小球藻对草鱼生长、免疫、消化酶活性和肌肉的影响
2023-09-27阙江龙徐先栋丁立云章海鑫张爱芳李艳芳
阙江龙,饶 毅,徐先栋,丁立云,章海鑫,张爱芳,李艳芳
( 1.江西省水产科学研究所,江西 南昌 330039; 2.江西省水生生物保护救助中心,江西 南昌 330096 )
草鱼(Ctenopharyngodonidella)因肉质鲜美、生长速度快、单位产量高、养殖成本低等优势备受养殖户青睐,是我国最主要的养殖鱼类。据《2021中国渔业统计年鉴》统计,2020年草鱼产量达5.57×106t,是我国养殖量最大的淡水鱼类[1]。
水产养殖中,饲料添加剂的应用不仅可缩短养殖周期,还可以提高养殖动物的生产水平和养殖经济效益[2]。为了解饲料添加剂对草鱼的生长作用,学者们在饲料中添加油脂[3]、生物素[4]、维生素[5]、植酸酶和蛋白酶[6]等,研究其对草鱼生长性能、肌肉品质等影响,结果表明各饲料添加剂对草鱼生长均有一定的促进作用。然而,在草鱼饲料中添加小球藻(Chlorella)的研究尚未见报道。
小球藻为绿藻门单细胞小型藻类,细胞呈球形或椭球形,在自然界分布广泛。小球藻有着很高的光合作用效率,生长繁殖速度极快,是动植物中唯一能在20 h增长4倍的生物[7],因其体内含有丰富的胡萝卜素、多糖、脂类、微量元素、蛋白质、不饱和脂肪酸和生物生长因子等高价值的化合物[8-11],应用价值非常高。目前,小球藻在水产饲料方面应用广泛,Mahmoud等[12-14]分别在尼罗罗非鱼(Oreochromisniloticus)、罗氏沼虾(Macrobrachiumrosen-bergii)和黄颡鱼(Pelteobagrusfulvidraco)饲料中添加小球藻,发现适量添加小球藻对机体生长或消化酶活性等起到促进作用。笔者通过在饲料中添加不同水平的小球藻,探讨其对草鱼生长性能、非特异性免疫机能、消化酶活性及肌肉品质的影响,旨在为开发适合草鱼养殖的小球藻饲料提供科学依据。
1 材料与方法
1.1 试验鱼
试验于2020年10月在江西省水产科学研究所养殖鱼池进行,试验鱼为平均体质量(105.92±2.79) g、体长(18.46±0.57) cm、体格健壮、规格整齐的450尾6月龄草鱼。
1.2 试验饲料及小球藻
试验所用小球藻购自深圳永晨实业有限公司生产的破壁蛋白核小球藻(C.pyrenoidosa)粉,粗蛋白59.1%,粗脂肪13.4%,灰分5.59%。试验基础饲料为自行配制,粗蛋白为28.7%,粗脂肪为7.6%(表1)。小球藻的添加量分别为0(对照组)、0.25%、0.50%、0.75%和1.00%,配制成5组试验饲料。饲料原料经粉碎后过80目筛,按配比定量后混合均匀,加入适量的水揉匀,经饲料加工机制成颗粒料,65 ℃烘干后贮存备用。

