等多家单位进行两次发掘,发掘面积共计4300平方米。遗址包含了河姆渡时期、良渚时期、商周至唐宋等四个不同时期的文化堆积,清理出300余处遗迹单位和各时期文化标本600余件,发现有大量的灰坑、灰沟、水井等遗迹和丰富的陶、瓷、铜、石、骨和木器等遗物。
发掘情况表明,该遗址地势自北侧山坡向南部平地逐渐降低,地层堆积深度1.0~2.5米,可分为9个大层。现以T0514西壁为例,结合遗址剖面整体变化,对鱼山遗址的地层堆积情况介绍如下。
①层为耕土层,浅灰褐色,土质较疏松,厚0~18厘米。
②层为黄灰色土,土质致密,堆积浅,遗迹少,距地表0~20厘米,厚8~20厘米,分布于整个探方。出土遗物以瓷器残片为多,也有少量陶器,可辨器形有青瓷碗、钵、盘、盒、盂等。该层属于唐宋时期的堆积。
③层为灰褐色砂土层,土质较疏松,距离地表10~25厘米,厚10~30厘米,分布于各探方。出土遗物丰富,包括印纹硬陶、原始瓷、陶器残片等在内。可辨器形有罐、碗、盏、鼎等。该层属于商周时期的堆积。
④層为黄灰色土层,土质致密坚硬,包含大量粉砂和褐色锈状斑,距离地表10~30厘米,厚度为60~80厘米。无包含物。
⑤层分为上下两个亚层,上部⑤a层为青灰色淤泥,土质纯净松软,距离地表60~110厘米,厚约10~60厘米,分布于各探方,为海相沉积层;下部⑤b层为黄砂土层,土质较硬且夹细砂,距离地表70~130厘米,厚约5~30厘米,仅分布于斜坡和高台上,包含物主要为陶片。⑤b层应为良渚文化晚期——钱山漾文化时期堆积。
⑥层可分为上下两个亚层,土质松软,距地表110~165厘米,厚约5~30厘米。呈水平堆积分布于南侧各探方中。其中⑥a层为泥炭层,出土少量遗物,可见鱼鳍形和“T”形鼎足等。⑥b层为浅黑色土,出土少量绳纹陶釜和鼎足残片,属于河姆渡文化晚期后段堆积。
⑦层为灰褐色土层,土质松软,距地表140~160厘米,地层厚10~25厘米。出土遗物以陶器为主,可辨器形有豆、罐、釜、支座等。属于河姆渡文化中期堆积。
⑧层为青灰色淤泥层,土质较黏且纯净,距地表160~190厘米,地层厚度10~60厘米,分布于各探方。未见人工遗物,属于海相沉积层。
第⑨层可分为上、下两个亚层,⑨a层为灰黑色土,土质较疏松,距离地表180~210厘米,地层厚度为5~10厘米,出土器物以陶器残片为主。⑨b层为浅灰色土,土质松软,距地表185~215厘米,地层厚度10~35厘米,遗物以陶器为主,器形有夹炭陶釜、豆、盘、罐等,动物骨骼较少,植物遗存较丰富。该层属于河姆渡文化早期堆积。发掘者认为⑨a层为泥炭层,⑨b层存在水稻田堆积。
⑨层下为细腻、纯净的青灰色生土。
(二)样品采集与实验
鉴于宁波市文物考古研究所将遗址几个剖面整体切割打包回库房,我们的土壤样品采集工作是在库房内进行的。选取了堆积较为深厚的T0514的西壁剖面进行柱状采样,自下而上按照5厘米或10厘米的间隔逐层位取样。取样时先用手铲对剖面表层进行清理,避免外来土壤和各层位之间的污染。每个样品采集约100克土样进行植硅体分析,共获取了52份土样。所有的样品均在中国科学院古脊椎动物与古人类研究所脊椎动物演化与人类起源重点实验室进行分析和处理。
