冬小麦叶片和冠层尺度的光合-蒸腾模拟
2023-10-10张传伟沈彦俊张玉翠
赵 阳,张传伟,沈彦俊,张玉翠
(1.中国科学院遗传与发育生物学研究所农业资源研究中心/中国科学院农业水资源重点实验室/河北省节水农业重点实验室,河北 石家庄 050022;2.中国科学院大学现代农业科学学院,北京 100049)
植物光合是生态系统有机物的重要来源,蒸腾作用是与之相伴的水分散失过程,两者之间的耦合模拟是评价全球和区域初级生产力、模拟作物生长、研究陆面过程与气候相互作用和预测生态环境变化的重要手段,对研究地球表层能量物质迁移、地球生物化学循环等有重要意义[1]。
植物光合和蒸腾作用耦合的常用方法是把气孔导度和蒸腾作用耦合到一个光合作用模型中[2-4]。气孔作为控制植物和外界进行水分与气体交换的通道,通过影响CO2和水汽的扩散对光合作用和蒸腾作用实现控制。因此,气孔导度是水碳耦合模型的关键变量。气孔导度受光合有效辐射、相对湿度、土壤含水量、温度、CO2浓度5种环境因子的影响,并且随着外界条件的改变而改变,相比将其设为一个常数代入模型中应用,引入气孔导度模型不仅可以提高模拟精度[5],而且对预测植物应对气候条件的改变更具有价值。目前,气孔导度模型的形式众多,可以广义地划分为3种类型——乘和或者多重因素限制模型(Jarvis,1976[6]);基于气孔导度和光合速率线性关系的半经验模型(Ball-Berry类[7-8])以及基于脱落酸(ABA)信号、叶片水势或水力控制的水分胁迫响应模型[9]。其中由于Ball-Berry类模型(以下简称BB模型)所需要的参数较少且易获取,被广泛用于叶片[2,10-11]、冠层[12-13]以及区域和全球系统模型(ESMs)中,用于模拟水和碳的循环过程。
在进行模型的耦合时,植物的功能类型、研究尺度及研究目的不同,模型耦合的侧重点就不同,所采用的模型表达形式也不同,这造成模型复杂程度的较大差异,对实际应用影响较大。Yu等[14]基于气孔控制光合与蒸腾的生理机制,引入了CO2内部导度的概念,开发了基于气孔行为的光合-蒸腾耦合模型(SMPT-SB模型),模型所需参数易获取,并且在玉米、大豆及树木的叶片尺度应用,获得了较好的结果。任传友等[15]将该模型进行尺度提升,在长白山温带阔叶林冠层尺度研究中获得了良好的适用性。以上研究均将光合速率和蒸腾速率独立进行研究,尚未涵盖水碳耦合方面的研究。张宝忠等[16]利用该模型计算了夏玉米叶片及冠层的光合、蒸腾及水分利用效率,与实测值一致性较好,在不同尺度研究水碳耦合关系。然而,利用该模型模拟冬小麦的光合和蒸腾特性还未有定论。此外,冠层尺度的水碳耦合是衡量农田蒸散和作物生产力以及农田水分利用状况的重要内容,同时也是区域或全球蒸散模型应用和验证的基础。