表1 基础饲料配方 %

表2 小球藻对草鱼生长的影响(n=30)
1.3 试验管理
养殖试验在江西省水产科学研究所循环水系统中进行。试验前,用2×10-5kg/L的高锰酸钾对养殖用聚乙烯塑料圆桶(直径80 cm)进行消毒。用质量分数3%食盐溶液药浴试验鱼10 min,后将试验鱼放入圆桶中暂养10 d,暂养期间每日投喂自行配制的基础饲料2次(9:00和16:00)。试验设置5个试验组,每组3桶,每桶放30尾试验鱼。每组3个平行,每日9:00和16:00投喂。投喂量为草鱼体质量的3%,视摄食情况作调整。试验期间水温24~26 ℃,24 h充气,溶解氧质量浓度7.0~8.0 mg/L,pH7.2~7.5。由室内日光灯提供光照,每日光照10 h,饲养试验时间28 d。
1.4 生长指标测定
饲养试验结束后,将试验鱼饥饿处理24 h后捞出,称量体质量,计算质量增加率(wWGR,%)、特定生长率(RSG,%/d)和饲料系数(RFC)[15]。
wWGR=(mt-m0)/m0×100%
RSG=(lnmt-lnm0)/t×100%
RFC=mF/(mt-m0)
式中,m0为鱼的初始总质量(g),mt为鱼的终末总质量(g),t为试验天数(d),mF为饲料投喂量(g)。
1.5 非特异性免疫机能和消化酶活性测定
自每个平行随机取6尾鱼进行尾静脉采血,冰上解剖提取肠道,血清和肠道分别置于1.5 mL灭菌的离心管中,3000 r/min(离心半径10 cm)离心2 min,制备血清样品保存于-80 ℃冰箱中,肠道样品于4 ℃保存。血清总蛋白含量使用日立7600-110型全自动生化分析仪进行测定。溶菌酶、碱性磷酸酶、超氧化物歧化酶、脂肪酶、胰蛋白酶、淀粉酶活性和总抗氧化能力采用南京建成生物工程研究所试剂盒检测,按说明书要求操作并计算。
1.6 肌肉氨基酸和脂肪酸测定
每组取3尾鱼的背部两侧肌肉,冰上剪碎后混匀,存放于干冰中寄送至江西省分析测试中心,按GB/T 14965—1994的方法使用Biochrom 20型氨基酸自动分析仪测定氨基酸,按GB/T 5009.168—2003的方法使用Agilent 6890型气相色谱仪测定脂肪酸[16]。
1.7 肌肉粗蛋白和粗脂肪测定
每组取3尾鱼的背部两侧肌肉,冰上剪碎后混匀,采用凯氏定氮法(GB/T 6432—1994)和索氏抽提法(GB 5009.6—2003)测定肌肉的粗蛋白和粗脂肪。
1.8 数据统计处理
试验结果用平均值±标准差表示,采用SPSS 26.0对数据进行单因素方差分析,并采用邓肯多重比较检验组间差异的显著性,P<0.05即认定为差异显著。
