3种环境因子对刚毛藻吸收氮的影响研究
2023-09-27王路平唐永涛唐伯平成永旭
王路平,唐永涛,汪 晨,唐伯平,成永旭
( 1.上海海洋大学 水产与生命学院,农业农村部淡水水产种质资源重点实验室,上海 201306;2.盐城师范学院 湿地学院,江苏省盐土生物资源研究重点实验室,江苏 盐城 224002 )
中国许多城市河道与湖泊都面临水体氮、磷超标问题[1-2]。刚毛藻(Cladophora)属绿藻纲刚毛藻科,对外界环境耐受程度大,具有很强的适应能力,使其在复杂的污水环境中生长成为可能,刚毛藻分布广泛,作为净水植物便于就地取材,避免了物种引入造成的生物入侵问题[3-4]。此外,刚毛藻具有快速繁殖和易于回收的特点,具有人为可控性,利用刚毛藻去除污水中的氮、磷具有经济环保和可控性强的优势[5]。因此,在目前大型净水丝状藻类中,刚毛藻是理想的净水物种之一[6]。但在虾、蟹养殖过程中,刚毛藻大量繁殖不仅会占据水体空间、遮挡光照,影响水草、微藻的生长,缠绕虾蟹,而且老化腐烂后会引起水质恶化、溶解氧下降等问题,引起虾、蟹死亡[7-9]。
氮、磷对刚毛藻的生长至关重要[10],尤其是氮[11]。马沛明[12]通过试验证明,刚毛藻优先利用氨氮。本试验中,笔者选择氨氮作为氮源来研究不同环境因子对刚毛藻吸收氮的影响,以期通过调控环境因子来控制刚毛藻吸收氮,进而为人为调控刚毛藻生物量提供参考。
1 材料与方法
1.1 藻种采集与鉴定
试验用藻种采自上海海洋大学崇明基地河蟹塘。将采回的新鲜藻体除杂、反复清洗,置于光照培养箱中用纯水暂养。用显微镜观察法和分子方法(ITS-9F5′-CCGCCCGTCGCTCCTACCGATTGGGTGTG-3′;ITS-7R5′-TCCCTTTTCGCTCGCCGTTACTA-3′)[13]进行种类鉴定。
1.2 试验仪器与测定方法
1.2.1 试验仪器
MGZ-120L-3三温区光照培养箱(上海丙林电子科技有限公司)、紫外分光光度计(北京普析通用T6新世纪)、显微镜(奥林巴斯CX33)等。
1.2.2 测定方法
氨氮的测定采用纳氏试剂分光光度法[14];谷氨酰胺合成酶活性、超氧化物歧化酶活性、总抗氧化能力的测定均采用南京建成科技有限公司试剂盒进行。
1.3 试验方法
1.3.1 试验时间程和氨氮浓度选择