因此,本文的研究目的主要有:(1)探究冬小麦叶片和冠层尺度的光合及蒸腾作用日变化规律,验证SMPT-SB模型对于冬小麦的适用性;(2)借助该模型实现作物水分利用研究从叶片水平到冠层尺度的扩展,明确冬小麦叶片和冠层水碳交换的关系,为提高冬小麦多尺度水分利用效率提供科学依据。
1 材料与方法
1.1 研究区概况
试验在中国科学院栾城农业生态系统试验站(37°53′N,114°41′E,海拔50.1 m)进行,该站位于太行山山前平原,属于半干旱半湿润季风气候,种植模式以冬小麦-夏玉米一年两熟为主。年平均温度为11.5℃,1月月均温度最低(-5.6℃),7月月均温度最高(26.7℃);年均降雨量487 mm,70%的降水出现在7—9月[17]。2011—2015年均蒸散量690 mm[18],冬小麦品种为‘科农199’,种植期间通常进行2~3次灌溉。小麦季关键生育期的划分见表1。

表1 冬小麦生育时期的划分Table 1 Growth periods of winter wheat
1.2 田间观测试验
在冬小麦生长发育期间选取4个晴日进行叶片尺度气体交换数据的测量,9∶00—18∶00每小时观测1组数据,冠层尺度数据由涡动相关系统测得,从中选出相对应的4个晴日数据。
叶片气体交换参数的获取:利用LI-6400便携式光合作用系统(LICOR Inc.,Lincoln,NE,USA),测定小麦第一片完全展开叶片的净光合速率 (AL,μmol·m-2·s-1)、气孔导度(gs,mol·m-2·s-1)、蒸腾速率(TL,mmol·m-2·s-1)等叶片气体交换参数以及叶温(T1,℃)、相对湿度(hs)等环境因子。观测时采用透明叶室进行自然光观测,随机选取6个小区,每个小区随机选择3株小麦,测量时每株计数3次,最后求均值作为每个小区的代表值。
冠层尺度数据的获取:大田中建有通量观测塔,涡度相关系统的安装高度为3.5 m,由三维超声风速仪(CSAT3,Campbell,USA)和开路式CO2/H2O分析仪(Li-7500A,LICOR,USA)组成,采样频率为10 Hz,数据采集器为CR3000,同时输出30 min平均值,测量数据主要包括生态系统的水碳通量、三维风速等。光合有效辐射、温湿度、降雨量等由塔上安装的光合有效辐射表(LI-190SL,LICOR,USA)、温湿度计(HMP155,Vaisala,Finland)、雨量筒(TE525MM,Texas Electronics,USA)等观测获得。
叶面积指数和株高的观测:在叶片气体交换参数测量的同时进行叶面积和株高的测量,每个试验小区随机选择10株小麦,用直尺量取每株植株所有有效叶片的长和宽,并记录茎数和作物种植密度,最后计算得到整个田块的叶面积指数。
