KlacPNP基因密码子优化及在枯草芽胞杆菌中的高效表达
2023-10-18赵越王新秀吴思陈作慧张会惠觅宙李杰双宝
赵越,王新秀,吴思,陈作慧,张会,惠觅宙,李杰,双宝
(东北农业大学 生命科学学院,黑龙江 哈尔滨,150006)
嘌呤核苷磷酸化酶(EC 2.4.2.1,purine nucleoside phosphorylase,PNP)是嘌呤补救合成途径中的关键酶,可逆的催化嘌呤核苷的磷酸化反应,生成嘌呤碱基和核糖-1-磷酸[1-2]。PNP属于N-水解和转移酶家族,普遍存在于细菌、真菌和哺乳动物中,根据PNP的寡聚状态、分子质量和底物特异性将其分成两类[3],第一类为低分子质量同源三聚体,特异性的以6-氧代嘌呤为底物,分子质量约为32 kDa,广泛存在于真核生物中,如人红细胞、牛脾脏细胞、酿酒酵母等[4-6];第二类为高分子质量同源六聚体,具有广泛的底物特异性,能够催化6-氧代嘌呤和6-氨基嘌呤等多种类型的核苷酸分解,分子质量约为26 kDa,普遍存在于细菌中,如大肠杆菌、蜡样芽胞杆菌、耐盐芽胞杆菌、嗜热栖热菌等[3,7-8]。研究发现,PNP在很多方面都起到重要作用,可作为前体物质合成利巴韦林,用作病毒性肺炎和支气管炎等病症的治疗[9],还广泛应用于低嘌呤食品的开发中。2016年,MAHOR等[10]将乳酸克鲁维酵母来源的PNP在大肠杆菌中成功重组表达,并对重组菌降低啤酒嘌呤方面的应用进行了初步的探索。在国内,李玉淼等[11]将大肠杆菌PNP基因在大肠杆菌BL21(DE3)中进行表达,酶活力较对照菌提高16倍,将酶添加到糖化醪液中,提高了嘌呤物质的利用率,能够降低发酵液中29%的游离嘌呤。
枯草芽胞杆菌(Bacillussubtilis)同大肠杆菌、毕赤酵母一样,是常见的工程菌株之一,普遍应用于外源蛋白的表达。相较于其他系统,枯草芽胞杆菌表达系统有一套高效分泌蛋白的启动子和信号肽,在蛋白表达过程中不易形成包涵体,能够稳定表达外源蛋白,易于分离纯化目的蛋白[12-13]。枯草芽胞杆菌表达存在一定的密码子偏好性,当外源基因含有非枯草芽胞杆菌偏好密码子时,将阻碍外源基因的表达。因此,根据表达宿主选择目的基因最优密码子,能有效提高目的蛋白异源表达效率,如陈灵艳等[14]通过密码子优化增加了目的基因在枯草芽胞杆菌中的表达量;KUMAR等[15]将密码子优化后的人干扰素基因转入枯草芽胞杆菌中表达,产物生成量比优化前提高30%。
本研究以NCBI中登录号CP042457.1的乳酸克鲁维酵母(Kluyveromyceslactis,Klac)PNP基因序列为基础,在不改变重组蛋白氨基酸序列的前提下,根据枯草芽胞杆菌的密码子偏好性对KlacPNP基因进行密码子优化。连接至枯草芽胞杆菌分泌表达载体,转入枯草芽胞杆菌WB600中,得到了高产PNP的重组菌株。通过摇瓶发酵及发酵条件单因素和响应面分析,初步优化了其发酵工艺。实现PNP在枯草芽胞杆菌表达系统中异源表达的同时,避免外源基因表达产生的弊端,提高PNP的表达水平,为PNP在枯草芽胞杆菌中生产奠定了基础,以期实现重组PNP的工业化生产,从而拓宽其在降低食品嘌呤中的广泛应用。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒
