3种赤眼蜂对甜菜夜蛾和小菜蛾卵的寄生功能反应
2023-10-19王金彦季香云
王金彦,张 浩,季香云
(上海市农业科学院生态环境保护研究所,上海市设施园艺技术重点实验室,农业农村部东南沿海农业绿色低碳重点实验室,上海 201403)
甜菜夜蛾SpodopteraexiguaHübner和小菜蛾PlutellaxylostellaLinnaeus均为世界性分布的害虫,主要为害十字花科植物,给农业生产造成了巨大的损失(李振宇等,2011;Jiangetal.,2013;王金彦等,2018;Wangetal.,2020,2021a)。目前,对于甜菜夜蛾和小菜蛾的防控,一直以化学防治为主。但是化学农药大量不规范的使用,严重影响了生态环境安全和农产品质量安全,也导致这两种害虫的抗药性日趋增强。因此,采用可持续的生物防控方法防治甜菜夜蛾和小菜蛾尤为重要。目前,在甜菜夜蛾和小菜蛾的生物防治方面,病原微生物、捕食性天敌和寄生性天敌(茧蜂)的应用已有较系统的研究(Jiangetal.,2011;季香云等,2013;周今一等,2016;时敏等,2020;Wangetal.,2021b),例如:核型多角体病毒Nucleopolyhedrovirus(蒋杰贤等,2005;金晶等,2021)、益蝽Asopinae(高卓,2010;唐艺婷等,2020)、绒茧蜂Apanteles(黄芳等,2014)等对甜菜夜蛾或小菜蛾均有一定的防控效果。但仍需要发掘更多生物防治资源用于甜菜夜蛾和小菜蛾的防治,确保蔬菜和粮食安全生产。
赤眼蜂Trichogramma,属膜翅目Hymenoptera赤眼蜂科Trichogrammatidae,作为一种卵寄生蜂,其控害作用能力强,寄主范围广,是国内外害虫生物防治中应用最广的天敌昆虫之一(向玉勇和张帆,2011;Koetal.,2014;李姝等,2018;Zangetal., 2021)。近年来,赤眼蜂被广泛用于防治水稻二化螟ChilosuppressalisWalker、稻纵卷叶螟CnaphalocrocismedinalisGuenée、松毛虫DendrolimuspunctatusWik、玉米螟PyraustanubilalisHübner等农林害虫(刘万学等,2003;陈洪凡等,2010;郭震等,2011;宋静等,2015)。目前研究发现稻螟赤眼蜂TrichogrammajaponicumAshmead,螟黄赤眼蜂TrichogrammachilonisIshii和玉米螟赤眼蜂TrichogrammaostriniaePangetChen均可寄生甜菜夜蛾卵和小菜蛾卵(舒锐豪等,2014;许晶,2017;刘兴龙等,2018;冷春蒙等,2020)。但3种赤眼蜂对甜菜夜蛾和小菜蛾的寄生功能反应尚未见报道。因此,本研究在实验室条件下评价了稻螟赤眼蜂、螟黄赤眼蜂和玉米螟赤眼蜂对不同密度甜菜夜蛾卵和小菜蛾卵的寄生效应,明确赤眼蜂对甜菜夜蛾和小菜蛾的控害潜能,旨在为推广和高效应用赤眼蜂防控甜菜夜蛾和小菜蛾提供依据。
1 材料与方法
1.1 供试赤眼蜂
稻螟赤眼蜂由浙江省农业科学院提供,螟黄赤眼蜂和玉米螟赤眼蜂来源于山东省农业科学院植物保护研究所天敌与授粉昆虫研究中心。3种赤眼蜂均在上海市农业科学院生态保护研究所养虫室内,用米蛾卵在室内温度25±1℃、相对湿度75%±5%、光周期L∶D=14 h∶10 h条件下繁殖多代后用于本试验。
1.2 供试寄主卵