1.3 不同尺度光合-蒸腾耦合模型
1.3.1 叶片尺度水碳耦合模型 忽略水分子进出气孔对CO2吸收造成的影响,叶片碳吸收过程受到叶片气孔、边界层及内部生物化学及光化学过程的影响,叶片尺度的光合速率[16]可以表示为:
(1)
其中,gsc、gbc、gic分别为叶片气孔、边界层、叶片内部对CO2的导度(mol·m-2·s-1);Ca为环境CO2浓度(μmol·mol-1);Γ*为叶片无暗呼吸时的CO2补偿点(μmol·mol-1)。
叶片的蒸腾受到边界层阻力和气孔阻力的共同作用,因此,叶片尺度的蒸腾速率[16]可以表示为:
(2)
其中,Wi、Wa分别为叶片内外水汽浓度(mmol·mol-1);gsw、gbw分别为叶片气孔和边界层对水汽的导度(mol·m-2·s-1)。
叶片气孔导度利用Luening修正的Ball-Woodrow-Berry模型进行估算:
(3)
其中,m为参数;f(Ds)为水汽响应函数;Cs为叶片周围CO2浓度(μmol·mol-1);Γ为有暗呼吸时的CO2补偿点(μmol·mol-1)。g0表示当净光合速率趋向于0时的最小气孔导度(mol·m-2·s-1),当考虑气孔对CO2扩散的影响时可忽略;而Γ和Γ*在田间试验条件下相差很小,对模拟结果造成的误差可忽略,因此,假设Γ=Γ*,同时满足关系gsw=1.56gbw和gbw=1.37gbc,将公式(3)代入公式(1)、(2)中,即可得到叶片尺度的光合和蒸腾速率模型:
(4)
(5)
1.3.2 冠层尺度水碳耦合模型 与叶片尺度不同的是,冠层尺度考虑水碳传输阻力时,需要考虑到空气动力学阻力的影响。因此,冠层尺度的光合速率模型[16]为:
(6)
式中,AC为冠层光合速率(μmol·m-2·s-1);Ca-C为参考高度处冠层大气CO2浓度(μmol·mol-1);Γ*-C为冠层无暗呼吸时的CO2补偿点(μmol·mol-1);gbw-C为冠层对水汽的边界层导度(mol ·m-2·s-1);gic-C为冠层内部导度(mol·m-2·s-1);ga-C为空气动力学导度(mol·m-2·s-1)。
冠层尺度的蒸腾速率模型为:
(7)
式中,TC为冠层蒸腾速率(mmol·m-2·s-1);Wi-C、Wa-C为冠层内和冠层参考高度处的水汽浓度(mmol·mol-1);g0-C为冠层净光合速率趋向于0时的气孔导度(mol·m-2·s-1)。
1.4 模型参数和水汽响应函数f(Ds)的确定
1.4.1 叶片和冠层阻力参数的确定
(1)内部阻力的计算。净同化速率不仅取决于气体的扩散,同时也受到叶肉中生化和光化学过程的控制,如光反应、暗反应等,Jones等[19]提出叶片内部导度:
(8)
其中,Ci为胞间CO2浓度(μmol·mol-1)。
研究表明,对于C3植物而言,胞内CO2浓度与外界CO2浓度之间的比值通常保持为一定值,因此,冠层内部CO2浓度可近似采用下面的关系式求解[19]:
Ci=0.7Ca
(9)
那么冠层内部导度为:
(10)
而Γ*可用下式[20]计算:
Γ*=42.7+1.68(T-25)+0.012(T-25)2
(11)
(2)边界层阻力的计算。叶片尺度的边界层阻力(rbw,m2·s·mol-1)可由LI-6400直接测量得到,冠层尺度的边界层阻力可以由下式表示[21]:
(12)
式中,α为冠层内风速衰减系数(α=3);W为叶宽幅(cm),根据试验中测量的叶片宽度取1.5 cm;Uh为冠层高度h处的风速(m·s-1);rbw-C为冠层对水汽的边界层阻力(m2·s·mol-1);2.24×10-2为单位由mol·m-2·s-1向m2·s·mol-1转换的转换系数。不同时期的叶面积指数(LAI)见表2。
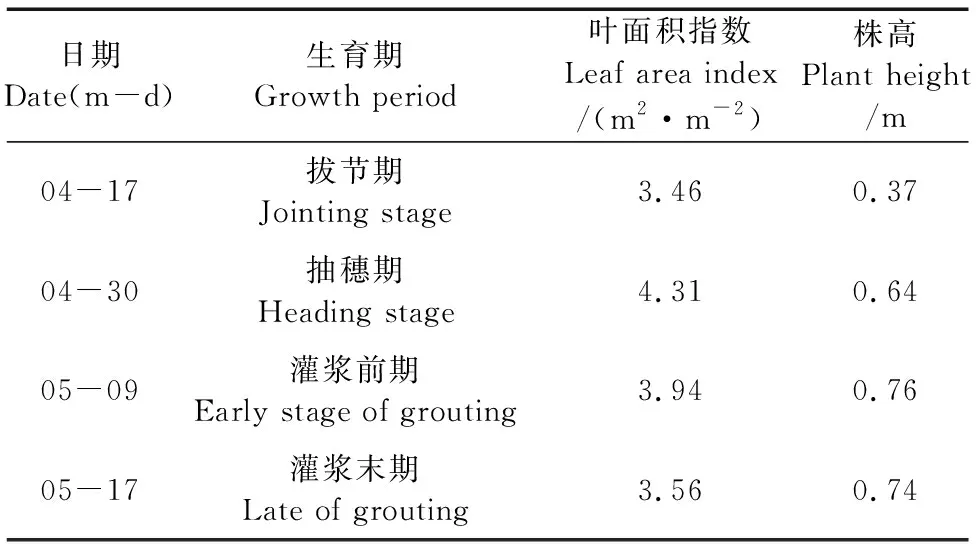
表2 不同生育期冬小麦叶面积指数及株高Table 2 Leaf area index and plant height of winter wheat at different growth periods
(3)空气动力学阻力的计算。空气动力学阻力由冠层内和冠层上方空气流动决定,冬小麦冠层在中性层结条件下,动力学阻力可由下式计算:
(13)
式中,UZ为参考高度处的风速(m·s-1);k为卡曼常数,其值取0.41;Z为观测湿度和风速的高度(m);d为零平面位移(m);Z0为表面粗糙长度(m)。
对于小麦而言,d和Z0的值可由下式进行估算[22]:
d=0.63h
(14)
Z0=0.13h
(15)
式中,h为植株的高度(m)。
1.4.2 水汽响应函数 Ball-Woodrow-Berry气孔导度模型中的参数m代表了气孔导度对光合速率、CO2浓度、相对湿度以及温度的综合敏感性,其值由于水分胁迫程度、物种以及作物功能类型的不同有所差异。气孔导度对水汽的响应函数具有多种形式,本研究中对比了常见的3种形式(图1),当f(Ds)=RH时,斜率m=9.05,这也与一些C3植物模型中的m=9.0[22-23]相符,气孔导度模拟值和实测值的决定系数(R2=0.75)也在3种形式中最高,因此,选择此函数作为水碳耦合时的水汽响应函数。
1.5 模型验证
将同时期的叶片和冠层尺度仪器观测的光合和蒸腾数据与模拟结果进行对比,建立模拟和观测值之间的相关性分析。通过计算模拟值与实测值的回归斜率b、决定系数R2、均方根误差RMSE对模型模拟效果进行评价。
