日本蚱的生活史及其它生物学特性
2023-10-19张荣娇王缉广黄丹玲邓维安
张荣娇,王缉广,黄丹玲,邓维安
(1. 河池学院化学与生物工程学院,广西宜州 546300;2. 广西师范大学生命科学学院,珍稀濒危动植物生态与环境保护教育部重点实验室,广西珍稀濒危动物生态重点实验室,广西桂林 541006)
日本蚱Tetrixjaponica隶属于直翅目Orthoptera蚱科Tetrigidae昆虫,主要分布于中国、日本、朝鲜和俄罗斯等东亚地区(邓维安等,2007)。日本蚱的鉴别特征:头部不突出于前胸背板之上,头顶宽为一眼宽的1.5倍,颜面隆起与头顶成钝角形。前胸背板前缘平直,侧面观上缘近平直,前段略呈屋脊形。前后翅正常。前、中足股节下缘直或略波状,中足股节的宽度明显大于前翅能见部分的宽度。目前,日本蚱的鉴定主要根据成虫的外部形态特征,尚未见有日本蚱若虫的形态特征、生活史及其生物学特性研究的相关报导。
昆虫生活史,是指昆虫在一定时间阶段中的生长、发育、行为的周期变化(彩万志,2001)。通过对昆虫生活史研究,可以掌握该种昆虫在一年中的发生代数,各虫态的出现时间与历期,及其生物学特性等情况。近年来,由于生物防治技术在农业上的广泛运用,昆虫生活史及其生物学特性的研究愈发受到人们的重视。然而,关于蚱类昆虫的研究报道极少,肖舒晴等对眼优角蚱Eucriotettixoculatus进行室内饲养,研究其生活史及相关生物学特性(肖舒晴等,2019)。此外,结合形态学和生物学特征,部分学者还对蚱类昆虫的前胸背板与翅型的多态性进行了探讨,如物种分类鉴定(Deng, 2021;Dingetal., 2021)、多态性的发生与季节及其栖息环境关系等(Pushkar, 2009;Tsuruietal., 2010;Lehmannetal., 2018)。
蚱类昆虫属于地栖性昆虫,其分布与环境植被密切相关,有些种类对环境变化非常敏感,只能生活在特定的环境中;而有些种类的适应能力却很强,广泛分布于多种环境中,如日本蚱在荒坡草丛、山谷草地、灌木丛、阔叶林和针叶林等多种环境的地表均可生活,适应能力强,具有多态性,是研究昆虫生态适应性的理想物种。本研究以日本蚱为对象,对其生活史及相关生物学特性开展研究,分析日本蚱在自然环境下各龄若虫的形态变化规律及其生物学特性,对研究昆虫的生活史及其生物学特性具有一定的理论意义,进而对了解物种形成和生态适应性,维护生态平衡以保护环境也具有一定的实践意义。
1 材料与方法
1.1 野外的取样