考虑到南方土壤的黏性较大,水分过多,不易分散,事先将土样进行烘干、研磨。同时为了降低炭屑或有机质过多等对植硅体提取和观察造成影响,将研磨后的样品放入电炉中450℃烧制8小时。采用常规的重液浮选法对植硅体、硅藻、海绵骨针化石进行同步提取,过程如下:将5克干燥土样放入烧杯,加入30毫升双氧水(H2O2)放置加热板上反应20分钟左右去除有机质;待反应结束样品冷却后继续加入稀盐酸(10%HCl)煮沸,用以去除铁、钙等矿物质;样品冷却静置24小时后加入纯水离心三次,以洗净盐酸为止;研磨石松孢子(27560个/片)溶解于土样中,再加入比重为2.3~2.4的溴化锌重液,吸取上清液后离心清洗三次,将提取物放置烘箱中晾干;最后用加拿大树胶进行制片,置于Nikon公司生产的eclipse LV100POL显微镜(200× 和500×)下进行观察、鉴定、统计和拍照。
植硅体形态的分类参考了现代植物植硅体分析结果和相关文献。每个样品随机统计700粒左右的植硅体,以期提高鉴定的分辨率。植硅体百分含量的计算是以所有植硅体类型总量为标准,每个类型数量与之相比得到的。同时在显微镜下尽量挑选平面形态对称、发育完整的水稻扇型植硅体进行测量,统计指标包括扇型植硅体的长度(VL)、宽度(HL)、扇柄长(a)、扇面长(b)以及鱼鳞状纹饰数量,每一份样品的统计基数为50粒。最后,将数据录入Excel表格中,利用SPSS等软件进行分析。此外,观察中发现的水稻双峰型植硅体也进行拍照和参数测量。在保证植硅体数量统计完成的同时,对观察中发现的硅藻和海绵骨针另计数统计。
三 、結 果
(一)测年结果
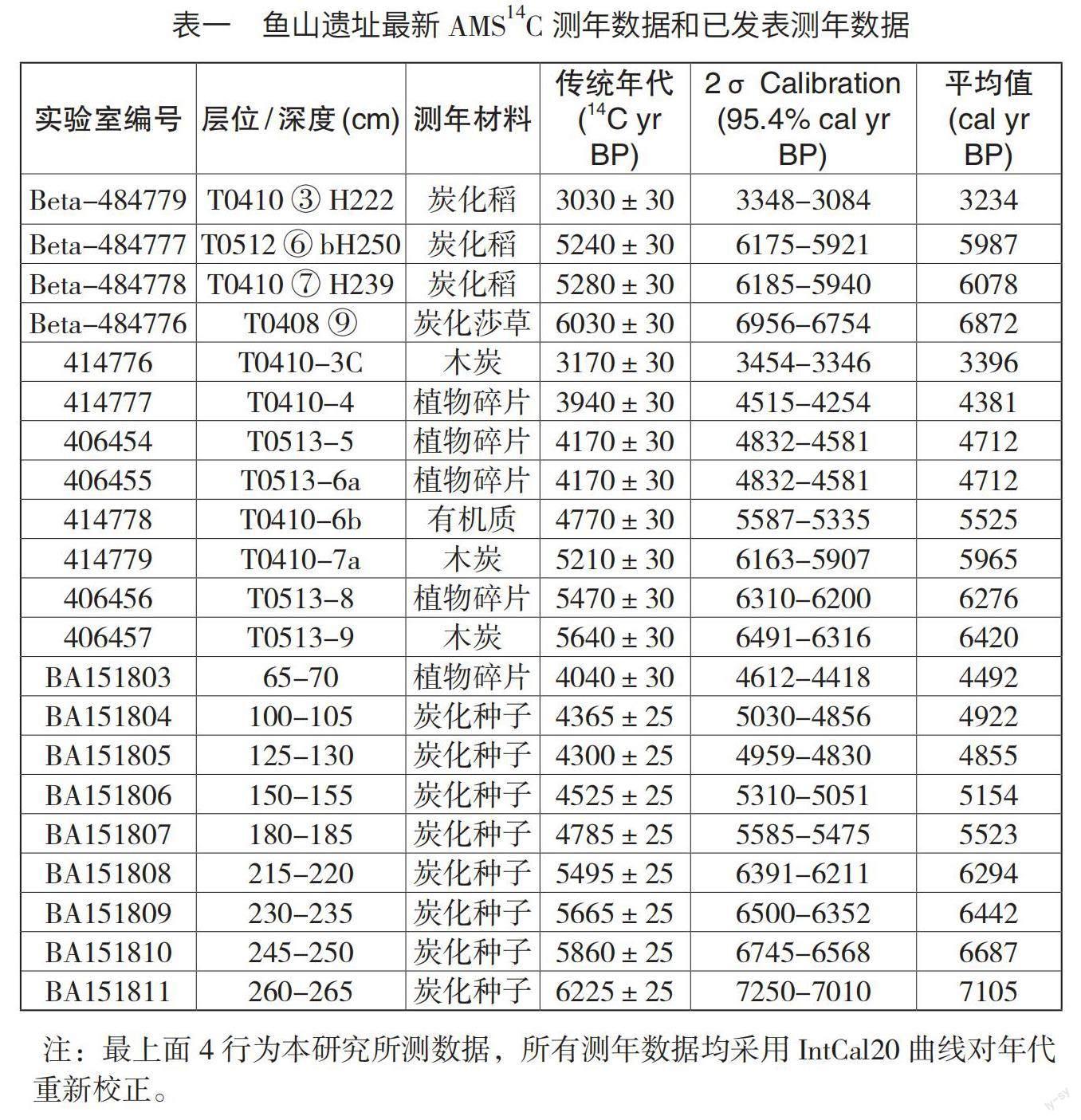
为了探索遗址各时期的发展脉络和不同阶段水稻驯化的进程,选用了该遗址初期浮选的一年生的炭化种子进行了测年,并采用IntCal 20曲线进行年代校正。结合遗址已经发表的测年数据与考古地层相对年代的一致性,同时参考宁波市文物考古研究所最新的分期断代结果,大致确定遗址各文化时期的年代框架(表一)。鱼山遗址文化层⑨层的绝对年代为6875-6295cal.a B.P.,相当于河姆渡文化早期前、后段,对应传统的河姆渡文化一、二期;文化层⑦层的绝对年代为6085-5815cal.a B.P.,相当于河姆渡文化中期,对应传统的河姆渡文化三期;鱼山遗址⑥b层绝对年代为5525-5155cal.a B.P.,相当于河姆渡文化晚期后段,对应河姆渡文化四期;鱼山遗址⑤b层绝对年代为4685cal.a B.P.,对应良渚文化晚期;③层的测年数据不多,集中在3400-3200cal.a B.P.,相当于商代中晚期,但是该层中也包含两周时期的遗物。因该遗址自然层和文化层相互叠压堆积,测年数据难免存在一些缺环。
(二)植硅体鉴定结果
在该剖面的样品中共鉴定出20余类植硅体形态,统计20000余粒植硅体。常见的植硅体形态有哑铃型、尖型、方型、长鞍型、短鞍型、竹扇型、芦苇扇型、齿型、平滑棒型、刺棒型、莎草型和阔叶木本植硅体等(图二)。还发现了相当数量的水稻特征型植硅体,包括水稻双峰型、水稻扇型及水稻横排哑铃型(图三)。此外,还发现了少量(疑似)稗子稃壳的植硅体形态。根据植硅体的气候指示原理和现代表土植硅体的分布规律,将发现的草本植物植硅体形态大致分为两类,示暖型植硅体(扇型、方型、长方型、哑铃型、短鞍型)和示冷型植硅体(棒型、尖型、帽型、齿型)。除植硅体外,同步观察到一定量的硅藻和海绵骨针。
根据不同层位植硅体的类型分布和百分比组合,T0514西壁剖面自下而上可分为5个组合带(图四)。