枯草芽胞杆菌(Bacillussubtills)WB600、表达载体pBSA43为本实验室保藏,大肠杆菌DH5α感受态购自TaKaRa生物工程有限公司。
1.1.2 主要试剂、仪器
PCR引物由睿博兴科公司合成;胶回收试剂盒、PCR产物纯化回收试剂盒、质粒提取试剂盒,OMEGA公司;限制性内切酶、T4 DNA连接酶,NEB生物科技有限公司;其他化学试剂均为国产分析纯。
PCR仪、电泳仪、凝胶成像系统,Bio-Rad公司;高效液相色谱仪,美国安捷伦公司。
1.1.3 培养基
LB(Luria-Bertani)液体培养基、LB固体培养基按文献[16]的方法配制。
发酵产酶培养基(g/L):蛋白胨 9.5, 葡萄糖12,K2HPO48.4,KH2PO42.8,(NH4)2SO41,CaCl20.2,MgSO40.5。
以上培养基均121 ℃灭菌20 min。
1.2 实验方法
1.2.1 PNP基因的合成及基因优化
根据KlacPNP的基因序列(GenBank登录号:CP042457.1)使用软件Signal P 3.0预测序列信号肽;在基因的5′端和3′端分别引入Hind III和BamH I限制性酶切位点,交由苏州金唯智生物科技有限公司进行基因合成(PNP-YP)和密码子优化(PNP-YO),并设计鉴定引物(表1)。
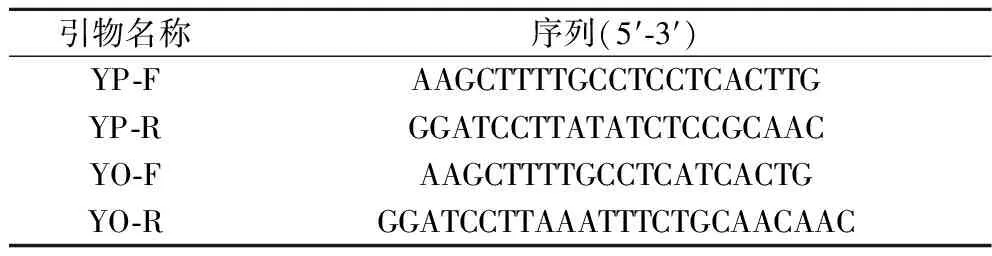
表1 基因扩增引物Table 1 Gene amplification primers
1.2.2 重组质粒的构建
合成的基因和载体pBSA43分别使用Hind III和BamH I进行双酶切,采用琼脂糖凝胶回收试剂盒回收目的片段。回收的目的片段按质量体积比1∶3(g∶mL)的用T4 DNA连接酶进行连接。连接产物转入到大肠杆菌DH5α感受态,加入LB培养基孵育1 h后取100 μL菌液,涂布于含100 μg/mL氨苄青霉素的LB平板上培养,12 h后挑取重组转化子,筛选鉴定,测序验证。
1.2.3 重组质粒在枯草芽胞杆菌中的表达
重组质粒在枯草芽胞杆菌WB600中的转化方法参照文献[17]。用表1中的引物进行质粒PCR鉴定,Hind Ⅲ和BamH I进行双酶切验证,获得枯草芽胞杆菌重组菌株pBSA43-YP-WB600、pBSA43-YO-WB600。
1.2.4 重组蛋白的表达和SDS-PAGE检测