供试日本蚱采自广西河池市宜州区,在春季(3-5月),夏季(6-8月),秋季(9-11月),冬季(12月-翌年2月)中,每月定点到日本蚱栖息地进行观察,记录日本蚱在野外生活环境、取食、卵及各虫龄的形态特征、生活习性等生物学特征,使用温湿光记录仪(GM8910)测量环境中的温度、湿度、光照强度等环境因子。而后将日本蚱的成虫带回实验室,经过鉴定后,模拟日本蚱在野外生存的条件,以栖息地的苔藓与腐殖质等为食物,利用智能人工气候室(MC-RG2019PA)对日本蚱进行饲养。
1.2 室内饲养方法
利用智能人工气候室(MC-RG2019PA)模拟野外日本蚱的生活环境,再以昆虫饲养盒(长27 cm×宽15 cm×高17 cm)为主体,在盒底铺上一层从栖息地带回的松软泥土,并在土上铺长满苔藓的土表层,每天早、中、晚各往盒内喷水一次,防止苔藓枯死以及保持盒内湿度。将野外带回的日本蚱越冬成虫,按比例(3♀∶5♂)放入饲养盒中饲养;在饲养过程中,定期投放新鲜苔藓、枯枝落叶等腐殖质食物,及时清理蚱的粪便,观察其交配、产卵。卵孵化后饲养若虫,连续饲养3代,记录其取食、卵及各虫龄的形态特征、生活习性等生物学特征。
1.3 生活史及其习性观察方法
在野外观察与室内饲养过程中,使用照相机(Canon G5X)记录雌性产卵时的行为。每天定时对其所产的卵块拍照,观察孵化前卵块的变化;待卵块孵化时,对孵化过程录像并记录孵化时间,计算卵发育历期。初孵化的若虫,用体式显微镜(Model JSZ8)观察后,再用基恩士超景深数码相机(KEYENCE VHX-6000)拍照与测量,随后用记号笔标记,分盒分批饲养。记录若虫每一个虫龄的蜕皮时间、每次蜕皮过程所需的时间,以及若虫期间蜕皮次数等,直到羽化为成虫,并对羽化过程录像;观察并记录各虫龄的形态特征、若虫及成虫的主要生活习性等。
2 结果与分析
2.1 卵块
日本蚱卵块的形状近似橘瓣状,卵粒数目通常为10~30粒;卵粒的大体形状似大米粒,外围的卵粒稍微向卵块中心弯曲,卵块近中心的卵粒轻微向卵块中心弯曲,卵块正中心的卵粒外形近似长卵形;卵粒紧紧粘附成一团形成类似橘瓣状结构,卵块纵切面似扇形;刚产出的卵粒的颜色是橙色(图1-A)或者柚红色(图1-B),外表整体稍微呈透明状态;卵块24 h后开始出现白色絮状保护膜,而后卵块外保护膜消失,卵块更容易松散,颜色由橙色或者柚红色变为褐色(图1-C)。卵粒随着发育会逐渐变大膨胀,并且各卵粒逐渐分开,卵粒端部有2个圆形黑色小区域(即复眼),表明卵即将孵化。卵块中心的卵粒相对卵块外围的卵粒细长,越往外卵粒越弯且粗短。卵粒长度与直径的平均值分别为2.069 mm、0.585 mm。