2 结果与讨论
2.1 叶片光合及蒸腾速率日变化实测与模拟
在外界环境条件影响下,光合与蒸腾的日变化可以反映出冬小麦通过气孔开闭对光合与蒸腾的调节作用。从实测结果来看(图2),冬小麦的光合速率呈先增后减的趋势,通常在11∶00左右达到峰值。上午随着光合有效辐射的增加以及相对湿度的减小,植物叶片气孔导度增大,光合速率逐渐增大;而14:00之后光合有效辐射快速下降,相对湿度也逐渐增大,光合速率迅速下降。4月17日以及30日12∶00时光合和蒸腾速率快速下降,其原因是温度较高且辐射强,部分气孔关闭以减少水分的散失,存在明显的“光合午休”现象。由于5月9日14∶00—15∶00不时有云层遮蔽阳光,平均光合速率下降较快。实测值与模拟值的日变化趋势一致,模型完整地模拟出了小麦叶片光合在一天中的波动情况。实测值显示,从整个生育期来看,4月17日的日平均光合速率为17.94 μmol·m-2·s-1,4月30日光合速率日均值达到最大20.95 μmol·m-2·s-1,这两个时期内小麦生长速度较快,5月9日光合速率日均值相对抽穗期下降了20%左右,5月17日达到最低13.85 μmol·m-2·s-1。模拟和实测的日均值最大相对误差为0.05%,每小时模拟结果的相对误差在0.14~3.85 μmol·m-2·s-1之间。
冬小麦叶片在4个典型日的蒸腾速率日变化实测和模拟值如图3所示,与光合速率类似,小麦叶片的蒸腾速率也呈现先增后减的趋势。4个典型日中,由于4月17日当天晴朗无风,叶片周围水汽凝聚,因此当天蒸腾速率变化波动较小。4月17日及30日蒸腾速率相差不大,但从4月30日到5月9日,蒸腾速率明显增大,达到了8.05 mmol·m-2·s-1,主要因为小麦叶面积随着发育进程而增大,水分消耗速率加快。模型模拟的小麦蒸腾速率日变化趋势与实测结果一致,并且实测值与模拟值相差的绝对值较小,最大差值仅为0.41 mmol·m-2·s-1,分时段的模拟结果中也不超过2 mmol·m-2·s-1。因此,模型在误差控制和趋势模拟方面表现良好,可以较准确地反映出小麦在一天之内的水分消耗和CO2吸收情况。
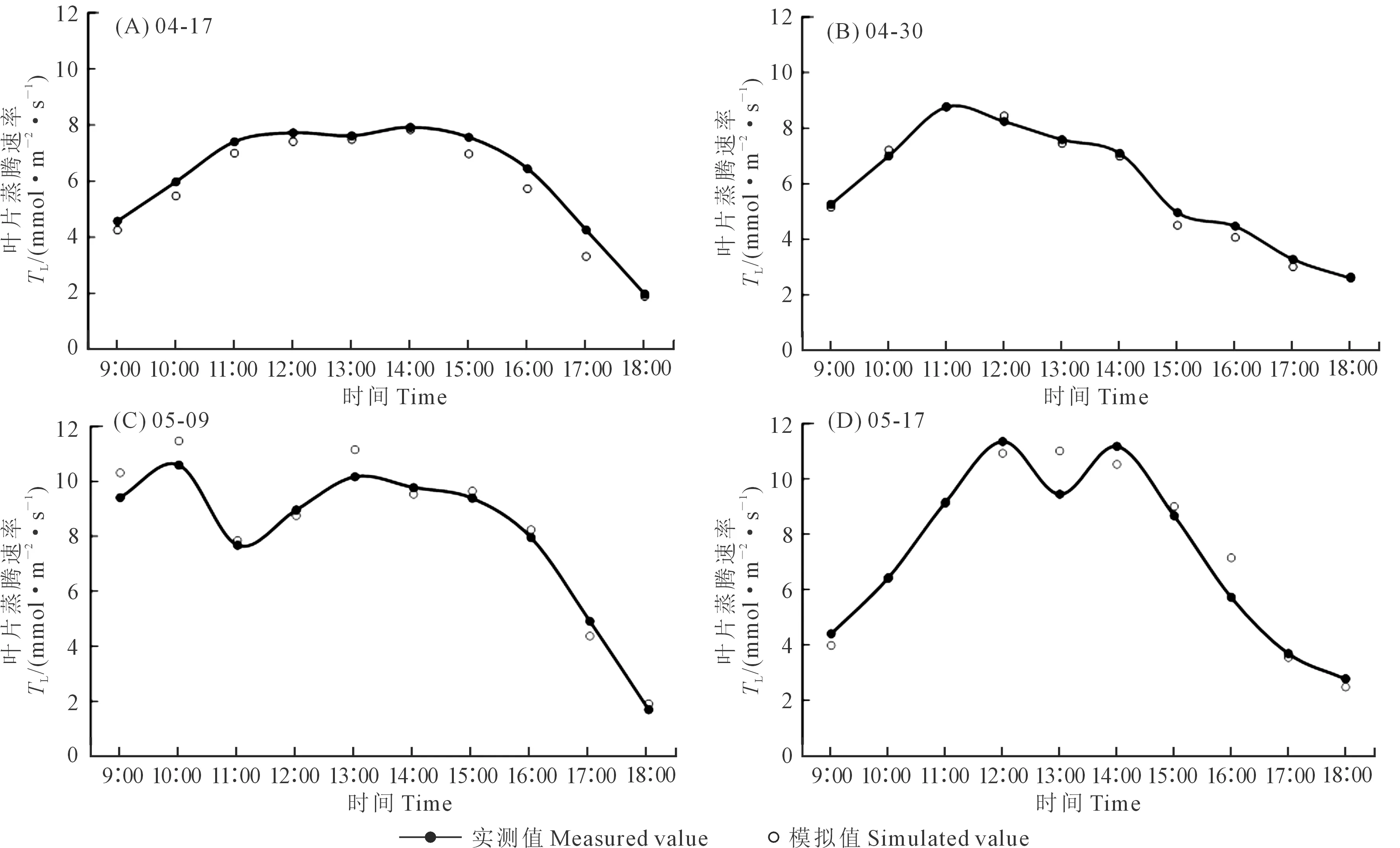
图3 叶片蒸腾速率日变化实测和模拟值对比Fig.3 Comparison of measured and simulated diurnal variation of leaf transpiration rate
2.2 冠层光合和蒸腾速率日变化实测与模拟