带1(河姆渡文化早期,对应⑨层):植硅体平均总浓度为147968个/克干样,植硅体类型丰富。代表暖湿环境的阔叶木本植硅体、鞍型、扇型、方型等占有相当数量(平均约51%),指示周边水环境的硅藻和海绵骨针含量(平均约3%)较高,呈现了温暖湿润的气候特征。其中水稻植硅体平均占约3.7%,以水稻扇型为主,未见水稻双峰型,表明稻作农业在该地的初步发展。
带2(自然堆积,对应⑧层):植硅体平均浓度骤降到65501个/克干样。示暖型植硅体数量显著减少(平均约14%),示冷型植硅体的含量有所增加(平均约58%),水稻植硅体含量锐减甚至极少发现;代表水环境的海绵骨针数量猛增(平均约29%),呈现了湿冷的气候特征,同时推测这一时期受到海侵的影响,稻作种植衰退。
带3(河姆渡文化中期-良渚文化晚期,对应⑦层、⑥层、⑤b层):植硅体总浓度持续升高,平均为272780个/克干样。示暖型的植硅体含量显著上升(平均约35%),示冷型的植硅体较前期有所减少,代表水体环境的硅藻和海绵骨针(平均约5%)大幅降低,在一定程度上表明水域面积减少,海平面较为平稳,表明了相对温暖湿润的气候特征。水稻植硅体的含量逐渐增多,在良渚晚期达到了峰值(9%),揭示了稻作生产的强化。
带4(自然堆积,对应⑤a层和④层):植硅体总浓度急剧下降,平均为75940个/克干样。良渚文化晚期堆积下又出现了自然淤积层,指示水环境的海绵骨针含量大幅增加(平均约26%),示冷型的植硅体含量略有升高(平均约52%),水稻植硅体的含量极低。整体呈现出略湿冷的气候特征,同时伴随着水稻生产的急剧衰退。
带5(商周-唐宋时期,对应③层、②层):植硅体总浓度增加至整个剖面带最大,平均约为819076个/克干样。示暖型的植硅体含量大幅升高(平均约51%),代表水体环境的海绵骨针(平均约5%)与硅藻浓度急剧减少,水稻植硅体含量较为平稳(平均约7.5%)。凸显了一种暖干的环境特征,稻作农业稳定发展。
(三)水稻植硅体形态测量结果
1. 水稻扇型植硅体测量结果
水稻扇型植硅体的长(VL)、宽(HL)和b/a值是扇型植硅体的一个重要形态测量指标(图三,4)。按照实际观察效果,对每个样品尽可能统计50粒以上的水稻扇型植硅体,共计756个。受海水侵蚀和埋藏环境等影响,部分层位观察效果不甚理想,统计数量低于50粒的标准。将实际观察和测量后的数值按照时代进行分类统计。从图五,a和表二可见,鱼山遗址水稻扇型植硅体河姆渡时期长、宽均值为46.62μm和43.71μm,至良渚晚期分别为45.72μm和38.5μm,商周时期为44.01μm和38.79μm,到了唐宋时期为39.46μm和35.24μm。水稻扇型植硅体长、宽值呈现出从早到晚不断减小的趋势,凸显了扇型个体渐趋变小的特点。
从图五,b可以看出,河姆渡时期水稻扇型b/a平均值为0.83,良渚时期增长到0.97,到商周和唐宋时期分别为1.01和1.02。河姆渡时期b/a平均值低于1,分布略松散;到良渚时期,b/a均值上升至0.97;同比之下,商周和唐宋时期均值都接近于1,数值分布较为集中。水稻扇型植硅体b/a值从河姆渡时期至唐宋时期,大致经历了一个由低到高的缓慢增长阶段,并最终趋于稳定。
2. 水稻扇型植硅体鱼鳞状纹饰数量变化
依据吕厚远等学者的研究,将水稻扇型鱼鳞状纹饰≥9个的归为驯化型,<9个的归为野生型(图六)。各时期观察到的扇型鱼鳞状纹饰清晰的个数总计614个,依据鱼鳞状纹饰个数不同分3组进行统计,并结合层位变化和时代归属将其整合(图六)。可以看出河姆渡文化早期水稻扇型鱼鳞状纹饰≥9个的比例为39%,河姆渡文化中期驯化型的比例为47%,河姆渡文化晚期驯化型的比例达到59%;到良渚文化晚期,这一比例攀升至71%,达到了最大值。商周和唐宋时期,驯化型比例分别达到64%和65%,趋于稳定。
