不同生物酶协同植物乳杆菌发酵对紫甘薯生全粉理化特性的影响
2023-10-20邹浩峰廖雨华黄师荣施建斌
邹浩峰, 廖雨华, 黄师荣, 隋 勇, 熊 添,施建斌, 蔡 沙, 蔡 芳, 梅 新
(湘潭大学化工学院1,湘潭 411105) (湖北省农业科学院农产品加工与核农技术研究所2,武汉 430064) (重庆文理学院园林与生命科学学院3,重庆 402168)
紫薯[Ipomoeabatats(L.)Lam]又名紫心甘薯、紫肉甘薯或黑薯,是甘薯的一个特殊品种,富含铁、锌、硒等多种矿物元素,含有18种氨基酸和多种维生素[1]。紫甘薯全粉是紫甘薯的主要深加工产品,基本保留了紫甘薯的营养价值和天然风味,是一种优质的食品原料。紫甘薯熟全粉是对新鲜紫甘薯进行熟化处理,制备过程比较繁琐,加热过程会使部分淀粉转化为还原糖,产品干燥效率较低,淀粉已基本糊化完全,会影响熟全粉的加工特性。紫甘薯生全粉制备工艺比较简单,原料没有经过熟化处理,淀粉含量较高,还原糖含量较低,细胞完整性较好,最终产品的持水、持油性、冻融稳定性和胶凝性等加工特性较紫甘薯熟全粉较好,此外,生全粉的糊化温度较低,热稳定性差,凝沉性强且不易老化[2]。
发酵是一种常见的食品加工方式,被用来提高食品的营养和风味。发酵过程可以产生多种高活性的酶,如糖苷酶、木聚糖酶、植酸酶等,可以将淀粉、蛋白质、纤维素等大分子物质水解为氨基酸、寡糖等小分子物质,有利于消化吸收,并增加食品中一些结合状态的功能成分如多酚等的释放[3],被广泛运用于如面包、馒头等淀粉类加工产品。发酵过程就是微生物生长繁殖过程,在此过程中微生物会向外界释放多种酶类物质,通过这些酶将环境中的大分子转化为易于吸收的小分子为其提供支撑生命活动的碳源和氮源[4]。Sun等[5]研究发现植物乳杆菌和酵母协同发酵使低分子量淀粉短链含量和结晶度下降,面团的抗回生能力、弹性和强度上升。Xu等[6]研究发现植物乳杆菌发酵使淀粉颗粒表面明显出现麻点和凹陷,增加了马铃薯淀粉的凝胶硬度和咀嚼性。王宏兹等[7]采用植物乳杆菌发酵紫薯粉制作的面包颜色鲜艳,具有独特芳香,比未添加紫薯粉的面包具有更高的抗氧化性和花青素含量。牛萌萌等[8]发现利用米根霉和乳酸菌混合发酵大麦仁发酵产品具有淡淡的酒香味,酸甜可口,抗氧化性也大幅提高。