图1 日本蚱的卵块Fig.1 Egg mass of Tetrix japonica注:A,刚产的橙色卵块;B,刚产的柚红色卵块;C,变色后的褐色卵块。Note: A, Orange egg mass just laid; B, Grapefruit red egg mass just laid; C, Brown egg mass after discoloration.
2.2 若虫的形态特征
若虫龄的划分以其蜕皮作为标准。刚开始孵化为1龄,而后蜕皮一次就增长一龄;蜕皮后,其大小和形态与前一个虫龄相比,会存在一定的差异,雄性共有6龄,其各龄若虫及成虫(短背短翅型)的形态特征见表1和图2;雌性共有7龄,其各龄若虫及成虫(短背短翅型)的形态特征见表2和图3。在雄性与雌性若虫羽化前,均无翅凹与膝前齿;在羽化为成虫后,前胸背板侧片后缘出现翅凹,前翅发育成熟且显现在翅凹处,后足股节出现膝前齿,后翅的翅脉出现横脉。
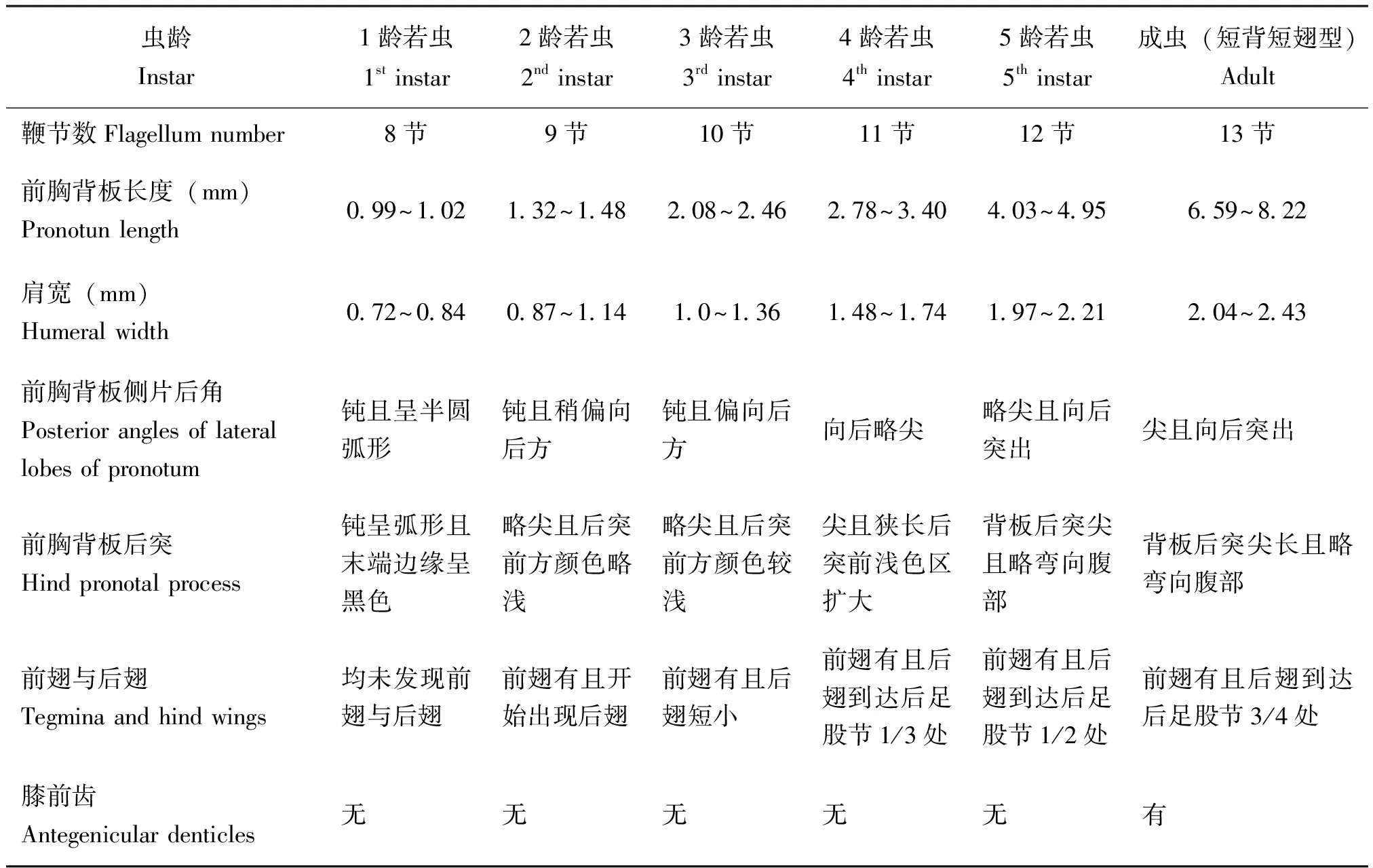
表1 日本蚱的雄性各龄形态特征表

表2 日本蚱的雌性各龄形态特征

图2 日本蚱的雄性若虫及成虫Fig.2 Nymphs and adult of Tetrix japonica male注:A,1龄雄性若虫;B,2龄雄性若虫;C,3龄雄性若虫;D,4龄雄性若虫;E,5龄雄性若虫;F,雄性成虫。标尺=1 mm。Note: A, 1st instar male nymph; B, 2nd instar male nymph; C, 3rd instar male nymph; D, 4th instar male nymph; E, 5th instar male nymph; F, Male adult. Scale bars=1 mm.

图3 日本蚱雌性若虫及成虫Fig.3 Nymphs and adult of Tetrix japonica female注:A,1龄雌性若虫;B,2龄雌性若虫;C,3龄雌性若虫;D,4龄雌性若虫;E,5龄雌性若虫;F,6龄雌性若虫;G,雌性成虫。标尺=1 mm。Note: A, 1st instar female nymph; B, 2nd instar female nymph; C, 3rd instar female nymph; D, 4th instar female nymph; E, 5th instar female nymph; F, 6th instar female nymph; G, Female adult. Scale bars=1 mm.