3. 水稻双峰型植硅体测量结果
双峰型植硅体,通常产生于水稻颖壳或稃壳之中,是识别水稻和鉴定水稻驯化性状的重要指标(图三,3)。在鱼山遗址的剖面植硅體鉴定中,共观察到30个水稻双峰型植硅体,其中唐宋时期文化层中有26个,商周时期3个,良渚晚期1个。赵志军等学者通过测量水稻植硅体双峰间距(TW)、体宽(MW)、垭深(CD)、双峰高(H1 和 H2)5 个变量提出了用判别函数区分野生稻和驯化稻的方法,鉴定准确性高达90%以上。该标准和判别方法已多次应用于史前遗址中。考虑到遗址发现的水稻双峰型植硅体以历史时期为主,水稻驯化属性明确,鉴定意义不大。仅对来自良渚时期和商周时期的4个水稻双峰型植硅体测量鉴定,结果表明都是置信度较高的驯化稻。
四、分析与讨论
(一)鱼山遗址各时期水稻遗存的性质和驯化趋势
水稻扇型植硅体形态测量和考察多次被用来区分现生野生稻和驯化稻,一定程度上可以反映水稻遗存的性质和驯化过程。一般而言,驯化植物的植硅体相比野生种类来说,尺寸更大。Fuller等对长江下游地区马家浜文化至良渚文化时期水稻扇型植硅体研究显示,其长、宽值逐渐增加。但是具体到鱼山遗址,其水稻扇型植硅体的长、宽均值从河姆渡早期到唐宋时期出现了不断减小的趋势,水稻扇型个体渐趋变小稳定。该结果与驯化稻具有更大的植硅体形态这一传统认识相矛盾,也不同于前人对上山遗址、跨湖桥遗址等地水稻扇型植硅体大小变化趋势的分析。推测可能是水稻驯化的节点从河姆渡时期以来发生了重大转变,前期不断变大的趋势在该时期之后趋于平缓和集中,预示着水稻驯化渐趋完成和结束。当然也可能是河姆渡时期水稻扇型样本量略少,一定程度上影响了统计分析结果。
不同于水稻扇型个体的长、宽均值变化,其个体b(扇面长)/a(扇柄长)值呈现出了从河姆渡时期到唐宋时期不断增加且渐趋稳定的趋势,均值基本上稳定在1左右。前人对扇型植硅体进行了分类,b/a<0.80为长柄型,b/a>1.20为短柄型,b/a=0.8~1.2为中间型。由图五,b和表三来看,鱼山遗址水稻扇型植硅体中间型占据了主体,其次是长柄型。唐先干等对现代水稻叶片植硅体大样本数据的统计分析表明,在遗址水稻鉴定中,如发掘的扇型植硅体中多数b/a≤1.20就有很大可能是粳稻。由此推断,鱼山遗址从河姆渡时期开始,应该是以种植粳型化的水稻为主。当然,有学者也提出,水稻扇型植硅体的大小并不是由单一遗传因素控制的,叶片成熟度、水热条件等均会对扇型植硅体的大小产生一定的影响。
研究表明,水稻扇型植硅体的鱼鳞状纹饰数量与水稻的驯化过程密切相关。统计分析显示,鱼山遗址河姆渡文化早期驯化型水稻植硅体占据39%,中期为47%,晚期达到59%;良渚晚期接近71%,商周和唐宋时期均在65%上下,呈现出了从早到晚逐渐增长并趋于稳定的变化趋势。马永超等对长江下游多个遗址的水稻扇型植硅体研究表明,上山文化水稻驯化型比例为34.18%~46.04%,跨湖桥文化接近58%,河姆渡文化早期约为35%,河姆渡文化晚期为52%,崧泽文化为69.33%,良渚时期达到74.4%,并据此推测水稻的驯化在良渚时期已经完成。郇秀佳等最新研究指出,长江中下游地区现代稻田中驯化型水稻扇型植硅体比例为57.6±8.7%,旱稻中这一比例只有11%左右。由此可知,鱼山遗址的水稻驯化在河姆渡文化晚期(5500-5100BP)发生了重大转变,驯化型比例基本达到了现代稻田的标准,预示着水稻驯化在此时基本完成。