目前研究主要是利用单一菌种如乳酸菌发酵淀粉,研究淀粉的理化特性和开发新产品,添加不同生物酶协同植物乳杆菌发酵的研究较少,特别是对紫甘薯生全粉的研究鲜有报道。因此研究使用紫甘薯生全粉为原料,利用植物乳杆菌(Lactobacillusplantarum)、植物乳杆菌+纤维素酶(Cellulase)+普鲁兰酶(Pullulanase)、米根霉(Rhizopusoryzae)、植物乳杆菌+纤维素酶+普鲁兰酶+米根霉4种方式发酵,研究不同发酵方式和时间对紫甘薯生全粉基本成分、理化性质和热特性的影响,以期为开发高质量的紫甘薯发酵产品提供参考。
1 材料与方法
1.1 材料与试剂
紫甘薯生全粉(日本凌紫);植物乳杆菌;米根霉;普鲁兰酶(1 000 U/g)、纤维素酶(15 000 U/g)、马铃薯葡萄糖琼脂、MRS琼脂、平板计数琼脂,BR;其余化学试剂均为分析纯。
1.2 仪器与设备
DSC 200 差示扫描量热仪,BCM-1000 生物洁净工作台,CF-100A 发酵箱,SHP-080 生化培养箱,YM-75 压力蒸汽灭菌器,GZX-9240 MBE电热鼓风干燥箱,SHA-B 水浴恒温振荡器,LGJ-25C 冷冻干燥机,UV-2800 紫外可见分光光度计,QJ-08 400 g 多功能粉碎机,TGL-24MC 台式高速冷冻离心机,FE20实验室pH计,SKD-08S2 红外智能消化炉,K9840自动凯氏定氮仪,TE214S 电子分析天平。
1.3 方法
1.3.1 发酵冻干粉的制备
植物乳杆菌发酵(Lac):在无菌条件下将冻干粉在MRS斜面培养基上37 ℃活化24 h,挑取单菌落于10 mL MRS液体培养基中进行二次活化,24 h后取1 mL菌液于MRS液体培养基,37 ℃振摇培养至菌液浓度为108CFU/mL。将所得菌液于4 ℃ 3 000 r/min条件下离心20 min,用生理盐水洗涤沉淀3次,将所得菌泥与无菌水按1∶10(质量体积比)的比例混匀后按紫甘薯生全粉质量的8%接种,水质量分数为71%。
植物乳杆菌+纤维素酶+普鲁兰酶发酵(Lac/Cel-Pul):将植物乳杆菌(质量分数为8%)、纤维素酶(30 U/g)和普鲁兰酶(20 U/g)混合均匀后接种于紫甘薯生全粉中。
米根霉发酵(Rhi):在无菌条件下于PDA平板培养基上划线接种米根霉,30 ℃条件下培养至长满孢子,用150 mL无菌水将孢子冲洗至250 mL锥形瓶后,振摇15 min后于无菌操作台上用纱布过滤,用100 mL无菌水冲洗纱布,即获得米根霉孢子悬浊液。将所得菌液按紫甘薯生全粉质量的8%接种。
植物乳杆菌+纤维素酶+普鲁兰酶+米根霉发酵(Lac/Cel-Pul/Rhi):将植物乳杆菌(质量分数为4%)、纤维素酶(30 U/g)、普鲁兰酶(20 U/g)和米根霉(质量分数为4%)混合均匀后接种于紫甘薯生全粉中。