冠层尺度的光合和蒸腾速率变化更能反映农田整体情况。如图4冠层尺度光合速率实测和模拟值日变化对比所示,与叶片尺度相同的是,冠层尺度光合速率也呈现上午逐渐增加、14∶00以后快速下降、至18∶00下降至0附近的趋势。与叶片尺度的光合速率变化不同,冠层尺度光合速率日变化没有明显的“光合午休”现象,冠层尺度的光合速率最大日平均值出现在5月9日(23.81 μmol·m-2·s-1),因此可以推断,影响冠层光合速率变化的环境因子与叶片尺度有所不同,冠层光合速率并不是叶片光合速率的简单加和。从模型的模拟效果上来看,模拟值与实测值也有着相同的变化趋势,模型的误差也较小,最大误差仅为0.84 μmol·m-2·s-1。
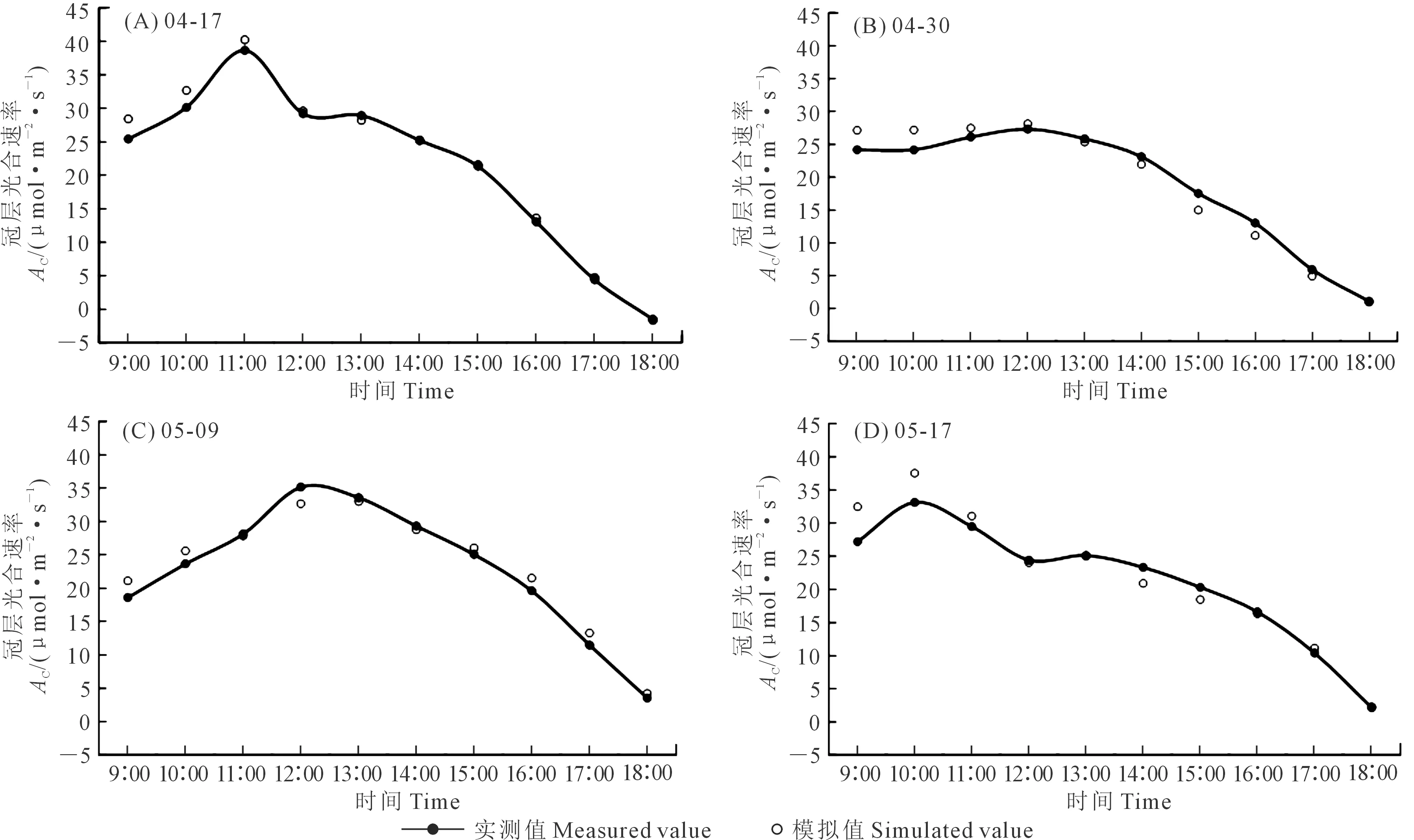
图4 冠层光合速率日变化实测与模拟值对比Fig.4 Comparison of measured and simulated diurnal variation of canopy photosynthetic rate
相比于冠层光合速率的模拟结果,冠层蒸腾速率效果略低,但仍然保持了与实测结果变化趋势的一致性(图5)。需要说明的是,模型中将冬小麦冠层视为“大叶模型”,将蒸腾视为唯一涌源,但涡动相关系统所观测的“蒸腾”速率实际为“蒸腾+蒸发”速率,虽然这种误差会随着小麦叶面积指数的增加逐渐减小,但仍然不可忽略。这也是冠层蒸腾速率模拟精度低于光合速率的原因之一。但从实际的模拟情况来看,模型仍然适用于冠层蒸腾速率的模拟,分时段模拟误差在-1.63~2.21 mmol·m-2·s-1之间,日均误差不超过0.15 mmol·m-2·s-1。
图5 冠层蒸腾速率日变化实测与模拟值对比Fig.5 Comparison of measured and simulated diurnal variation of canopy transpiration rate
2.3 不同尺度光合和蒸腾速率模拟效果评价
利用LI-6400便携式光合仪观测的叶片尺度和涡度相关系统观测的冠层尺度的实测数据,与模型模拟的数据进行相关性分析表明(图1,图6),不论是叶片还是冠层尺度,各散点均匀、紧密地分布在1∶1线的两侧,并且没有明显偏离的点。叶片和冠层尺度光合、蒸腾速率模拟的回归斜率在0.8~1.0之间,叶片和冠层光合速率模拟的RMSE分别为2.14 μmol·m-2·s-1和2.48 μmol·m-2·s-1(表3),而蒸腾速率模拟的RMSE分别为0.86 mmol·m-2·s-1

图6 叶片和冠层尺度光合及蒸腾速率模拟值及实测值对比Fig.6 Comparison of simulated and measured photosynthetic and transpiration rates at leaf and canopy scales