甜菜夜蛾卵块和小菜蛾卵块均采集于上海市农业科学院庄行综合试验站(30°53′25.93″ N,111°22′49.93″ E)甘蓝试验田。将卵块置于人工气候培养箱(28℃、14 L∶10 D、RH 80%±5%)中洁净的塑料盒(直径15 cm、高10 cm)内,待卵块孵化后,饲喂人工饲料。将羽化的甜菜夜蛾或小菜蛾雌雄成虫配对后分别置于圆柱形产卵筒(直径12 cm、高23 cm)中。甜菜夜蛾产卵筒中放置白色纸卡,小菜蛾的产卵筒中放置透明塑料纸供其产卵,并在筒底放置含10%蜂蜜水的脱脂棉球为其补充营养。收集甜菜夜蛾或小菜蛾当天产的虫卵供试验用(Ramalhoetal.,2018)。
米蛾CorcyracephalonicaStainton卵由浙江省农业科学院提供。在养虫室内温度25±1℃、相对湿度75%±5%、光周期L∶D=14 h∶10 h条件下用玉米粉和麦麸混合而成的饲料饲养米蛾幼虫。成虫收集至用40目不锈钢网做成的养虫笼中。用毛刷收集养虫笼壁上的米蛾卵,单层平铺在粘性纸上制成卵卡,于30 W紫外灯下杀胚30 min供赤眼蜂寄生。
1.3 赤眼蜂对不同密度寄主卵的寄生功能反应
选取合适的卵块,连同产卵纸一同剪下,最终形成相似大小的卵卡(甜菜夜蛾卵卡大小约为1.5 cm×1.5 cm;小菜蛾卵卡大小约为3 cm×1.5 cm)。在平底玻璃管(直径2 cm、高15 cm)中分别接入单层附着有10、20、40、60、80粒的新鲜甜菜夜蛾或小菜蛾卵的卵卡,每管引入1头当日羽化并已充分交配12 h的赤眼蜂(稻螟赤眼蜂或螟黄赤眼蜂或玉米螟赤眼蜂)雌蜂,并在玻璃管壁内放置含20%的蜂蜜水的脱脂棉球为雌蜂提供食物,棉塞封口。将其置于温度为25±1℃,RH 70%±5%,光周期14 L∶10 D的人工气候箱中,寄生24 h后引出雌蜂。甜菜夜蛾卵和小菜蛾卵在人工气箱中继续培养,及时移除未被寄生的孵化的幼虫。待虫卵变黑且幼虫不再孵化后,将其置于在解剖镜下解剖、观察并统计被寄生的卵量(陈万斌等,2020)。每个处理重复20次。用HollingⅡ模型方程拟合寄生功能反应数据,模型为:Na=aTN/(1+aThN)(Holling,1959),式中,Na为被寄生的卵量,a为瞬间攻击率,T为寄生时间(d),N为供寄生蜂寄生的卵粒数,Th为赤眼蜂处理一粒卵所需的时间。
1.4 赤眼蜂对不同密度寄主卵的的搜寻效应
根据1.3所得a、Th和N的值拟合Holling(1959)所提出的寻找效应(S)与寄主密度的模型方程:S=a/(1+aThN)。
1.5 数据处理
试验数据采用SPSS 23.0和Sigmaplot 14.0软件进行分析及制图。对3种赤眼蜂的寄生率和寄生量进行单因素方差分析,应用Tukey’s HSD法检验处理间差异的显著性。
2 结果与分析
2.1 3种赤眼蜂对寄主卵的寄生率
3种赤眼蜂对甜菜夜蛾和小菜蛾卵的寄生率的大小分别表现为:螟黄赤眼蜂>稻螟赤眼蜂>玉米螟赤眼蜂(图1),稻螟赤眼蜂>螟黄赤眼蜂>玉米螟赤眼蜂(图2)。对于甜菜夜蛾卵,当寄主密度为10粒时,螟黄赤眼蜂对甜菜夜蛾的寄生率显著高于玉米螟赤眼蜂(F=4.456,df=2, 57,P<0.05);当寄主密度为20粒时,螟黄赤眼蜂的寄生量显著高于稻螟赤眼蜂和玉米螟赤眼蜂(F=6.553,df=2, 57,P<0.05);3种赤眼蜂在寄主密度为40、60和80粒时的寄生率没有显著性差异。在5个不同寄主密度时,稻螟赤眼蜂对小菜蛾卵的寄生率均显著高于螟黄赤眼蜂和玉米螟赤眼蜂(P<0.05);仅当寄主密度为20粒/管时,螟黄赤眼蜂对小菜蛾卵的寄生能力显著高于玉米螟赤眼蜂(P<0.05)。

图1 3种赤眼蜂对甜菜夜蛾卵的寄生情况Fig.1 Parasitism rate of three Trichogrammas onSpodoptera exigua egg注:图中不同小字母表示显著性差异(P<0.05)。Note: Different small letters in the figure indicated significant difference (P<0.05).