参考徐忠等[9]的方法,稍作修改。将各菌液与紫薯生全粉混合均匀后于37 ℃、pH 6.0、85%湿度条件下发酵,选取发酵时间为3、6、9、12 h的样品,冻干后粉碎过100目筛保存备用。
1.3.2 基本成分测定
发酵紫甘薯生全粉水分、灰分、脂肪、蛋白质、淀粉、膳食纤维含量分别参考GB/T 5009.3—2016、GB/T 5009.4—2016、GB/T 5009.6—2016、GB/T 5009.5—2016、GB 5009.9—2016、GB 5009.88—2014。
可溶性糖含量参考王佩芬等[10]的方法,稍作修改。称取1 g发酵粉,加入10 mL 80%(体积分数)乙醇,60 ℃水浴振荡30 min,冷却后定容至50 mL。取滤液0.1 mL,以蒸馏水作空白,采用硫酸-蒽酮比色法测定。葡萄糖标准曲线为:y=14.058x-0.178 5,R2=0.999。式中:y为吸光度,x为葡萄糖质量浓度(mg/mL)。根据标准曲线计算可溶性糖含量。
1.3.3 pH和总酸度(TTA)的测定
参考王宏兹等[7]的方法。取1 g发酵粉,加入9 mL纯水,涡旋混匀后静置10 min,测定溶液的pH;以0.1 mol/L的NaOH溶液滴定该悬浊液pH为8.5,所消耗的NaOH的体积即为发酵粉的TTA值。
1.3.4 直链淀粉含量的测定
参考何洁等[11]的方法采用双波长法测定直链淀粉含量。
1.3.5 总酚的测定
参考杜俊娜等[12]的方法测定总酚含量。没食子酸标准曲线为:y=10.187x+0.040 3,R2=0.998 8。式中:y为吸光度,x为没食子酸质量浓度(mg/mL)。根据没食子酸标准曲线换算出总酚含量。
1.3.6 DPPH自由基清除能力的测定
参考王宏兹等[7]的方法测定DPPH自由基清除能力。
1.3.7 花青素含量的测定
参考Meng等[13]的方法测定花青素含量。
1.3.8 色泽测定
将样品平铺在平皿中,采用色彩色差计分别测定各样品的L*(明暗度)、a*(红绿色)、b*(黄蓝色)值。计算与未发酵样品的色差ΔE,计算公式为:
1.3.9 水/油吸附能力测定
参考Yousif等[14]的方法测定吸水/油性。
1.3.10 吸水膨胀性测定
参考郭亚姿等[15]的方法测定吸水膨胀性。
1.3.11 热力学特性分析