(二)鱼山遗址各时期环境变化与水稻驯化
鱼山遗址孢粉、有机碳、海洋微体化石等多种代用指标研究表明,受极端气候事件和海水入侵的影响,该地河姆渡文化早晚期和良渚文化时期出现了多个文化间歇期。贺可洋等对鱼山遗址剖面的微体植物遗存研究认为河姆渡文化早晚期的中断和良渚文化的终止分别对应于距今6300-5600、5000-4500年的两次海水入侵。王张华等基于鱼山遗址多个剖面的环境代用指标分析指出,该地在2560BC年左右遭遇一次古台风事件,从而导致海平面上升和生态急速恶化。唐亮等研究揭示了鱼山遗址剖面记录了2次海水入侵事件,分别发生于6300-6000 cal.a B.P和4500-3400cal.a B.P。本文剖面植硅体、硅藻分析也证实了在河姆渡文化发展期间以及良渚文化时期前后均受到海平面变化和水环境的影响,从而对水稻驯化和稻作生产产生直接的影响。
依据高精度的测年数据,鱼山遗址河姆渡文化从距今约6800 年延续到距今约5100 年,持续的时间较长江下游核心区略长,结束的时段较晚。良渚文化晚期的地层堆积较浅,测年时间集中在距今约4900年到距今约4500年间,实际延续时间可能更短。贺可洋等对河姆渡遗址沉积钻孔研究指出该地在距今6400-5600年间因海侵导致了水稻驯化进程的间断并延长了驯化时间。同比之下,鱼山遗址更靠近海岸线,受海洋环境变化的频繁影响,水稻驯化进程具有一定的滞后性,驯化的周期也有所延长。
(三)鱼山遗址各时期生业模式变化
水稻的驯化进程与稻作的生产水平、先民生业模式密切相关。分析显示,河姆渡文化早、中期驯化型水稻植硅体的比例分别为39%和47%,水稻扇型植硅体的形态系数也处在动态变化中。炭化植物遗存显示的河姆渡早晚期水稻小穗轴驯化型比例从26.4%上升至32.3%,稻米的尺寸呈现逐渐增大的趋势。由此说明河姆渡文化早中期(6800-5800BP)是水稻驯化的快速发展期。基于该时期炭化水稻整体数量较少、水稻植硅体剖面含量较低、野生植物资源仍占有相当比例,表明处在驯化中的水稻尚未成为先民植物性资源的主体。河姆渡文化晚期驯化型水稻植硅体的比例上升至59%,已经达到现代稻田中驯化型水稻扇型植硅体的比例。从河姆渡文化早期至晚期,地层中水稻扇型植硅体的平均密度由3000/克干样增加到8600个/克干样,野生植物资源如菱、橡子、壳斗科植物的发现较少,表明该时期处在一个减少对野生植物资源的利用而转向依赖稻作农业的阶段。相距约200米的乌龟山遗址鱼类遗存的调查表明,河姆渡时期先民以利用乌鳢为主的淡水资源为主,同时利用多种近岸海鱼资源。河姆渡文化晚期长江下游近海的塔山遗址稳定同位素分析揭示,先民主要依赖包含C3类食物的稻作农业经济,可能辅以少量采集经济,肉食资源主要来自陆生系统的渔猎经济,部分来自家畜饲养。综合现有的研究成果和遗址的环境背景来看,鱼山先民水稻的驯化在河姆渡文化从早至晚绵延一千多年的过程中基本完成,稻作农业获得了较大发展;至河姆渡文化晚期,水稻逐渐成为先民植食性资源的主体,野生植物资源的利用在持续进行中;近海地区的淡水鱼和海鱼亦是肉食资源的重要补充。