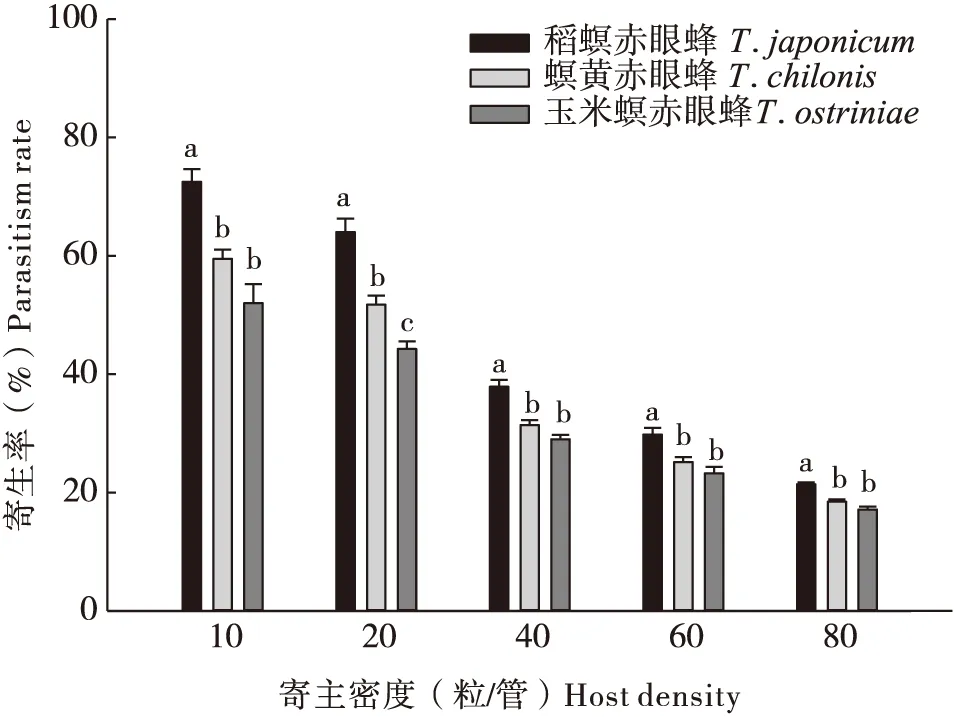
图2 3种赤眼蜂对小菜蛾卵的寄生情况Fig.2 Parasitism rate of three Trichogrammas onPlutella xylostella egg注:图中不同小字母表示显著性差异(P<0.05)。Note: Different small letters in the figure indicated significant difference (P<0.05).