参照Pham等[16]的方法测定发酵粉热力学特性。称取(5.0±0.1)mg样品至铝坩埚中,加入10 μL去离子水,加盖密封,于室温下平衡2 h后采用DSC分析其热力学特性,空坩埚作为参比。测定参数为:以10 ℃/min升温速率从20 ℃升温至 120 ℃后以40 ℃/min速率从120 ℃迅速降至20 ℃,氮气流量20 mL/min,根据吸热曲线分析计算得糊化起始温度(To)、峰值温度(Tp)、终止温度(Tc)及糊化焓(ΔH)。
1.4 数据处理
所有实验均重复3次,结果以平均数±标准差表示;采用Excel和SPSS 26.0分析处理数据,Origin 2017绘图。
2 结果与分析
2.1 发酵对紫甘薯生全粉基本成分的影响
由表1可见,发酵使紫薯生全粉淀粉含量下降,植物乳杆菌参与的发酵淀粉含量下降更为明显,这可能是因为淀粉酶和有机酸加速了对淀粉的分解[17],Rhi发酵的淀粉水解程度更低,可能是因为其酸度变化最弱。植物乳杆菌参与的3种发酵方式的蛋白质含量升高,这可能是因为植物乳杆菌使用紫甘薯生全粉作为碳源和能源产生的真菌菌丝体蛋白使蛋白质含量升高[18]。发酵3 h后,Lac-Cel/Pul-Rhi脂肪含量显著降低,除Lac发酵外,其余发酵方式灰分含量均显著上升。乳酸菌能在一定程度上降解纤维素[19],并且在酶的作用下降解的更显著。Rhi发酵12 h后SDF含量要明显小于Lac/Cel-Pul/Rhi发酵,这表明米根霉和植物乳杆菌对膳食纤维有明显的协同作用,单纯的生物酶处理效果远逊于植物乳杆菌与生物酶共同作用[20]。

表1 发酵前后紫甘薯生全粉主要组成成分(干基)质量分数/%
由图1可见,4种发酵方式的可溶性糖含量在0~3 h内快速上升,在3 h后呈下降趋势。在0~3 h内植物乳杆菌和米根霉迅速繁殖,向环境内分泌大量的淀粉酶和糖化酶等活性物质,使淀粉含量下降,可溶性糖含量上升;3~9 h内,由于可溶性糖含量达到一定浓度且有机酸的积累开始抑制酶类物质的释放,植物乳杆菌和米根霉的生长速度减慢,部分可溶性糖被微生物消耗以在后期发酵中繁殖和代谢;9 h后,由于大量的有机酸等有害物质的积累,使得酶类物质的活性被严重抑制,植物乳杆菌等的生长能源只能依靠可溶性糖供给,因此可溶性糖含量快速下降。此外,多种菌混合发酵可溶性糖含量比单一菌种增加的更明显,这可能是因为真菌产生的淀粉酶、菊粉酶、纤维素酶等多种生物活性酶更多,以及大分子淀粉转化为更多的小分子糖有关[21]。
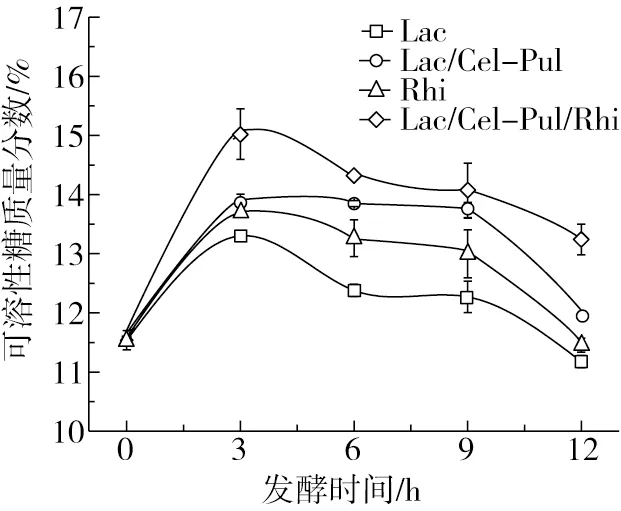