触角的鞭节数也可作为若虫龄划分的一个重要依据。1龄若虫鞭节为8节,而后若虫每增长一龄,其鞭节数也会相应的增加一节;每一龄若虫在该龄的末期,鞭节的第一亚节开始出现分节现象,但未形成新的亚节,当蜕皮后,鞭节的亚节数增加一节,如1龄初期触角的鞭节为8节,末期鞭节第一亚节开始出现分节现象,但未形成新的亚节,当刚蜕皮进入2龄时,鞭节的亚节数明显增加一节变为9节;其他龄若虫以此类推。雄性羽化后鞭节为13节;雌性由于比雄性多1龄,羽化后鞭节为14节。
2.3 生活史
通过对栖息于广西河池市宜州区的日本蚱的长期野外观察和室内实验种群的观察,日本蚱一年发生两代,常可见以成虫越冬和以卵越冬同时出现的现象。以成虫越冬的日本蚱生活史:越冬成虫翌年4月开始活动,5月上旬开始交配,5月中旬至7月上旬为交配产卵的高峰期,交配产卵会持续到8月上旬,6月上旬至8月下旬陆续孵化出若虫。越冬成虫于5月下旬至8月下旬陆续死亡,新一代若虫于7月下旬至9月下旬羽化为成虫,于10月下旬至11月上旬陆续进入冬眠状态。以卵越冬的日本蚱生活史:越冬虫卵翌年4月开始孵化,5月至6月中旬为发育期,6月下旬至7月上旬羽化为成虫,于7月中旬至下旬为交配产卵期,新一代若虫于8月至9月孵化为若虫期,10月成虫交配产卵期,产卵后成虫死亡,以卵越冬。
日本蚱的雄性和雌性,卵发育期相同,通常为6~10 d。日本蚱的雄性,若虫在整个发育过程中一共蜕皮5次,历期长达37~42 d,如1龄若虫期通常为6~7 d;2龄若虫期为6~7 d;3龄若虫期为7~8 d;4龄若虫期为8~9 d;5龄若虫发育历期最长,为10~11 d。日本蚱的雌性,若虫在整个发育过程中一共蜕皮6次,历期长达45~50 d,如1龄若虫期通常为6~7 d;2龄若虫期为6~7 d;3龄若虫期为7~8 d;4龄若虫期为7~8 d;5龄若虫期通常为9 d;6龄若虫发育历期最长,为10~11 d。雄性和雌性若虫为单独饲养,均能全部羽化为成虫。
2.4 主要生活习性
2.4.1求偶和交配
日本蚱的若虫羽化为成虫,7 d后出现交配的现象,交配时间通常为1~1.5 h(图4)。日本蚱交配前,有求偶行为:雄性会摆动触角,慢慢靠近雌性,在雌性保持相对静止时,爬到其背上;若雌性拒绝交配,则通过后足踢开雄性,或者直接跳离原地;若雌性接受雄性,则进行交配。在交配时,雄性身体会紧紧依附于雌性张开后足的一侧,保持静止状态;交配期间,雄性生殖器会膨大,弯曲腹部形成“J”形,使外生殖器插入到雌性的生殖器中,待交配完成后,雄性往往会出现轻微的抖动,再与雌性分开;交配过程中,雌性仍然有进食行为。日本蚱通常在白天交配,尤其是在中午时段,交配较为频繁,也有少数蚱在夜晚进行交配。雌性可以交配两次以上,极少出现交配一次后便死亡的现象;雄性通常在交配第二次后,一个星期内死亡。蚱在临死前,表现为行动迟缓,左侧与右侧后足不停颤抖,静息一段时间后仍然颤抖,直到最后死亡。
2.4.2产卵
日本蚱在交配成功后,雌性腹部会逐渐膨大,行动较之前也会变迟缓。日本蚱交配完成后,通常在1~3 d内会选择合适的地方产卵,卵一般产在泥土中或苔藓下,深度约为其腹部的长度。产卵时,雌性先是前后左右弯曲拱土形成凹槽,将腹部弯曲,扎入泥土中进行产卵。一般而言,产卵的数量与产卵的时间成正比。雌性产卵完成后,腹部从泥土中拔出,用后足拨土填平产卵形成的凹槽,以达到掩埋卵块的目的,而后静息约10 min,再跳离产卵的位置。若产卵时,受到人为的多次干扰,雌性会暂时选择放弃产卵,待确认无危险后,才会继续选择新地方产卵。雌性可以产卵两次或三次,且第一次产的卵块的卵粒数目较多,第二次产的卵粒数较少,有极少数雌性可以产三次卵。