2.2 3种赤眼蜂对两种寄主卵的寄生功能反应
在试验设定范围内,3种寄生蜂的寄生量随寄主密度的增加而显著升高(P<0.05)(表1、表2)。当甜菜夜蛾卵的密度大于20粒时,寄生蜂对甜菜夜蛾的寄生量趋于平稳(图3);当寄主密度大于40时,寄生蜂对小菜蛾的寄生量不再显著增加(表2、图4)。数据采用HollingⅡ圆盘方程进行拟合,发现3种赤眼蜂对两种寄主卵的寄生量均符合该模型(表3、表4)。判定系数R2均高于0.8,说明HollingⅡ模型能很好地反应3种赤眼蜂对甜菜夜蛾和小菜蛾卵的寄生作用。3种赤眼蜂对甜菜夜蛾卵的瞬间攻击率和寄生效能由高到低依次为:螟黄赤眼蜂>稻螟赤眼蜂>玉米螟赤眼蜂;处理时间与之相反,表现为:玉米螟赤眼蜂>稻螟赤眼蜂>螟黄赤眼蜂(表3)。稻螟赤眼蜂对小菜蛾的瞬间攻击率和寄生效能最高,处理时间最短,螟黄赤眼蜂次之,玉米螟赤眼蜂最低(表4)。
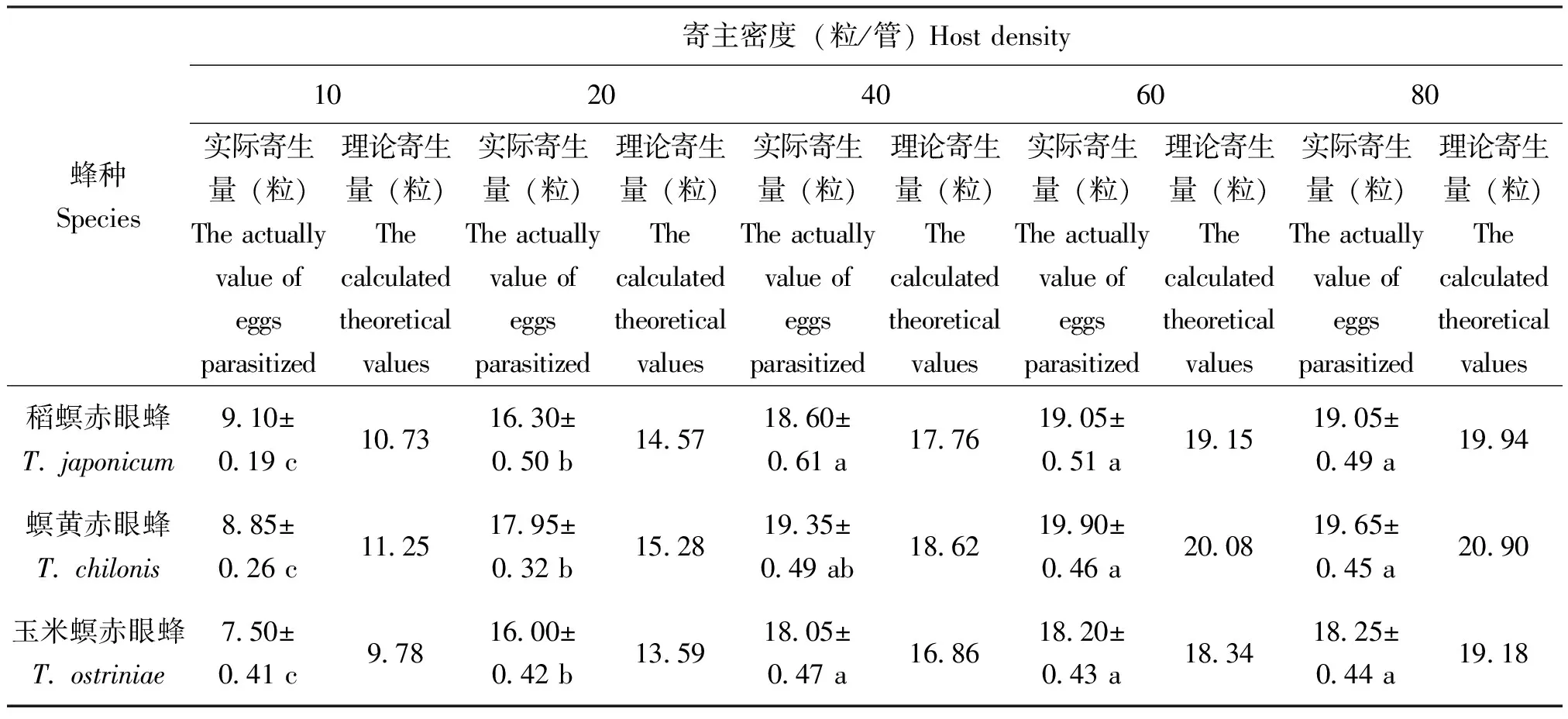
表1 3种赤眼蜂对甜菜夜蛾卵的寄生量
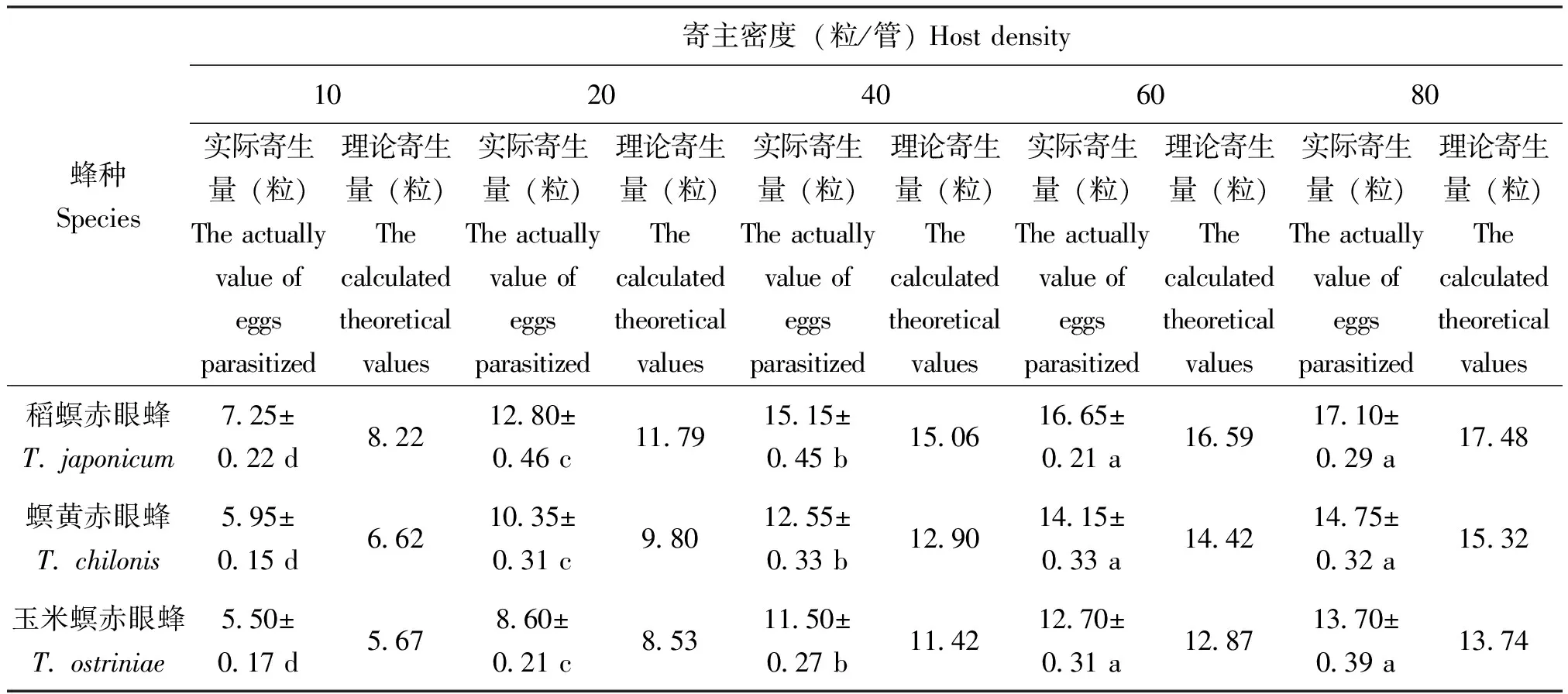
表2 3种赤眼蜂对小菜蛾卵的寄生量