图1 发酵前后紫甘薯生全粉可溶性糖含量的变化
2.2 发酵对紫甘薯生全粉pH和TTA的影响
如图2所示,不同发酵方式的产酸能力差异性显著(P<0.05)。在发酵过程中生物酶能利用碳水化合物产生大量乳酸,使紫甘薯生全粉的pH降低,同时也能抑制有害微生物的生长繁殖[22]。4种不同发酵方式的紫甘薯生全粉在3 h内酸度明显上升,3~9 h内酸度呈缓慢上升趋势,在 9 h后酸度剧烈上升,pH的变化也与此现象相互吻合。在第12 h,除Lac/Cel-Pul/Rhi发酵外,其余发酵pH均在5.0以下,其中Lac发酵产酸能力最强。4种发酵方式pH在3 h内迅速下降,这是因为在初始发酵阶段生长环境适宜且营养丰富,使得植物乳杆菌和米根霉快速生长繁殖,产生大量乳酸。3~9 h内Lac/Cel-Pul、Rhi、Lac/Cel-Pul/Rhi发酵进入平稳期,pH缓慢下降,这可能是因为酶的添加促使紫甘薯中缓冲物质的释放,从而保证溶液体系的稳定。9 h后pH迅速下降是由于乳酸不断积累,逐渐破坏体系稳定性。不同发酵产酸能力 Lac/Cel-Pul>Lac>Lac/Cel-Pul/Rhi>Rhi,通过对比可以明显发现植物乳杆菌在发酵初期就成为优势主导菌,并且有酶参与的发酵组酸度下降更慢[20]。

图2 发酵对紫甘薯生全粉pH和酸度的影响
2.3 发酵对紫甘薯生全粉直链淀粉含量的影响
由表2可知,不同发酵方式使紫甘薯生全粉直链淀粉含量下降,但均无显著性差异(P>0.05),结合淀粉含量变化情况分析,虽然直链淀粉含量在下降,但直链淀粉与支链淀粉的比率在不断上升。有研究报道发酵过程中的微生物产生的酶和酸会水解支链淀粉,提高直链淀粉的比例,多种菌种混合发酵时优势微生物越多,产生的酶和酸就越多,直链淀粉增加的更明显[23]。

表2 发酵前后紫甘薯生全粉中直链淀粉质量分数(干基)/%
2.4 发酵对紫甘薯生全粉总酚含量和DPPH自由基清除率的影响
由图3可知,紫甘薯生全粉具有较高总酚含量,发酵使样品的总酚含量降低,Hector等[24]研究指出乳酸菌发酵降解了某些酚类物质,这与本研究结果一致。3 h后总酚含量有上升的趋势,可能是在发酵时产生的一些酶使细胞结构结合的多酚物质得以释放,从而提高总酚含量[25]。植物乳杆菌和米根霉单独发酵在12 h后总酚含量较为接近,但略低于混合菌发酵。不同发酵方式均会提高紫甘薯生全粉的抗氧化活性,发酵12 h时DPPH自由基清除能力大小为Lac/Cel-Pul>Lac>Lac/Cel-Pul/Rhi>Rhi,但差异不显著。有研究报道乳酸菌发酵会使一些酚类物质降解为抗氧化性更强的物质[7],从而提高其抗氧化能力。此外,可以明显看出米根霉参与的发酵DPPH自由基清除能力要略低一些,可能是因为米根霉的参与抑制了部分酚类物质降解。