2.4.3孵化
日本蚱通常在白天孵化,尤其是在中午时段,孵化较为集中,也有少数在早晨和夜晚孵化。孵化时,若虫从卵粒稍尖的一端,破卵壳爬出:破卵壳时,头先从卵壳伸出来,然后两前足伸出,协助虫体爬出,当虫体爬出卵壳约3/4时,若前足附近无苔藓等抓附物时,则若虫会通过左右扭动身体,使其全部爬出卵壳;若虫爬出卵壳的速度较快,整个过程历时约2 min。刚孵化出来时,虫体颜色是淡黄色,且都有一小块半透明膜状物穿在两尾须上,若虫会不断扭动腹部并用后足将其踢掉;在约30 min后颜色开始不断变暗,变为暗黄色,约1 h后变为褐色;孵化后的1龄若虫24 h后便可以取食苔藓。
2.4.4蜕皮及羽化
刚孵化的若虫为1龄,而后每增长一个虫龄都需要经过一次蜕皮。1龄若虫的腹部相对较长,其余各龄若虫,当腹部明显变长且超过后足股节末端时,表明即将进入蜕皮时期;若虫蜕皮时,头朝下方,六足紧抓苔藓等抓附物,腹部先收缩使新皮与旧皮分离;再由末端以波浪式的行进方式推向前胸背板,并不停的左右扭动身体,用力挤裂背板,接着头顶的旧皮开裂,虫体从背板裂口处爬出。低龄若虫蜕皮通常需要5~10 min,而后再延伸其背板;随着虫龄的增长蜕皮时间也会稍微延长。每次蜕皮后体色均为淡黄色,约1 h后颜色变深,成为正常体色。若虫最后一次蜕皮(羽化)多发生在下午6点至晚上10点左右,需要约10 min;蜕皮后,伸展折叠的后翅通常需要约12 min,延伸背板则需要约20 min,伸展后翅与延伸背板是同时进行的。刚羽化的成虫体质柔软,六足和前胸背板侧片后角均为浅白色,约20 min后颜色加深,约1 h后为正常体色;通常在约11 h后可以进食,48 h后背板基本变为正常硬度,即完成羽化过程,到达成虫阶段。
2.4.5前胸背板与翅型的多态性
在春、夏、秋、冬四季中,每月定点到广西河池市宜州区的日本蚱栖息地进行野外观察,并观察室内建立的实验种群,发现日本蚱雄性与雌性的前胸背板与翅型均存在多态现象。日本蚱的1龄若虫,其前胸背板上无明显的斑纹,体色也几乎一致;当若虫蜕皮至2龄时,少部分的若虫会出现多样化的斑纹;当若虫蜕皮至3龄时,斑纹会变得更加明显;在之后的蜕皮中,斑纹不再发生变化,但体色会有一定的差异(图5)。此外,日本蚱在1龄至羽化前期的若虫,其前胸背板与翅型是同步发育的,均为短背短翅型(即是:前胸背板不超过后足股节顶端,同时后翅不超过前胸背板顶端的个体);当若虫最后一次蜕皮(羽化)后,伸展折叠的后翅,同时延伸背板,到达成虫阶段。在这个时期,有时会同时出现短背短翅型(图6-A)、长背长翅型(即是:前胸背板明显超过后足股节顶端,同时后翅明显超过前胸背板顶端的个体)(图6-B)的日本蚱。此外,在长期的野外的观察中,也发现同一栖息地的日本蚱,其前胸背板与翅型大多数是相同的,但也有少数特殊情况,即同时存在多态性的背板斑纹、短背短翅型、长背长翅型的日本蚱。

图5 日本蚱前胸背板斑纹的多态性Fig.5 Polymorphism of Tetrix japonica in the pronotal dapple注:A~D,无斑纹;E~H,具两个对称的斑纹;I~L,具四个对称的斑纹;M~P,具四个对称的斑纹,且背板边缘具白边。标尺=1 mm。Note: A~D, No dapple; E~H, Two symmetrical dapple; I~L, Four symmetrical dapple; M~P, Four symmetrical dapple, and the edge of the pronotum in white. Scale bars=1 mm.