分别挑取重组菌株pBSA43-YP-WB600、pBSA43-YO-WB600单菌落,接种于5 mL LB液体培养基中,培养至OD600值为0.6~0.8,按照10%接种量接至发酵产酶培养基,培养60 h后4 ℃、8 000 r/min离心10 min收集上清液,检测PNP的活力,同时进行十二烷基硫酸钠-聚丙烯酰氨凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis, SDS-PAGE)检测。所有试验均设置3个重复。
1.2.5 PNP活力的测定
取发酵液1 mL,8 000 r/min离心10 min,上清液即为粗酶液,酶活力参照文献[18]的方法测定。
重组PNP活力定义为:在上述反应条件下,每分钟产生1 μmol次黄嘌呤所需的酶量定义为1个酶活力单位(U/mL)。
1.2.6 PNP表达单因素试验
发酵温度37 ℃、培养基初始pH为7.0、发酵时间60 h作为初始发酵条件,分别探究发酵温度(27、32、37、42、47 ℃)、发酵时间(24、36、48、60、72 h)、初始pH值(5.0、6.0、7.0、8.0、9.0)对菌株产PNP活力的影响。
1.2.7 Box-Behnken 响应面试验设计与分析
在单因素试验的基础上,对重组菌产酶能力采用3因素3水平N=17的Box-Behnken响应面实验设计和研究,以发酵温度、培养基初始pH、发酵时间为响应面的自变量设计因素水平表(表2)。

表2 响应面因素水平表Table 2 Response surface factors and levels
2 结果与分析
2.1 PNP基因的密码子优化及人工合成
密码子优化方式参照文献[19],在保持氨基酸序列不变的基础上,合成优化的基因序列中共替换了227个碱基,优化后的密码子序列全部是枯草芽胞杆菌的偏爱密码子,为目的基因在枯草芽胞杆菌中成功表达提供了保证。基因序列如图1所示。

图1 基因优化前后PNP基因的核苷酸序列Fig.1 Alignment of nucleotide of the optimized and original PNP genes注:阴影部分为相同的核苷酸。
2.2 重组质粒的构建
将合成的包含目的基因的载体和pBSA43分别用Hind III和BamH I双酶切,并利用琼脂糖凝胶回收试剂盒回收目的片段和表达载体片段,然后用T4 DNA连接酶将目的片段连接到表达载体上,并转化到大肠杆菌DH5α中,提取重组质粒PNP-YP-pBSA43、PNP-YO-pBSA43,经质粒PCR(图2),Hind Ⅲ和BamH I双酶切(图3)鉴定,说明目的基因PNP-YP、PNP-YO成功连接到表达载体pBSA43上。
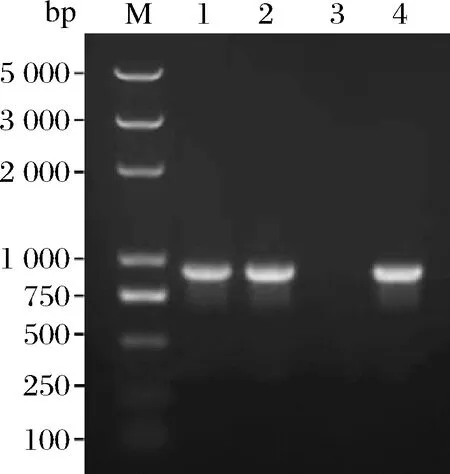