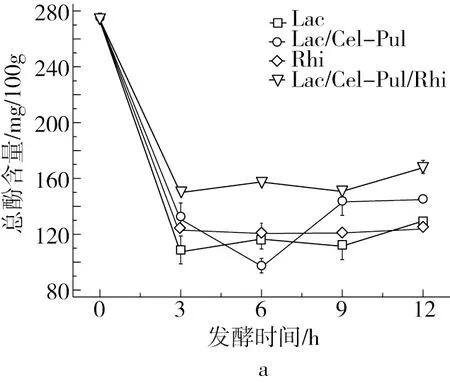
注:不同大写字母表示同一发酵时间不同发酵方式的差异显著(P<0.05),不同小写字母表示同一发酵方式不同发酵时间的差异显著(P<0.05),余同。
2.5 发酵对紫甘薯生全粉花青素含量的影响
如图4所示,不同发酵方式对花青素含量的影响显著。研究发现乳酸菌发酵对花青素有一定的降解作用[7],在本实验中的降解作用比较明显,0~3 h内所有发酵花青素含量剧烈下降。与总酚含量变化情况相同,添加米根霉的发酵对花青素的有影响较小,初步判断可能是米根霉的加入使植物乳杆菌分泌的一些微生物受到抑制,因此对花青素含量影响较小,但这需要进一步去研究论证。此外,随着发酵的进行,体系的pH降低,酸度变大,花青素的稳定性提高[26],因此几种发酵的花青素含量下降趋势变缓。Pretorius等[27]指出在发酵时一些微生物的次级代谢活动的产物可以与花青素结合成一种复合物,进而提高花青素物质的稳定性,这或许是花青素含量在发酵后期提高的原因。

图4 发酵对紫甘薯生全粉花青素含量的影响
2.6 发酵对紫甘薯生全粉色泽的影响
色差值越小证明与原样品颜色越接近,当色差值大于3时可以用肉眼分辨出来。由表3可见,所有发酵方式对紫甘薯生全粉的色泽影响较大,明显可以用肉眼分辨。随着发酵时间的延长,色差值有一个先上升后下降的过程,其中Lac发酵对紫甘薯生全粉的色泽影响最明显,发酵6 h时色差值达到最大。色泽变化情况可能与酸度变化有关,12 h时色差值变化情况为Lac>Lac/Cel-Pul>Lac/Cel-Pul/Rhi>Rhi,与pH变化情况一致(图2)。王宏兹等[7]指出主要花青素的显色会根据溶液pH的变化而变化,pH较低时为红色,随着pH的升高溶液逐渐变为浅紫色。
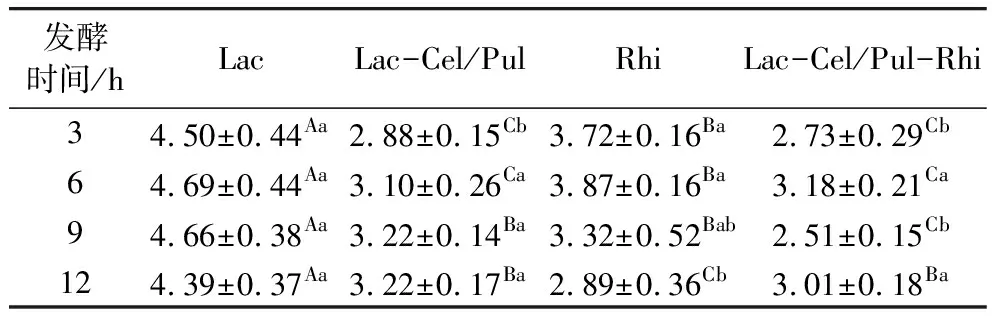
表3 发酵对紫甘薯生全粉色泽的影响
2.7 发酵对紫甘薯生全粉水合特性的影响