图6 日本蚱前胸背板与翅型的多态性Fig.6 Polymorphism of Tetrix japonica in the pronotum and wing morphs注:A,短背短翅型;B,长背长翅型。标尺=1 mm。Note: A, Short morph of pronotum and wing; B, Long morph of pronotum and wing. Scale bars=1 mm.
3 结论与讨论
本研究通过野外观察与室内饲养相结合的方法,对日本蚱开展了研究,详细描述了其卵块和各虫龄的形态特征、生活史,以及主要的生活习性,如求偶和交配、产卵、孵化、蜕皮及羽化、前胸背板与翅型的多态性等。日本蚱在羽化7 d后开始出现交配的行为,卵块通常产在苔藓下或松软的泥土中,卵块的整体形状呈橘瓣状,最外侧卵粒弯向卵块的中心,中心的卵粒直径小,长度较长;卵粒通常经过6~10 d孵化为若虫;依据蜕皮次数,雄性共有6龄,雌性共有7龄。各龄若虫的外部形态也具有一定的差异,1龄若虫触角的鞭节为8节,前胸背板不具明显斑纹,而后每增长一个龄期,鞭节数也会相应增加一节,前胸背板的斑纹也随虫龄的增长而更为明显;若虫最后一次蜕皮(羽化)为成虫后,后翅的翅脉具有横脉,后足股节具有膝前齿等。
此外,通过野外观察及室内饲养,还发现了日本蚱雄性与雌性的前胸背板与翅型均具有多态现象。前胸背板斑纹的多态性:日本蚱的 1龄若虫,其前胸背板上无明显的斑纹,体色也几乎一致;当若虫蜕皮至2龄时,少部分的若虫会出现多样化的斑纹;当若虫蜕皮至3龄时,斑纹会变得更加明显;在3龄之后的蜕皮中,斑纹不再发生变化,但体色会有一定的差异。早期也有学者曾对不同栖息环境的日本蚱的前胸背板斑纹开展研究,推测斑纹可能是日本蚱适应不同环境而出现的一种伪装现象(Tsuruietal., 2010)。本研究结果与Tsurui等的研究结果相似,推测日本蚱前胸背板斑纹具有多态性,主要作用是使其总有一定比例的体色会隐匿在与其背景相匹配的环境中,从而不易被捕食者察觉,这或许更有利于物种的生存和延续。
前胸背板与翅型的多态性:日本蚱在1龄至羽化前期的若虫,其前胸背板与翅型同步发育,均为短背短翅型;而当若虫最后一次蜕皮(羽化)后,伸展折叠的后翅,同时延伸背板,到达成虫阶段。在这个时期,则出现了短背短翅型和长背长翅型的日本蚱,即存在多型现象。该结果与直翅目的一些类群相似,如蝗虫(Riffatetal., 2009;Rosettietal., 2018),也存在翅型多态性现象,即每个物种有两个型:短翅型和长翅型。短翅型主要把能量用于繁殖以扩大种群;长翅型主要把能量投资用来迁移扩散。早期也有学者结合形态学和生物学特征,对部分蚱类昆虫的前胸背板与翅型的多态性进行探讨,如分类鉴定(Deng, 2021;Dingetal., 2021)、多态性的发生与季节及其栖息环境关系等(Pushkar, 2009;Lehmannetal., 2018)。本研究结果与前人的研究结果相似,推测日本蚱前胸背板与翅型的多态性,是其对应于栖息环境的时间和空间变化适应的结果,是应对复杂和不确定环境的进化而来的权衡策略。
昆虫的多态性在自然界普遍存在,通常被认为是一种进化适应结果,如昆虫体色的多态性,通常与其防御捕食者和抵御不良环境有关,其适应功能主要表现在隐匿与警戒色等方面(Guerra-Grenier, 2019)。在自然界中,有些昆虫体表的颜色与其周围的背景颜色高度匹配,从而隐藏在环境中不易被天敌发现。昆虫的前胸背板与翅型也存在多态性现象,常见的有短背短翅型和长背长翅型。然而,导致昆虫发育出不同前胸背板与翅型的因素十分复杂,通常认为环境因素和遗传因素均对多型的发育产生影响(戴华国等,1997)。综上所述,日本蚱的前胸背板与翅型的多态性是否同时具有以上多种适应功能需进一步研究。