M-DNA 分子质量标准;1-PNP-YP-pBSA43 PCR 扩增产物;2-PNP-YO-pBSA43 PCR扩增产物;3-阴性对照;4-合成基因扩增产物(阳性对照)
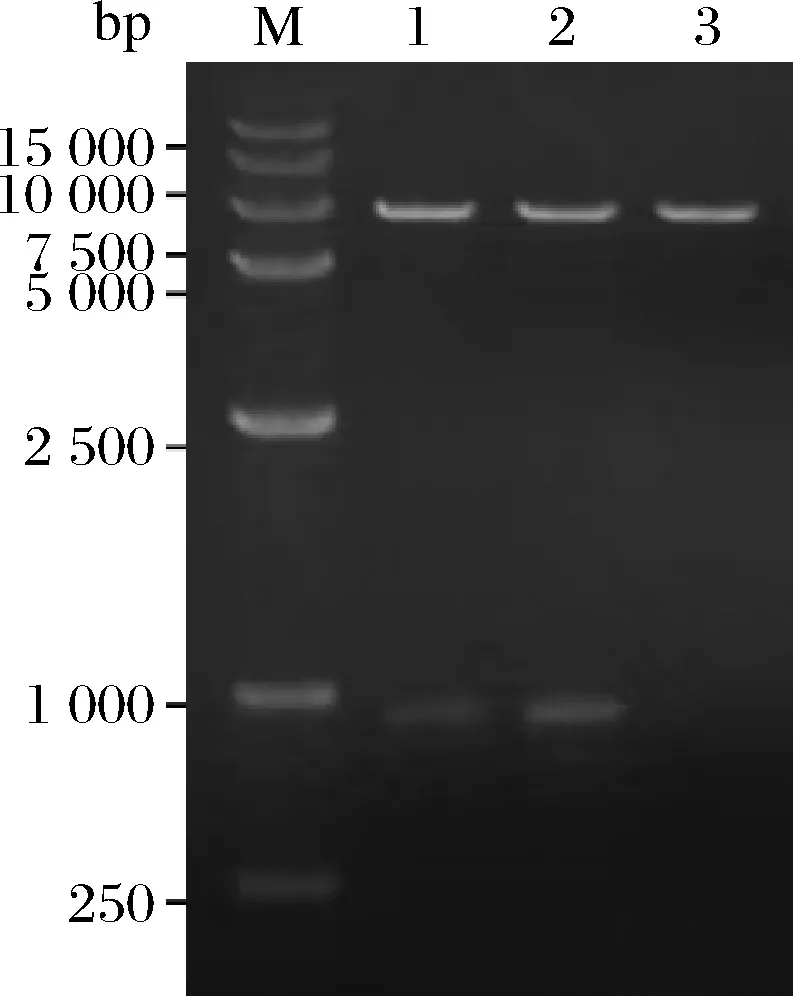
M-DNA分子质量标准;1-PNP-YP-pBSA43/Hind Ⅲ+BamH Ⅰ;2-PNP-YO-pBSA43/Hind Ⅲ+BamH Ⅰ;3-pBSA43/Hind Ⅲ+BamH Ⅰ(阴性对照)
2.3 重组表达菌株的构建
鉴定正确获得的PNP-YP-pBSA43、PNP-YO-pBSA43经化学转化法转化入枯草芽胞杆菌WB600中,通过卡那霉素抗性筛选,挑取单菌落鉴定后送睿博兴科公司测序,序列正确,说明重组菌株PNP-YP-pBSA43-WB600、PNP-YO-pBSA43-WB600构建成功。
2.4 重组菌株的表达目的蛋白
2.4.1 重组PNP-YP-pBSA43-WB600、PNP-YO-pBSA43-WB600的表达
收集重组菌株PNP-YP-pBSA43-WB600和PNP-YO-pBSA43-WB600发酵60 h的上清液,以原始菌株pBSA43-WB600作为对照,进行SDS-PAGE检测,如图4所示,PNP相对分子质量约为32 kDa(泳道2、3),而对照菌株未见相应条带(泳道1)。其中,重组菌株PNP-YO-pBSA43-WB600条带比PNP-YP-pBSA43-WB600条带更粗。表明PNP-YP-pBSA43-WB600、PNP-YO-pBSA43-WB600成功在枯草芽胞杆菌中分泌表达,且密码子优化后的重组菌株PNP-YO-pBSA43-WB600比PNP-YP-pBSA43-WB600表达量更高。

M-蛋白分子质量标准;1-pBSA43-WB600发酵液上清液(阴性对照);2-PNP-YP-pBSA43- WB600发酵液上清液;3-PNP-YO-pBSA43- WB600发酵液上清液
2.4.2 重组菌株最佳发酵时间的确定
微生物发酵过程中,利用培养基中的营养成分满足自身的生长,进而分泌蛋白,发酵时间的延长有利于胞外产物的累积。不同发酵时间的重组菌株PNP-YP-pBSA43-WB600和PNP-YO-pBSA43-WB600的PNP活力如图5所示,在24~60 h,随着发酵时间的延长,重组PNP活力逐渐增加,且密码子优化后的PNP-YO-pBSA43-WB600酶活力明显高于PNP-YP-pBSA43-WB600;发酵时间为60 h时,酶活力达到最高,此时PNP-YP-pBSA43-WB600酶活力为333.69 U/mL,PNP-YO-pBSA43-WB600酶活力为351.61 U/mL。当超过60 h后,重组PNP活力均有所下降,密码子优化后的PNP-YO-pBSA43-WB600酶活力仍然高于PNP-YP-pBSA43-WB600。因此,选择将重组菌株发酵至60 h,以实现对重组PNP的高效表达。