良渚晚期,植硅体统计分析显示,驯化型水稻的比例达到74%,超越现代稻田表土的平均标准。这一时期地层堆积较单薄,炭化浮选仅发现11粒水稻和20粒小穗轴。大植物遗存发现大量属于一年生或多年生的水生草本植物种子,指示该地为池塘、沼泽或者湿地景观。微体植物遗存鉴定发现了大量指示水体环境的海绵骨针和硅藻,以及稗草、芒草、芦苇等湿地杂草植物。大植物和微体遗存鉴定证实了该地的植被体现了水田植物种群的特点,适宜滨海地区水稻的种植和栽培。同时期的浙江余杭茅山遗址、余姚田螺山遗址和施岙遗址均发现了相当规模的水稻田耕作系统,并利用植硅体分析和量化手段对良渚时期的稻田亩产进行了恢复。位于鱼山遗址东侧约1公里处的吕岙遗址于2021年的发掘中确定发现了大片良渚时期的水稻田遗迹层。囿于发掘区域的限制亦或地理环境的特殊性,目前尚无法从发掘层位上判断鱼山遗址水稻田的有无。但是综合鱼山遗址良渚时期出土的斧、锛、犁、刀、镰等多种生产工具和较高的水稻扇型植硅体密度,有理由推测鱼山遗址存在系统的水田耕作。长江下游地区多个遗址的动植物考古和人骨同位素研究证实,良渚文化的中心区存在先进的生业方式,即发达的稻作农业和以养猪为代表的家畜饲养。鱼山遗址处于良渚文化势力范围的边缘地带,水稻驯化完成并成为最重要的农作物资源,此时极大可能存在依靠湿地景观构建的稻田耕种系统。
商周时期,该地处在以越族为主体的古越文化圈和春秋末年越国所辖境内,其文化遗存与越文化密切相关。该时期植硅体分析显示水稻驯化完成,形态变化稳定。剖面中水稻植硅体百分比平均约7.5%,水稻扇型植硅体的平均密度高达3.5万个/克。大植物浮选中,水稻的出土數量和概率都有极大的提升,绝对数量占据该时期出土植物遗存的92.3%,未发现除水稻以外的农作物遗存。驯化稻的栽培种植已经成为了农业活动的主体,也是最重要的植食性食物来源。作为鱼山遗址文化堆积最丰厚的时期,考古发掘出土了布局清晰、功能明确的居住生活区及数量丰富的陶、瓷、青铜器等。遗物中包含诸多生产工具,如青铜斧、耨、锸、凿等,石器有斧、锛、刀、镰等,木器有木耙。形式多样的生产工具,满足了水田开垦、种植、管理、收获等一系列工作,极大地提高了农业生产力和集约化水平。长江下游地区在马桥文化时期(距今3900-3200年)和两周时期,普遍经历了偏干凉的环境和频繁的水患,农业生产明显衰退,稻作农业发展处于低谷,并在后期引入了粟、牛、马等北方文化因素。不同的是,鱼山遗址多种环境分析指示该地呈现出盐沼环境,海水影响减弱,淡水化趋势明显,植物生产力升高。植硅体分析也揭示商周时期以来是相对暖干的环境。与此同时,宁绍平原地区的遗址数量较前期也逐渐增多。上述分析综合表明了商周时期以来,鱼山先民偏安一隅,依托暖干的盐沼环境,持续发展稻作农业,为古越国的兴起与发展奠定了基础。
五、 结 语
1.结合鱼山遗址剖面高精度的植硅体分析,可对水稻驯化过程进行详细解读:水稻扇型个体从河姆渡时期至唐宋时期呈现了不断变小稳定的趋势;扇型个体逐渐向中间型和长柄型演化,推断鱼山先民以种植粳型化的水稻为主;水稻扇型植硅体鱼鳞状纹饰的变化揭示了该地区在河姆渡文化晚期,水稻的驯化基本完成。受海平面变化的频繁影响,该地的水稻驯化进程具有一定的滞后性,驯化的周期也有所延长。
2.综合水稻驯化进程和多种研究成果,推断在河姆渡文化早期至晚期,鱼山遗址稻作生产获得了较大发展,水稻逐渐成为先民植食性资源的主体,渔猎、采集和稻作并存的生业模式是早期先民适应滨海多变环境的结果。良渚晚期,伴随着水稻驯化的完成和生产工具的进步,推断出现了系统的水田耕作。商周时期,依托暖干的环境,稻作生产向集约化发展,为古越国的兴起奠定了基础。
3.鱼山遗址剖面的植硅体分析,为探讨沿海地带河姆渡文化早期至商周时期的水稻驯化过程与生业模式变化提供了更加翔实的证据,为深入了解滨海地区的文化演进、农业发展与人地关系提供了新的视角。