为确定合理的取样时间和处理浓度,采取多氨氮浓度梯度及多时间点取样法。将暂养的刚毛藻置于装有无氮BG11培养基(用氯化铁代替柠檬酸铁铵,排除铵根离子的影响)的塑料培养箱中,然后氮饥饿培养48 h。选择氯化铵作为氮源,在磷浓度为10 μmol/L条件下,根据预试验结果,设置6个氨氮浓度组:20、40、80、120、180 μmol/L和300 μmol/L ,培养液体积为1 L,对照组不放藻,每组3个平行。用吸水纸将刚毛藻表面水分吸干,称取1.0 g放入烧杯中,及时用玻璃棒将其搅拌分散,使其均匀分布在烧杯中,然后放入光照培养箱中培养(温度25 ℃、pH 8、光照度4500 lx)。在放入藻后的0、0.5、1.0、1.5、2.5、3.5、5.0、7.0 h分别取水样5 mL,用分光光度法测定水样中氨氮浓度,并计算氨氮的吸收速率。
1.3.2 正交试验
如上所述,对清洗的刚毛藻进行氮饥饿48 h。在刚毛藻稳定吸收氨氮的时间程和浓度下,选择温度、pH、光照度3个环境因子,按照表1进行正交试验设计。温度、光照度通过光照培养箱进行调节,pH通过0.10 μmol/L的氢氧化钠和盐酸进行调节。培养液体积和刚毛藻的添加量与上述试验相同,过程中不曝气。在0 h和1.5 h取水样测定氨氮含量,计算刚毛藻在0~1.5 h时的氨氮吸收速率。在1.5 h时采集刚毛藻样品,测定藻体的谷氨酰胺合成酶活性、超氧化物歧化酶活性、总抗氧化能力。
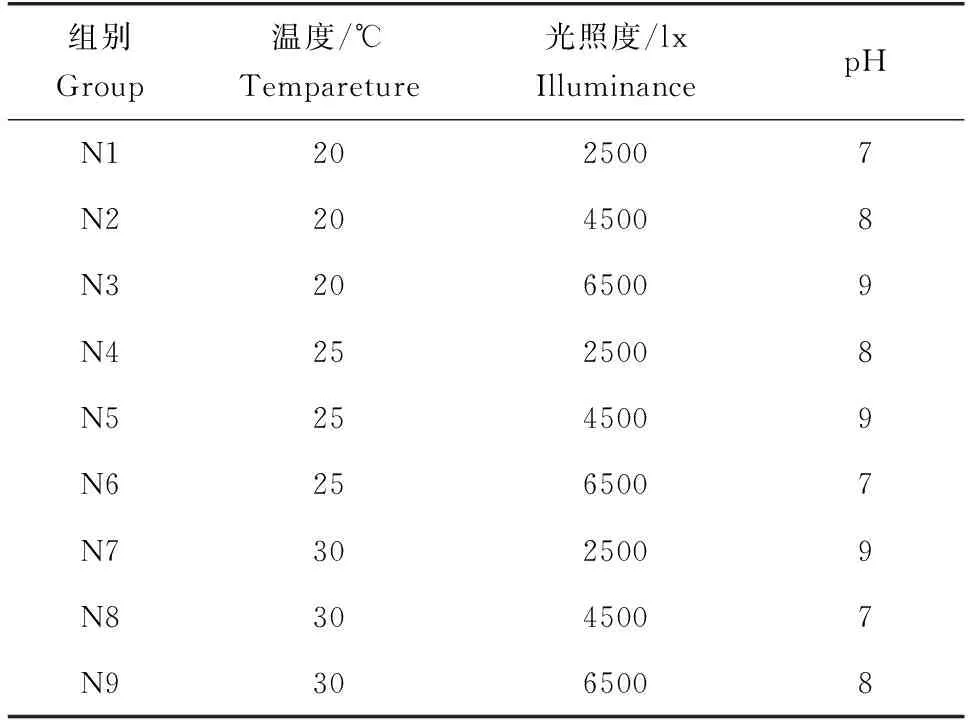
表1 正交试验设计[L9(33)正交表]
1.4 数据处理与分析
刚毛藻对氨氮的吸收速率计算公式如下:
v=[(c1-c2)V]/(m·t)
式中,v为吸收速率[μmol/(g·h)],c1为前一次取样浓度(μmol/L),c2为后一次取样浓度(μmol/L),V为前一次取样时培养液的体积(L),m为刚毛藻的湿质量(g),t为两次取样的时间间隔(h)。
用Excel计算每组试验的平均值和标准误差,用SPSS 19.0对氨氮的吸收速率和谷氨酰胺合成酶活性进行单因素方差分析。对正交试验结果进行多因素方差分析,P<0.05表示差异显著。
2 结 果
2.1 刚毛藻种类鉴定
通过显微观察、图谱比对,发现该试验用藻类为长筒形[(22~28) μm×(83~272) μm],具分支,叶绿体网状贴壁,淀粉核、细胞核多个,其与刚毛藻属物种相似性最高。将PCR扩增获得的核糖体ITS序列提交到GeneBank中进行检索,结果发现其与刚毛藻(Cladophorasp.)ZZ-2016(系列号KU186059.1)的相似性高达98.89%,基于此鉴定其为绿藻门刚毛藻属物种。
2.2 氨氮线性吸收时间程和适宜浓度