表3 3种赤眼蜂对甜菜夜蛾卵的寄生功能反应参数
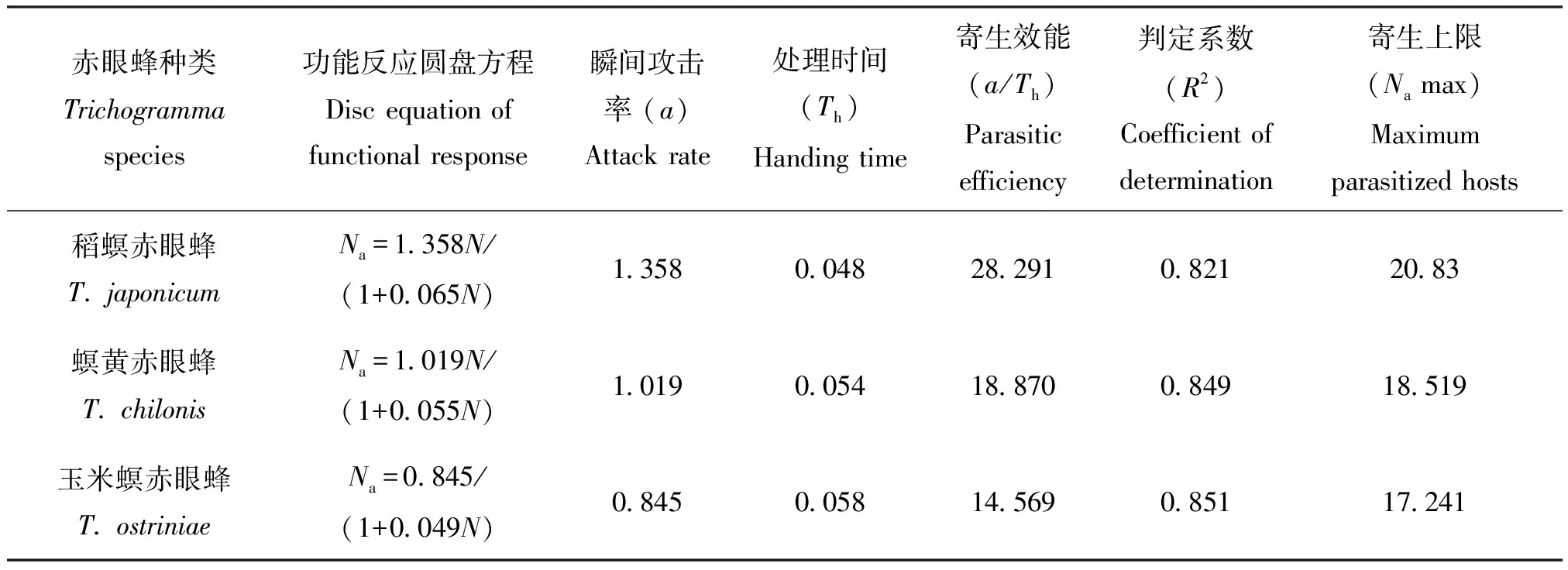
表4 3种赤眼蜂对小菜蛾卵的寄生功能反应参数
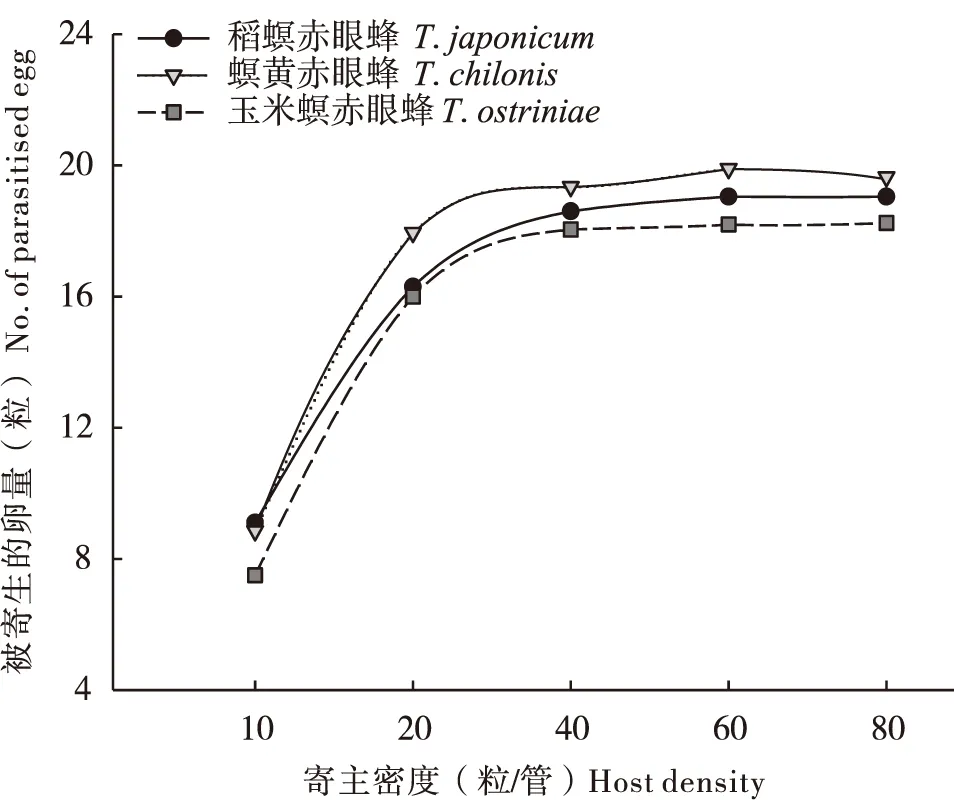
图3 3种赤眼蜂对甜菜夜蛾卵的寄生功能反应曲线Fig.3 Functional responses curves of three Trichogrammaon Spodoptera exigua egg

图4 3种赤眼蜂对小菜蛾卵的寄生功能反应曲线Fig.4 Functional responses curves of three Trichogrammaon Plutella xylostella egg
2.3 3种赤眼蜂对两种寄主卵的搜寻效应
相同条件下,螟黄赤眼蜂、稻螟赤眼蜂和玉米螟赤眼蜂对甜菜夜蛾卵和小菜蛾卵的搜寻效应均随寄主密度的增加而降低,其中螟黄赤眼蜂对甜菜夜蛾的搜寻效应最高,稻螟赤眼蜂对小菜蛾的搜寻效应最高,玉米螟赤眼蜂对2种寄主的搜寻效应均最低(图5)。