唐亮:《姚江-宁波平原中全新世海水入侵及古人类活动的快速响应》,华东师范大学博士学位论文,2020年。
郑云飞、蒋乐平:《上山遗址出土的古稻遗存及其意义》,《考古》2007年第9期。
郑云飞、陈旭高、王海明:《浙江嵊州小黄山遗址的稻作生产——来自植物硅酸体的证据》,《农业考古》2013年第4期。
Qiu Z., Jiang L., Wang C., et al., “New evidence for rice cultivation from the Early Neolithic Hehuashan site,” Archaeological and Anthropological Sciences, 2019, 11(4): 1259~1272.
郑云飞、蒋乐平、Crawford G.W:《稻谷遗存落粒性变化与长江下游水稻起源和驯化》,《南方文物》2016年第3期。
a. 赵志军:《中国稻作农业起源研究的新认识》,《农业考古》2018年第4期;b. Fuller D. Q., Qin L., Zheng Y., et al.,“The domestication process and domestication rate in rice: spikelet bases from the Lower Yangtze,” Science, 2009, 323(5921): 1607~1610.
王结华、丁友甫、雷少等:《浙江宁波镇海鱼山遗址Ⅰ期发掘简报》,《东南文化》2016年第4期。
郑云飞、孙国平、陈旭高:《全新世中期海平面波动对稻作生产的影响》,《科学通报》2011年第56卷第34期。
傅稻镰、秦岭、赵志军等:《田螺山遗址的植物考古分析——野生资源采集与水稻栽培、驯化的形态学观察》,《田螺山遗址自然遗存综合研究》,文物出版社,2011年。
潘艳:《长江三角洲与钱塘江流域距今10000-6000年的资源生产:植物考古与人类生态学研究》,复旦大学博士学位论文,2011年。
Ma Y., Yang X., Huan X., et al., “Rice bulliform phytoliths reveal the process of rice domestication in the Neolithic Lower Yangtze River region,” Quaternary International, 2016, 426: 126~132.
郑晓蕖、雷少、王结华等:《宁波鱼山遗址浮选结果及分析》,《农业考古》2019年第6期。
王永吉、吕厚远:《植物硅酸体研究及应用》,海洋出版社,1993年。
Lu H., Liu Z., Wu N., et al., “Rice domestication and climatic change: phytolith evidence from East China,” Boreas, 2002, 31(4): 378~385.
a. 同[11];b.Wu Y., Jiang L., Zheng Y., et al., “Morphological trend analysis of rice phytolith during the early Neolithic in the Lower Yangtze,” Journal of Archaeological Science, 2014, 49: 326~331.