表3 叶片、冠层尺度光合和蒸腾速率模拟统计参数Table 3 Simulation statistical parameters of photosynthetic and transpiration rates at leaf and canopy scales
和0.30 mmol·m-2·s-1,R2除冠层蒸腾速率为0.88外,其他均在0.9以上,相关性显著。但从叶片和冠层两个尺度比较来看,叶片尺度的模拟效果要优于冠层尺度,这是因为将冠层假设为“一片叶子”,忽略了土壤蒸发,即使小麦的冠层高度小,但是仍然存在辐射和风速在冠层中衰减等因素,整个冠层并不是完全的均一状态;且叶片尺度向冠层尺度的提升,需要考虑辐射、湿度、温度、土壤水分、风速等各方面的综合影响,这是一个复杂的过程。即便如此,通过对模拟结果与实测值的对比分析,该模型的模拟效果仍然处于较高水平,至于上面所提到的缺点,需要尽可能提高仪器测量精度,同时也需要将更多的影响因素逐渐归纳至模型中。
2.4 不同m值确定方式对模拟结果的影响
气孔导度对水汽响应函数的斜率值m作为模型的重要参数,通常会对模拟蒸腾[24]和估算碳通量[25]产生显著影响,其值通常在物种间或者同一物种内都有较大差异,目前尚难以量化这种差异。因此,在明确“大叶模型”适用于冬小麦的前提下,模拟冠层光合和蒸腾速率时忽略了土壤蒸发以及冠层中不同部位叶片光合能力不同的影响,采取了不同时期拟合不同m值的方式,将其模拟结果与各个时期采用同一个m值的模拟结果进行了对比。如图7所示,在4月17日和5月17日,采用一个恒定的m=9.6模拟时,模拟效果仍然较好,其模拟的趋势与实测结果一致,且模拟值与实测值之间绝对误差小。这两天的相对湿度日均值(9∶00—18∶00)较大并且相差很小,4月17日为54%,5月17日为51%;而4月30日和5月9日两天的相对湿度较小分别为22%和31%,导致该因子对光合速率模拟的影响作用突出,其m值恒定为9.6时冠层光合速率AC-mm模型模拟值相对于实测值明显低估。虽然仍能反映出基本相同的变化趋势,但与实测值之间的绝对误差较大,不能反映真实光合情况。
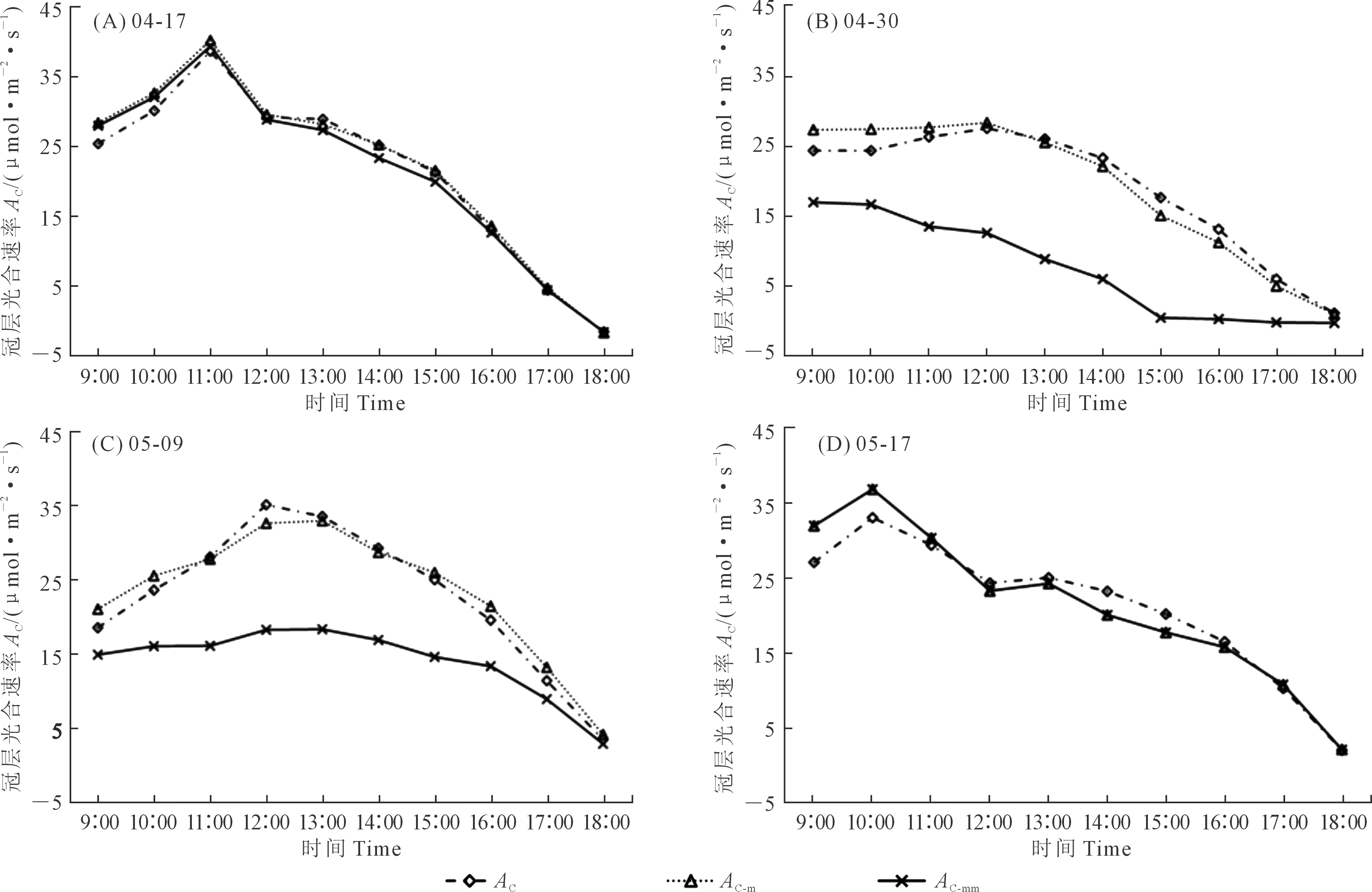