图5 3种赤眼蜂的搜寻效应与甜菜夜蛾卵密度的关系Fig.5 Relationship between the searching efficiency of the three Trichogramma and Spodoptera exigua egg
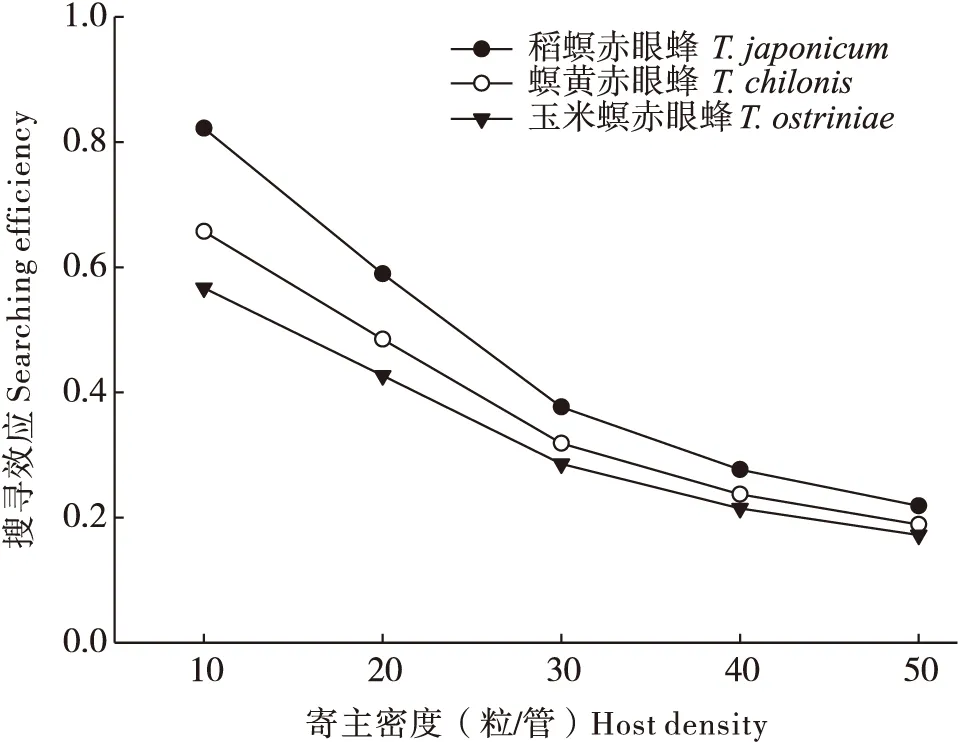
图6 3种赤眼蜂的搜寻效应与小菜蛾卵密度的关系Fig.6 Relationship between the searching efficiency of the three Trichogramma and Plutella xylostella egg
3 结论与讨论
功能反应不仅描述了害虫天敌与害虫种群数量动态关系,也是评价天敌对害虫寄生或捕食效率的重要指标(Solomon,1949;Jonesetal.,2003;Farrokhietal.,2010)。在功能反应的相关研究中,Holling提出的圆盘方程是最早被广泛用于评估天敌对其猎物对象作用大小的模型之一,其中HollingⅡ模型方程已被证实是适用于寄生蜂功能反应研究的主要模型(黄寿山,等1993;王甦,2014;王金彦等,2020)。本研究也发现3种赤眼蜂(稻螟赤眼蜂、螟黄赤眼蜂和玉米螟赤眼蜂)对甜菜夜蛾和小菜蛾卵的寄生量均符合HollingⅡ模型。