培养介质中氨氮浓度整体上不断下降(图1a)。在0.5~1.5 h时,培养介质中氨氮浓度快速下降,而在1.5~7.0 h时,刚毛藻对氨氮的吸收速率逐渐减慢。对比不同浓度组,在氨氮初始浓度为80 μmol/L、0.5~1.5 h时间程内,刚毛藻对氨氮吸收呈线性关系(r2>0.99)。不同氨氮浓度组吸收速率随时间变化的曲线见图1b,在氨氮初始浓度为80 μmol/L、时间程为0.5~1.5 h时,0.5~1.0 h和1.0~1.5 h时间程的吸收速率均约为45.10 μmol/(g·h),吸收速率基本相等。另外,由图1b可见,在0~0.5 h时,刚毛藻对氨氮的吸收速率较小。从整体上看,刚毛藻对氨氮的吸收速率呈现波动式变化。

图1 刚毛藻对氨氮的吸收Fig. 1 Absorption of ammonia nitrogen by Cladophora
2.3 正交试验
2.3.1 氨氮的吸收速率
根据上述试验结果,选择氨氮初始浓度为80 μmol/L的培养介质进行正交试验,采样时间在1.5 h。各试验组中刚毛藻对氨氮的吸收速率见图2a,其中N5组刚毛藻对氨氮的吸收速率最高,而N8组的吸收速率最低。多因素方差分析结果(表2)显示,温度对刚毛藻吸收氨氮的速率有显著影响(P<0.05),而pH和光照强度则无显著影响(P>0.05),3个因子的F值大小排序为:温度> pH>光照度。
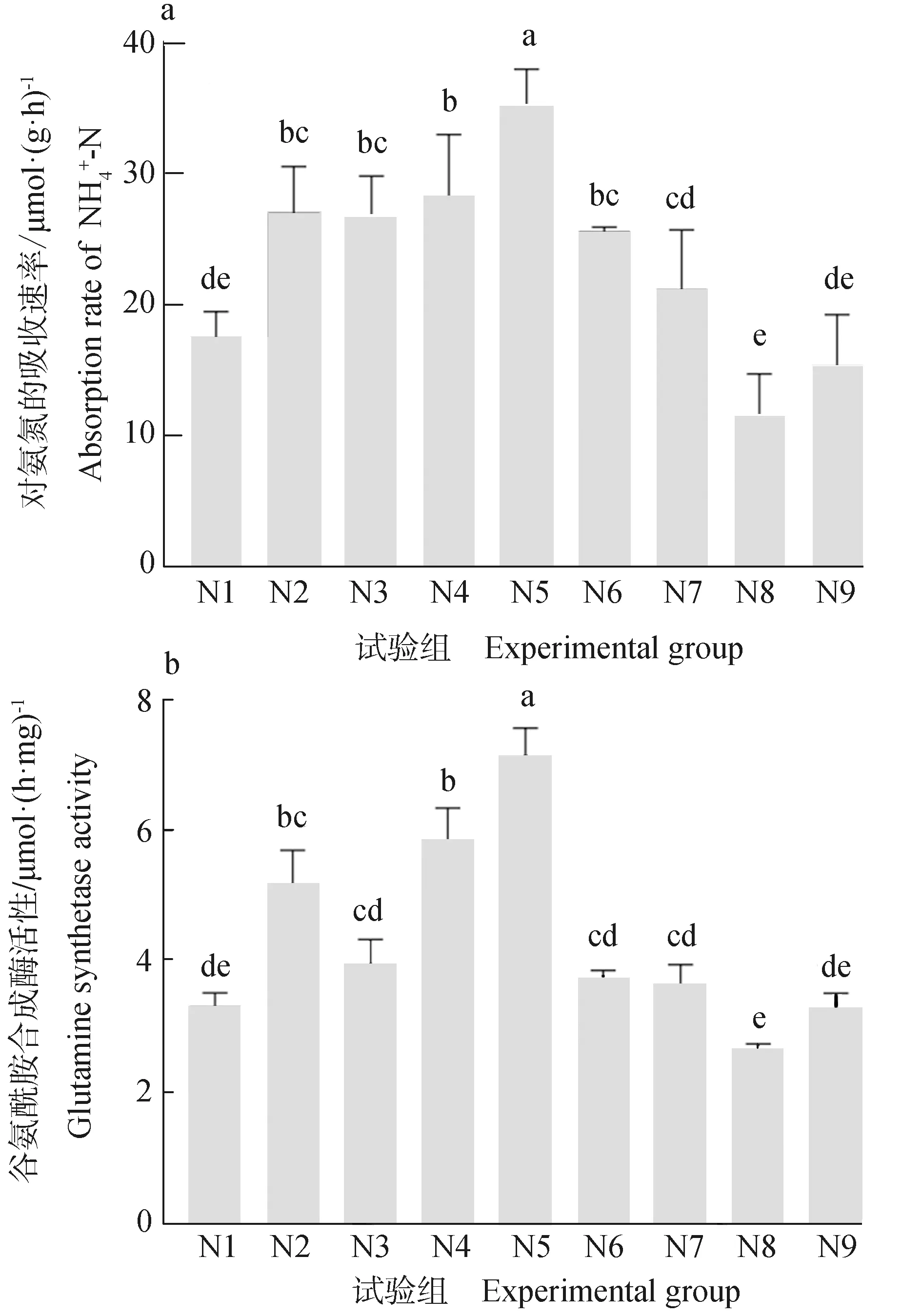
图2 不同试验组刚毛藻氨氮吸收速率及对应的谷氨酰胺合成酶活性Fig. 2 The absorption rate of NH4+-N of Cladophora and corresponding GS activity in different experimental groups

表2 多因素方差分析(吸收速率)
2.3.2 酶活性
N5组的谷氨酰胺合成酶活性最高,而N8组的谷氨酰胺合成酶活性最低(图2b)。另外,除N3组谷氨酰胺合成酶活性略低,谷氨酰胺合成酶曲线与氨氮吸收速率曲线整体走势相似。对谷氨酰胺合成酶活性与氨氮吸收速率进行皮尔逊相关分析发现,谷氨酰胺合成酶活性与氨氮吸收速率呈极显著正相关(P=0.001<0.01,r=0.898)。不同组超氧化物歧化酶活性和总抗氧化能力结果检测表明,两组曲线整体上走势相似,最低值均出现在N5组,而超氧化物歧化酶活性最高值出现在N7组,总抗氧化能力最高值出现在N8组(图3)。分别对超氧化物歧化酶活性和总抗氧化能力与氨氮吸收速率进行皮尔逊相关分析发现,超氧化物歧化酶活性与氨氮吸收速率虽然呈负相关关系,但不呈显著负相关(P=0.108>0.05,r=-0.572),而总抗氧化能力与氨氮吸收速率则呈显著负相关(P=0.029<0.05,r=-0.719)。