图5 发酵时间对重组PNP活力的影响Fig.5 Effect of fermentation time on activity of recombinant PNP
2.4.3 发酵温度对重组菌株酶活力的影响
培养温度是微生物发酵中的重要条件,温度过高或过低都会影响菌体生长代谢和产酶能力。由图6可知,随着温度的升高,重组酶活力呈先升高后下降的趋势,且密码子优化后的PNP-YO-pBSA43-WB600酶活力始终高于PNP-YP-pBSA43-WB600,分析原因可能是温度低于32 ℃时,菌体生长减缓,产生酶蛋白含量少,总体酶活性不高。当发酵温度为32 ℃时,酶活力最高,PNP-YP-pBSA43-WB600为323.50 U/mL,PNP-YO-pBSA43-WB600为333.79 U/mL,由此确定重组菌株的最佳发酵温度为32 ℃。随着温度的升高,菌体生长旺盛,不利于产物的积累。当温度>37 ℃时,菌株的生长环境遭到破坏,其产酶能力下降。

图6 发酵温度对重组PNP活力的影响Fig.6 Effect of fermentation temperature on the activity of recombinant PNP
2.4.4 初始pH对重组菌株产酶活力的影响
pH是影响细胞膜通透性和表面电荷性质的重要因素,影响着微生物对于营养物质的摄取及代谢产物的分泌,从而影响菌株生长状态和产酶能力。由图7可知,当初始pH<7时,随着pH值的升高重组菌株酶活力呈上升趋势;当初始pH=7时,酶活力达到最大值,PNP-YP-pBSA43-WB600酶活力为333.02 U/mL,PNP-YO-pBSA43-WB600为341.27 U/mL;当初始pH>7时,随着pH值的升高酶活力呈递减趋势,说明培养基初始pH升高或降低都会降低重组菌株的产酶能力,但密码子优化后的PNP-YO-pBSA43-WB600酶活力明显高于PNP-YP-pBSA43-WB600。

图7 培养基初始pH重组PNP活力的影响Fig.7 Effect of initial pH of the medium on the activity of recombinant PNP
2.5 重组菌响应面实验优化
在单因素的基础上,设计了17个试验的响应分析试验,以培养基初始pH值(A)、发酵时间(B)、发酵温度(C)3个因素为自变量,发酵液酶活力为因变量,响应面实验设计与结果如表3所示。
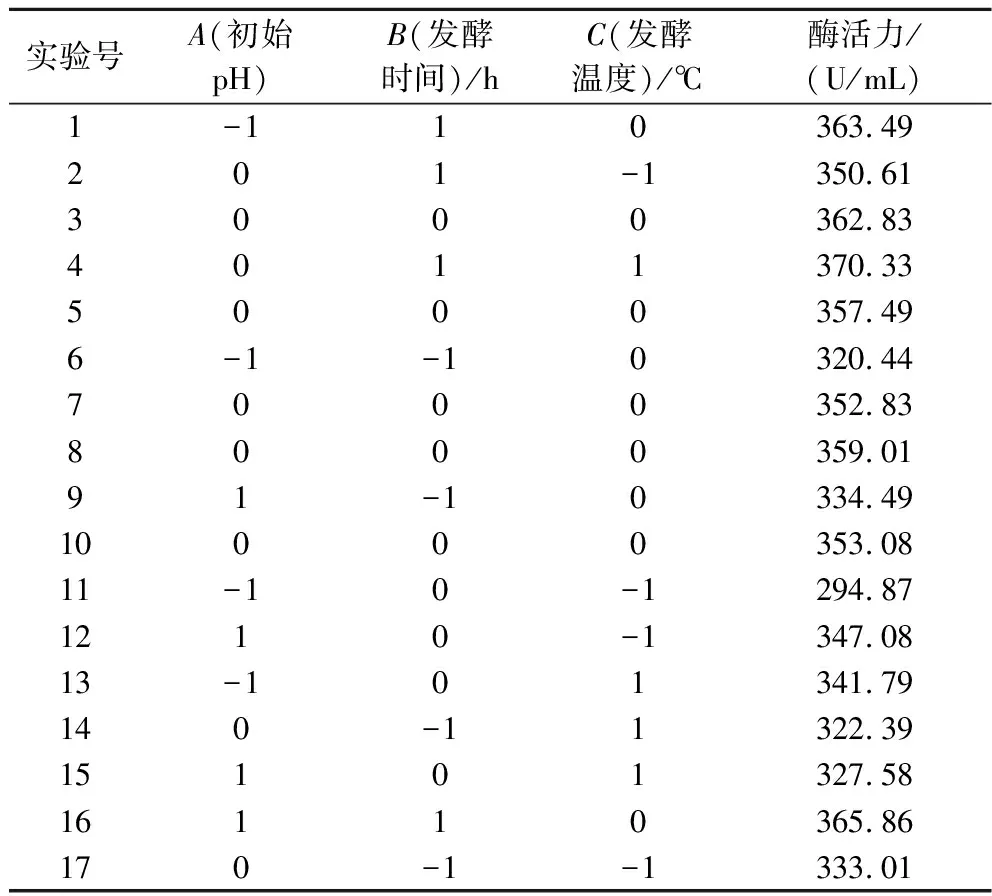
表3 响应面实验计划表Table 3 Response surface experiment schedule