3种赤眼蜂的寄生量随寄主密度的增加而增加,当上升到一定程度后,寄生量增幅减缓并趋于平稳,这可能与寄生蜂体内有限的成熟卵量有关。在功能反应方程中,寄生效能是反映寄生蜂对寄主控制效果的重要参数(陈俊谕等,2013),a/Th值越大,说明控制效果越好。总体而言,由于物种之间生理特性、行为因素等差异,不同种赤眼蜂对寄主的寄生效能存在差异。何余容等(2002)的研究结果表明拟澳洲赤眼蜂TrichogrammaconfusumViggiani对小菜蛾卵的a/Th值为0.0169,大于短管赤眼蜂TrichogrammapretiosumRiley对小菜蛾卵的a/Th值0.0106。这与本研究发现的螟黄赤眼蜂对甜菜夜蛾的寄生效能最高,稻螟赤眼蜂对小菜蛾卵的寄生效能高于螟黄赤眼蜂和玉米螟赤眼蜂研究结果相符合。
搜寻效应反映寄生蜂寻找猎物的能力。本研究发现,螟黄赤眼蜂对甜菜夜蛾的搜寻能力最强,稻螟赤眼蜂对小菜蛾有较高的寻找效应,说明赤眼蜂对不同猎物的搜捕能力有一定差异。并且,3种赤眼蜂对甜菜夜蛾或小菜蛾的搜寻效应均随寄主密度的增加而降低,说明寄主密度对赤眼蜂搜寻有干扰作用。