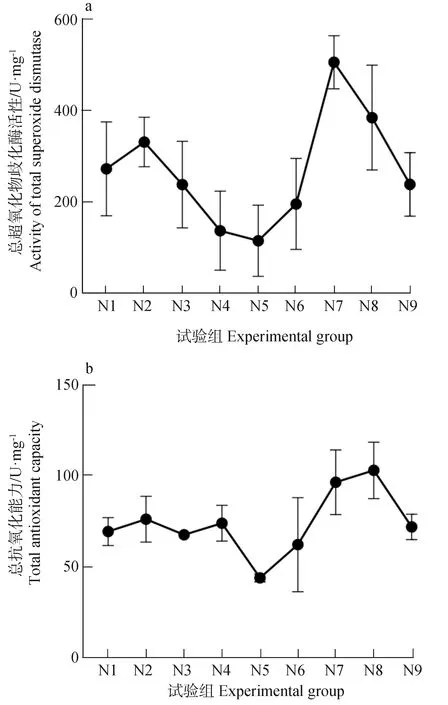
图3 不同试验组的总超氧化物歧化酶活性和总抗氧化能力Fig. 3 The total SOD activity and T-AOC in different experimental groups
3 讨 论
3.1 试验时间程和氨氮浓度选择
在研究环境因子如何影响藻类吸收营养时,确定适宜的取样时间和浓度对准确获取试验结果至关重要。在本试验中,初步确定时间程为0.5~1.5 h、氨氮浓度为80 μmol/L时,刚毛藻吸收氨氮效果较好。目前,有关淡水刚毛藻对氨氮稳定吸收的时间程和适宜浓度尚未见报道,李俭平[15]曾研究了浒苔(Enteromorphaprolifera)对氨氮的吸收动力学,指出浒苔在0~1.5 h时呈线性吸收。本试验中,刚毛藻在0~0.5 h时对氨氮的吸收速率处于较低水平,而在0.5~1.5 h时则呈现较高的线性吸收水平。笔者推测,0~0.5 h时间程可能是刚毛藻吸收氮有关酶的反应时间。本试验中,初步确定刚毛藻在0.5~1.5 h时间程内对氨氮呈线性吸收,且氨氮浓度为80 μmol/L时刚毛藻吸收氮效果较好。曹学彬等[16]对刚毛藻吸收营养盐的研究也发现,放入刚毛藻后,环境中的营养盐浓度不会立即降低,其初入新环境需要适应生长时间。方焰星等[17]研究了水生植物的除氮效果,发现金鱼藻(Ceratophyllumdemersum)、眼子菜(Potamogetondistinctus)等对氨氮去除效果较好,效率约为8.0 μg/(g•L•h)。陈巧玲等[18]研究发现,挺水植物水生美人蕉(Cannaglauca)对氨氮的吸收效率约为0.13 μg/(g•L•h)。本试验中,添加刚毛藻后7 h内溶液中氨氮的浓度从80 μmol/L降至0 μmol/L,对氨氮的吸收效率达到160 μg/(g•L•h),说明刚毛藻具有较强的吸收氮能力,是处理污水的理想物种之一。另外,也解释了在自然水域中为何刚毛藻具有明显的营养竞争优势。狄文亮等[19]研究也发现,添加刚毛藻对表流湿地净水能力有明显提升,而且金鱼藻受到了刚毛藻因营养盐竞争而造成的胁迫影响。
3.2 不同环境因子交叉作用对刚毛藻吸收氮的影响