注:AC为实测值,AC-m和AC-mm分别为分时期拟合m值和恒定m值拟合结果。Note: AC is the measured value, AC-m and AC-mm are the fitting results of period-specific fitting m value and constant m value, respectively.图7 不同m值率定方式下光合速率日变化实测与模拟趋势对比Fig.7 Comparison of measured and simulated diurnal variation trends of photosynthetic rate under different m value calibration methods
m值的确定对于冠层尺度蒸腾速率的模拟效果也有影响(图8),当采用恒定的m值时,模型亦能大致反映出冠层尺度蒸腾速率的日变化情况,但其模拟值相对于实测值出现明显波动(5月17日),模拟效果并不理想。冠层尺度的光合和蒸腾速率如果用恒定的m值进行模拟,那么“m×RH”这个因子在相对湿度变化较大时,不能真实准确反映出其实际情况,从而降低模型的模拟精度。因此,冠层尺度的模拟分时期模拟参数m值会大大提高模型模拟精度。
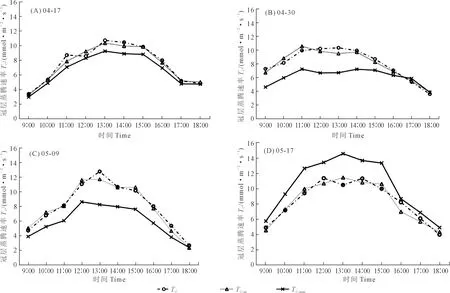
注:TC为实测值,TC-m和TC-mm分别为分时期拟合m值和恒定m值拟合结果。Note: TC is the measured value, TC-m and TC-mm are the fitting results of period-specific fitting m value and constant m value, respectively.图8 不同m值率定方式下蒸腾速率日变化实测与模拟趋势对比Fig.8 Comparison of measured and simulated diurnal variation trends of transpiration rate under different m value calibration methods
3 结 论
本研究基于SMPT-SB模型,将其与改进的Ball气孔导度模型耦合,建立了叶片和冠层尺度的水碳耦合模型。系统分析和模拟了冬小麦叶片和冠层尺度光合、蒸腾速率的日变化趋势,讨论了模型在两个尺度的适用性。冬小麦叶片和冠层光合、蒸腾速率存在“单峰”和“双峰”两种形式,这是当辐射大、温度高时作物自身气孔调节造成的。耦合在气孔导度模型中的参数m,在叶片尺度模拟时其值通常在8~10之间,但将其提升到冠层尺度后,在相对湿度差异较大的时期,其值存在较大误差,m值无法精确反映气孔导度的变化,从而降低模拟效果,而分时段率定m值则会大幅提高模型模拟精度。叶片尺度模拟结果优于冠层尺度。模型对于小麦在日尺度上的水分消耗和碳吸收变化模拟效果良好,模拟值与实测值具有相同的变化趋势,并且绝对误差控制合理,因此该模型适用于冬小麦叶片及冠层尺度的土壤-植物-大气之间水碳交换的模拟。