这与陈俊谕等(2013)以及陈万斌等(2020)报道的随寄主密度的增加,寄生蜂搜寻时间减少和搜寻效应下降的结果一致。因此在田间应用赤眼蜂防控甜菜夜蛾时,应当根据赤眼蜂种类和寄主密度合理释放赤眼蜂,在节约成本的同时达到最好的防控效果。
赤眼蜂在害虫生物防治的应用中,引种和释放前的科学评价不仅能够避免盲目引种导致资源浪费和防治的失败,并且引用优势蜂种还可以达到更好的控害效果。综合本研究结果,3种赤眼蜂对甜菜夜蛾和小菜蛾均有一定的防控作用。相对稻螟赤眼蜂和玉米螟赤眼蜂,螟黄赤眼蜂更适用于甜菜夜蛾的防治,而稻螟赤眼蜂对于小菜蛾的防控能力优于螟黄赤眼蜂和玉米螟赤眼蜂,可作为小菜蛾防治的优势赤眼蜂种进行进一步研究。这与舒锐豪等(2014)报道的稻螟赤眼蜂以小菜蛾卵作为中间寄主的适合度高于螟黄赤眼蜂类似。而谌江华等(2016)的研究发现螟黄赤眼蜂对小菜蛾卵的寄生率要高于稻螟赤眼蜂,这可能与赤眼蜂地理隔离、试验环境差异等因素有关。由于野外环境中其他竞争者、捕食者、温度、湿度、降水等不可控因素较多,对赤眼蜂的寄生效应有不同程度的影响,因此,在应用赤眼蜂防控甜菜夜蛾和小菜蛾时,还需要结合实际的田间环境进一步探索和完善应用方案。