多因素方差分析的F值可用于比较不同环境因子的影响强弱[20]。在上述浓度梯度下,不同环境因子对刚毛藻氨氮吸收速率的影响强弱排序为:温度>pH>光照度。胡帅龙等[21]也发现,温度对营养盐的吸收有重要影响,在初始总磷质量浓度低于2 mg/L、温度为20~25 ℃时,刚毛藻的总磷去除率可高达93.66%。然而,上述结论不同于李俭平[15]有关环境因子对浒苔氨氮吸收速率的影响强弱排序(光照度>温度>pH),原因可能是海水与淡水环境中不同刚毛藻种类之间存在差异。叶碧碧等[22]对刚毛藻氮、磷吸收研究发现,20 ℃、光照度3000 lx更有利于氮、磷的吸收。试验中发现,N5组的氨氮吸收速率最高,即环境条件为:25 ℃、pH 9、光照度4500 lx。本试验中的温度和光照值与曾令昭[23]的研究结果相似。关于pH对氮吸收的影响,马淑娟等[24]发现,碱性环境更有利于刚毛藻脱氮除磷。周军[25]发现:pH 8是影响藻类膜去除污水中氮的临界值;当pH<8时,对氨氮的去除效果有限;当pH>8时,对氨氮的去除效果明显。这与本试验结论相似,同时说明pH是影响藻类氮吸收的重要因素之一。
3.3 酶活性与氮吸收的关系
谷氨酰胺合成酶在高等植物氮代谢中至关重要,是氮代谢的关键酶。无机氮必须同化为谷氨酰胺和谷氨酸等有机氮才能为植物体所吸收和利用,而谷氨酰胺合成酶是这一氨同化过程的关键酶[26]。若无谷氨酰胺合成酶的调控或调控紊乱,则植物体氮代谢紊乱,进而影响植物的光合作用和生理代谢等方面[27]。颜昌宙等[28]研究了不同浓度氨氮对轮叶黑藻(Hydrillaverticillata)的生理影响,发现在高浓度氨氮条件下谷氨酰胺合成酶活性增加显著。氨氮吸收速率与谷氨酰胺合成酶活性的这种明确的响应关系,内在生理原因是氨氮在谷氨酰胺合成酶和腺苷三磷酸的参与下,与谷氨酸反应,生成谷氨酰胺,进而在其他酶的参与下再合成其他类型的氨基酸[29],也即谷氨酰胺合成酶是氨氮由无机氮转化成可利用有机氮的第一种酶,也是必不可少的酶。本试验中,谷氨酰胺合成酶活性与氨氮吸收速率的极显著正相关关系证实了氨氮吸收速率与谷氨酰胺合成酶活性的响应关系,显示了谷氨酰胺合成酶在刚毛藻吸收氮中起重要作用。另外,本试验结果显示,只有温度对氨氮吸收速率有显著影响,推测可能是因为相对于pH和光照度,谷氨酰胺合成酶对温度更加敏感。