利用Design Expert 13软件对响应面结果进行统计学分析,通过方差分析(表4)可知,在误差允许范围内,3个因素的P>F值均小于0.01,表明该模型显著且具有统计学意义。通过最小二乘法回归方程得到的重组菌PNP-YO-pBSA43-WB600产酶活力对于发酵温度、发酵初始pH和发酵时间的二次多项回归模型:酶活力(Y)=357.05+6.80A+17.49B+4.57C-2.92AB-16.6AC+7.58BC-13.62A2+2.64B2-15.60C2。当变量为1时,线性拟合系数R2=0.935 8,说明回归方程拟合度良好。

表4 回归模型方差分析结果Table 4 Regression model ANOVA results
响应面优化分析结果如图8所示,3个因素在不同水平对酶活力产生不同的影响,最大值处于三维模型的中心。通过回归方程求得3个因素最佳发酵条件为初始pH=6.67、发酵时间55 h、发酵温度28.7 ℃,理论预测酶活力为333.97 U/mL,在优化后的条件下摇瓶发酵酶活力达到372.79 U/mL,为预测值的111.62%。

a-培养基初始pH值和发酵时间;b-培养基初始pH值和发酵温度;c-发酵时间和发酵温度
3 结论与讨论
本研究以NCBI上登录号为CP042457.1的乳酸克鲁维酵母的PNP基因为基础,根据枯草芽胞杆菌的密码子偏好性,对KlacPNP进行密码子优化,在不改变蛋白质序列、增强目的基因在枯草芽胞杆菌中的适应性的情况下高效表达,得到了高产PNP的重组菌株。密码子优化前摇瓶发酵的酶活力为333.69 U/mL,优化后摇瓶发酵的酶活力达到351.61 U/mL。基于单因素和响应面优化确定了最佳发酵条件为:初始pH 6.67、发酵时间55 h、发酵温度28.7 ℃。最后在优化后的条件下进行发酵试验,摇瓶发酵酶活力达到372.79 U/mL,为预测值的111.62%。优化后的产量是优化前的1.12倍,得到了理想的优化结果。
前期报道中,MAHOR等[10]将klacPNP转入大肠杆菌成功表达,证明了klacPNP能够有效降低啤酒中的嘌呤含量,在klacPNP的基础上,通过点突变构建了对6-氧代嘌呤和6-氨基嘌呤具有较强底物特异性的突变体klacPNP 256D和对肌苷具有高度特异性的klacPNP 256E,二者对于啤酒中嘌呤含量都有较好的效果。TIMOFEEV等[20]将嗜热菌HB27的PNP在大肠杆菌中表达,TthPNP I高度作用于肌苷底物,每分钟降解肌苷能力330 μmol/mg,TthPNP II高度作用于腺苷底物,酶活力达到830 μmol/mg。嗜极菌嗜色还原卤单胞菌AGD 8-3[21]转化入大肠杆菌,酶活力达到28.6 U/mg。已报道人源PNP的酶活力为80 U/mg[22]。但以上重组酶表达宿主均为大肠杆菌,本试验利用枯草芽胞杆菌作为宿主细胞,通过密码子优化提高表达量,其表达乳酸克鲁维酵母来源的PNP活性有一定的提高,且对于PNP的安全生产具有重要意义。