郑云飞、陈旭高、丁品:《浙江余杭茅山遗址古稻田耕作遗迹研究》,《第四纪研究》2014年第1期。
a. 同[7];b. 雷少、王结华:《镇海鱼山·乌龟山遗址考古发掘——来自浙东滨海的远古回声》,《大众考古》2016年第2期。
陈书迁:《宁波鱼山遗址史前文化演进》,南京大学硕士学位论文,2016年。
a. 同[13];b. Piperno D. R., Phytolith: A Comprehensive Guide for Archaeologists and Paleoecologists. New York: Altamira Press, 2006.
a. He K., Lu H., Zheng Y., et al.,“Middle-Holocene sea-level fluctuations interrupted the developing Hemudu culture in the lower Yangtze River, China,” Quaternary Science Reviews, 2018, 188: 90~103;b. Wang Z., Ryves D. B., Lei S., et al., “Middle Holocene marine flooding and human response in the south Yangtze coastal plain, East China,” Quaternary Science Reviews, 2018, 187: 80~93.
王灿、吕厚远:《水稻扇型植硅体研究进展及相关问题》,《第四纪研究》2012年第2期。
a. 同[13];b.吴乃琴、吕厚远、聂高众等:《C3、C4植物及其硅酸体研究的古生态意义》,《第四纪研究》1992年第3期。
同[14]。
Zhao Z., Pearsall D. M., Benfer R. A., et al., “Distinguishing Rice (Oryza sativa Poaceae) from Wild Oryza Species through Phytolith Analysis, II: Finalized Method,” Economic Botany, 1998, 52(2): 134~145.
a. 同[21];b. Ball T., Chandler-Ezell K., Dickau R., et al., “ Phytoliths as a tool for investigations of agricultural origins and dispersal around the world,” Journal Archaeological Science, 2016, 68: 32~45.
a. 同[11];b.同[21]。
Piperno D. R., Phytolith Analysis: An Archaeological and Geological Perspective. London: Academic Press, 1988.
Fuller D.Q., Harvey E., Qin L., “Presumed domestication? Evidence for wild rice cultivation and domestication in the fifth millennium BC of the Lower Yangtze region,” Antiquity, 2007, 81(312):316~331.
a. 郇秀佳、李泉、馬志坤等:《浙江浦江上山遗址水稻扇形植硅体所反映的水稻驯化过程》,《第四纪研究》2014年第1期;b. 马永超:《长江下游地区的水稻驯化过程——水稻扇型植硅体的证据》,山东大学硕士学位论文, 2016年。
张文绪、王莉莉:《7个稻种叶片硅酸体的研究》,《中国农业大学学报》1998年第3期。
唐先干、谢金水、陈大洲等:《基于大样本数据的中国水稻扇型植硅体形态特征研究》,《古生物学报》2019年第4期。
a. Zheng Y., Dong Y., Matsui A., et al., “Molecular genetic basis of determining subspecies of ancient rice using the shape of phytoliths,” Journal of Archaeological Science, 2003, 30(10): 1215~1221; b. Wang C., Lu H., Zhang J., et al., “Bulliform phytolith size of rice and its correlation with hydrothermal environment: A preliminary morphological study on species in Southern China,” Frontiers in Plant Science, 2019: 1037; c. 郇秀佳、吕厚远、王灿等:《水稻扇型植硅体野生-驯化特征研究进展》,《古生物学报》2020年第4期。
同[11]。
a.同[32]c; b. Huan X., Lu H., Wang C., et al., “Bulliform phytolith research in wild and domesticated rice paddy soil in South China,”PLoS One, 2015, 10: e0141255.
同[20]a。
同[20]b。
同[1]。
He K., Lu H., Zheng H., et al., “Role of dynamic environmental change in sustaining the protracted process of rice domestication in the lower Yangtze River,” Quaternary Science Reviews, 2020, 242: 106456.
同[12]。
同[12]。
朱旭初、董宁宁、雷少:《宁波镇海乌龟山遗址出土鱼类遗存研究》,《南方文物》2020年第2期。
张国文、蒋乐平、胡耀武等:《浙江塔山遗址人和动物骨的C, N稳定同位素分析》,《华夏考古》2015年第2期。
同[12]。
a. 同[16];b. Zheng Y., Sun G., Ling Q., et al.,“Rice fields and modes of rice cultivation between 5000 and 2500 BC in east China,” Journal of Archaeological Science, 2009, 36(12):2609~2616; c. 曾毅:《浙江余姚发现世界上最大最早的“大规模水稻田”》,《光明日报》2020年12月16日第1 版。
本成果可见于“浙江考古”公众号,网址:https://mp.weixin.qq.com/s/CUdzhWHgrplbJIRPeLpZew。
a. 潘艳、袁靖:《新石器时代至先秦时期长江下游的生业形态研究 (下)》,《南方文物》2019年第1期; b. 袁靖、潘艷、董宁宁等:《良渚文化的生业经济与社会兴衰》,《考古》2020年第2期。
同[7]。
同[7]。
同[7]。
同[46]a。
同[1]。
同[1]。