超氧化物歧化酶作为重要的抗氧化酶,是植物细胞的第一道抗氧化防线[16],具有清除活性氧自由基的功能。机体防御体系中总抗氧化能力的作用主要是维持内环境活性氧的动态平衡,清除过高的活性氧,使机体处于氧化还原相对稳定的状态,代表体内酶类和非酶类抗氧化物的总体水平[30]。超氧化物歧化酶和总抗氧化能力均是自由基损伤和酶促、非酶促系统清除氧自由基、抗氧化的敏感指标[31]。不利的环境条件会引发藻体的抗逆反应,表现为抗逆性酶活性等升高,而升高的超氧化物歧化酶活性和总抗氧化能力指标则反映了机体受胁迫的程度。如果将不同环境因子下的刚毛藻超氧化物歧化酶活性和总抗氧化能力指标与刚毛藻氨氮的吸收速率进行相关性分析,则可以间接反映环境因子与刚毛藻吸收氮的关系。如上文所述,超氧化物歧化酶活性与氨氮吸收速率呈非显著负相关,说明超氧化物歧化酶活性的升高能阻碍刚毛藻的氨氮吸收速率,但不是唯一途径或起主导作用。本试验中,最高活性超氧化物歧化酶组并没有对应最低的氨氮吸收速率即证明了上述推论。但总抗氧化能力与氨氮吸收速率呈显著负相关,说明总抗氧化能力的升高能显著阻碍刚毛藻对氨氮的吸收。对比超氧化物歧化酶和总抗氧化能力可知,超氧化物歧化酶仅是抗逆性酶的一种,而总抗氧化能力代表体内酶类和非酶类抗氧化物的总体水平,说明环境因子对刚毛藻氨氮吸收的影响不是仅通过某一种酶发挥作用,更可能是通过多种酶以及非酶类发挥影响作用。廖秀睿等[32]对曲褶刚毛藻(C.flexuosa)无机氮(氨氮、亚硝态氮、硝态氮)去除的研究表明,同为小分子无机氮,曲褶刚毛藻对氨氮的去除效率最高,原因可能是由于刚毛藻对氨氮吸收存在酶或非酶等多种调控途径,使得其具有较高的吸收率。
氮作为刚毛藻生长所需的重要营养盐,与其生物量大小直接相关[33]。笔者确定了刚毛藻吸收氮的最佳环境因子组合,试验结果为调控刚毛藻的生长或生物量提供了依据,同时可为预测和抑制刚毛藻暴发和净化污水提供参考。
4 结 论
在氨氮浓度为80 μmol/L、时间程为0.5~1.5 h时,刚毛藻对氨氮呈线性吸收(r2>0.99)。温度对刚毛藻氨氮吸收有显著影响(P<0.05),而光照度和pH无显著影响(P>0.05)。上述3种环境因子对刚毛藻吸收氨氮的影响顺序为:温度>pH>光照度。刚毛藻对氨氮吸收的最佳3因子组合为:25 ℃、pH 9、光照度4500 lx。另外,谷氨酰胺合成酶活性与刚毛藻氨氮吸收速率呈极显著相关(P<0.01),超氧化物歧化酶活性与氨氮吸收速率呈负相关 (r=-0.572),而总抗氧化能力与氨氮吸收速率呈显著负相关(P=0.029<0.05,r=-0.719),说明环境因子通过多种酶以及非酶类影响刚毛藻吸收氮。