由图5可见,发酵后紫甘薯生全粉的持水率和吸水膨胀性显著下降,尤其是米根霉参与的发酵下降的更为明显,持油率随发酵时间的延长而增大,混合菌种的效果更明显。胡畔等[19]研究发现发酵使一些复合大分子在某些连接处断裂生成小分子,小分子的含量逐渐增大,而持水率的测定需要经过离心等操作,离心时小分子物质无法沉淀在底部而随上清液倒出,从而使发酵后玉米粉的持水率降低。混合菌的发酵加剧了此类小分子的生成,因此混合发酵的持水率、吸水膨胀性下降的更为严重。徐忠等[9]使用植物乳杆菌液态发酵玉米粉,发现持水率的降低和蛋白质、淀粉分子结构被破坏有关。持油率的上升可能是SDF含量的增加导致,Li等[28]研究发现发酵使SDF的比表面积增加,形成松散空洞的结构,更多非极性基团的暴露也可能使持油率上升。

图5 发酵对紫甘薯生全粉持水、持油、吸水膨胀性的影响
2.8 发酵对紫甘薯生全粉热力学特性的影响
由表4可见,与未发酵组相比,发酵组紫甘薯生全粉ΔH、Tc显著提高,To显著降低,Tp变化不显著,但有明显下降的趋势,特别是植物乳杆菌参与的发酵效果明显优于米根霉发酵,混合菌种的发酵效果显著优于单一菌种发酵。王东坤等[29]指出发酵过程产生的酶和酸能够水解或破坏淀粉的无定型区,水分子更易进入淀粉颗粒内部,从而使淀粉更易糊化,To降低。此外,由于淀粉的无定形区被破坏,其比例有所降低,结晶区比例相对增加,造成ΔH升高,Lac/Cel-Pul/Rhi发酵12 h时ΔH显著高于其余发酵。莫琰等[23]指出普鲁兰酶对淀粉无定形区和不完美结晶酶解后,剩余的完美结晶需要更高的温度才能熔融。此外,溶出的直链淀粉与脂类物质形成更为复杂的双螺旋结构,糊化所需的热量随之上升。直链淀粉相对含量上升可能会减弱淀粉结晶结构在糊化过程中受到的破坏,因此直链淀粉含量的增加可能也可能是造成ΔH升高的原因。
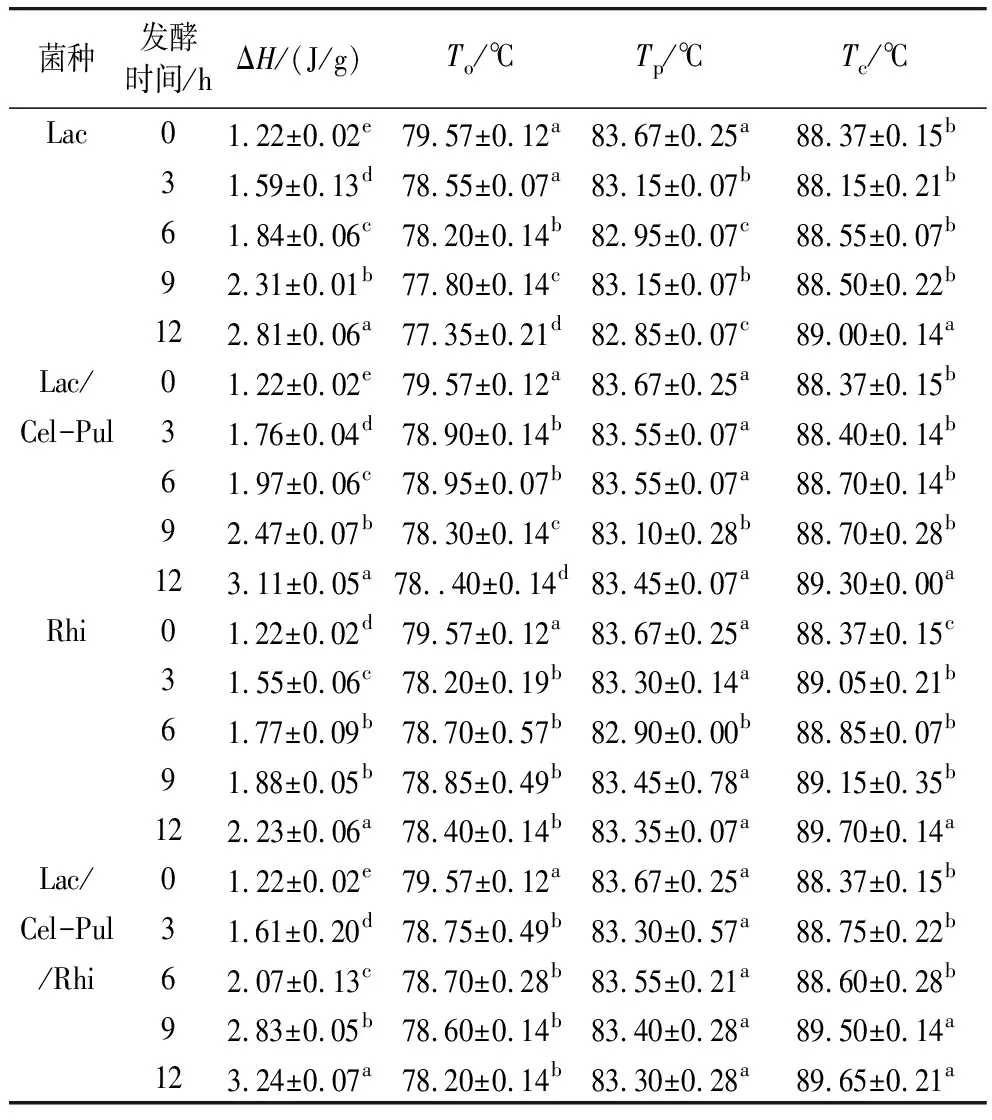
表4 发酵对紫甘薯生全粉热力学特性的影响
3 结论
研究不同生物酶协同植物乳杆菌发酵对紫甘薯生全粉基本成分、理化特性、色泽和热特性的影响。结果表明,Lac/Cel-Pul/Rhi发酵品质最好,SDF、可溶性糖、总酚、花青素、持油性明显高于其他发酵组,但持水性、吸水膨胀性较差,热特性结果显示其ΔH最高,To提前,Tc推迟,具有良好的热特性。紫甘薯生全粉颜色变化与产酸能力变化情况一致,由强到弱依次为Lac/Cel-Pul>Lac>Lac/Cel-Pul/Rhi>Rhi,这是因为花青素在不同pH条件下显色不同造成紫甘薯生全粉色泽的差异。此外,不同发酵方式均使紫甘薯生全粉抗氧化能力升高。不同生物酶协同植物乳杆菌发酵得到的紫甘薯生全粉品质较好,有利于发酵在紫甘薯生全粉工业的应用和高品质紫甘薯生全粉产品的开发。