2 结果与分析
2.1 小球藻对草鱼生长性能的影响
试验结束后,各组试验鱼无死亡,成活率100%。在0.5%~1.0%添加组中,随着饲料中小球藻添加量的增加,草鱼的终末平均体质量、质量增加率和特定生长率均逐渐上升,饲料系数逐渐下降,与对照组差异显著(P<0.05)。当添加量为0.75%时,草鱼的终末平均体质量、质量增加率和特定生长率最高,显著高于对照组(0)和0.25%、0.50%添加组(P<0.05),饲料系数最低,显著低于对照组和0.25%、0.50%添加组(P<0.05)。当小球藻添加量达到1.00%时,草鱼的终末平均体质量、质量增加率和特定生长率又显著下降,但仍显著高于对照组,饲料系数虽然显著上升,但显著低于对照组(P<0.05)。
2.2 小球藻对草鱼血清非特异性免疫机能和抗氧化性能的影响
随着小球藻添加量的增加,草鱼血清的溶菌酶和碱性磷酸酶活性总体呈上升趋势,0.50%以上的添加组中溶菌酶活性与对照组差异显著(P<0.05),0.75%和1.00%添加组的碱性磷酸酶活性显著高于其他组(P<0.05)。各组间血清总蛋白含量差异不显著(表3)。
在血清抗氧化性能方面,超氧化物歧化酶活性和总抗氧化能力随着小球藻添加量的增加也呈上升趋势,0.50%、0.75%和1.00%添加组的超氧化物歧化酶活性和总抗氧化能力均显著高于对照组和0.25%添加组,且3组间差异不显著(表3)。
2.3 小球藻对草鱼肠道消化酶活性的影响
0.25%添加组淀粉酶活性与对照组无显著差异,但其他试验组皆显著低于对照组(P<0.05);小球藻添加组胰蛋白酶活性显著高于对照组,其中0.25%组最高,与其他各组差异显著(P<0.05);各添加组脂肪酶活性高于对照组,0.25%、0.75%和1.00%添加组水平与对照组差异显著(P<0.05)(表4)。
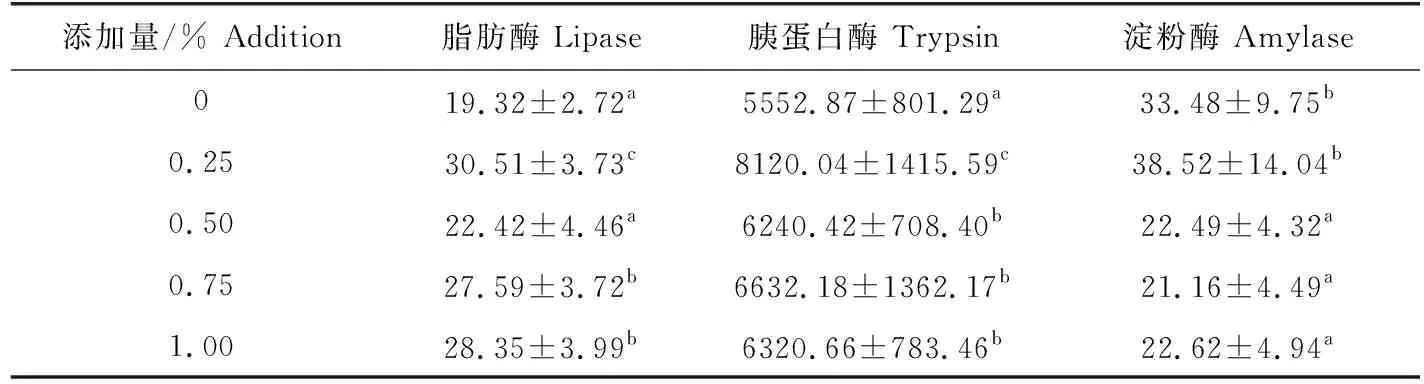
表4 小球藻对草鱼肠道消化酶活性的影响 U/g
2.4 小球藻对草鱼肌肉成分和品质的影响
各添加组草鱼的肌肉粗蛋白、粗脂肪、灰分和水分与对照组均无显著差异(表5)。在肌肉品质方面,小球藻各添加组草鱼的氨基酸组成与对照组也无显著差异(表6)。1.00%添加组中,4种不饱和脂肪酸即亚麻酸、花生四烯酸、二十碳五烯酸和二十二碳六烯酸显著高于对照组(P<0.05),分别比对照组高40.17%、21.20%、30.95%和46.70%,脂肪酸含量中,1.00%添加组显著高于对照组(P<0.05)(表7)。

表5 小球藻对草鱼肌肉成分的影响 %
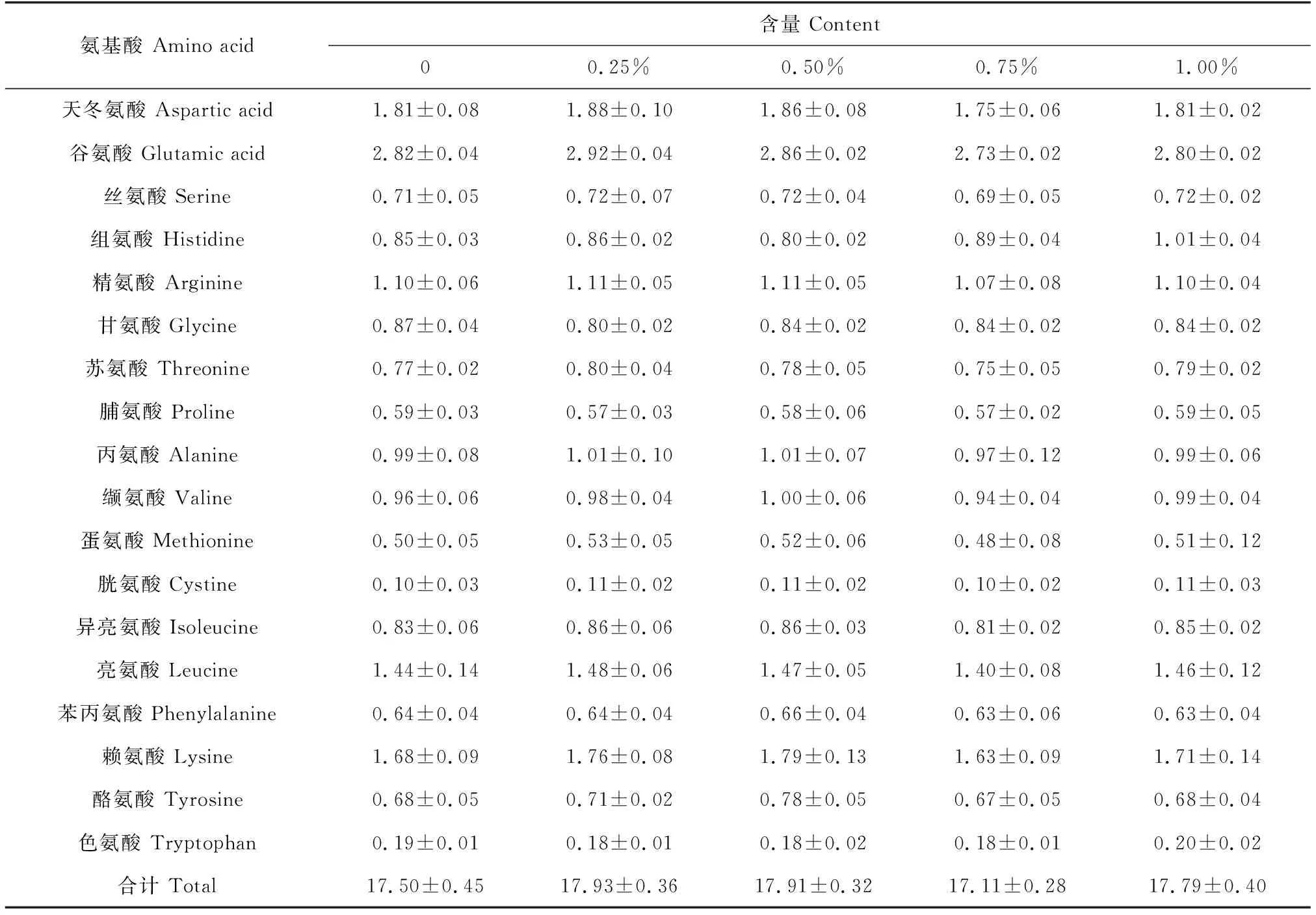
表6 小球藻对草鱼肌肉氨基酸的影响 %
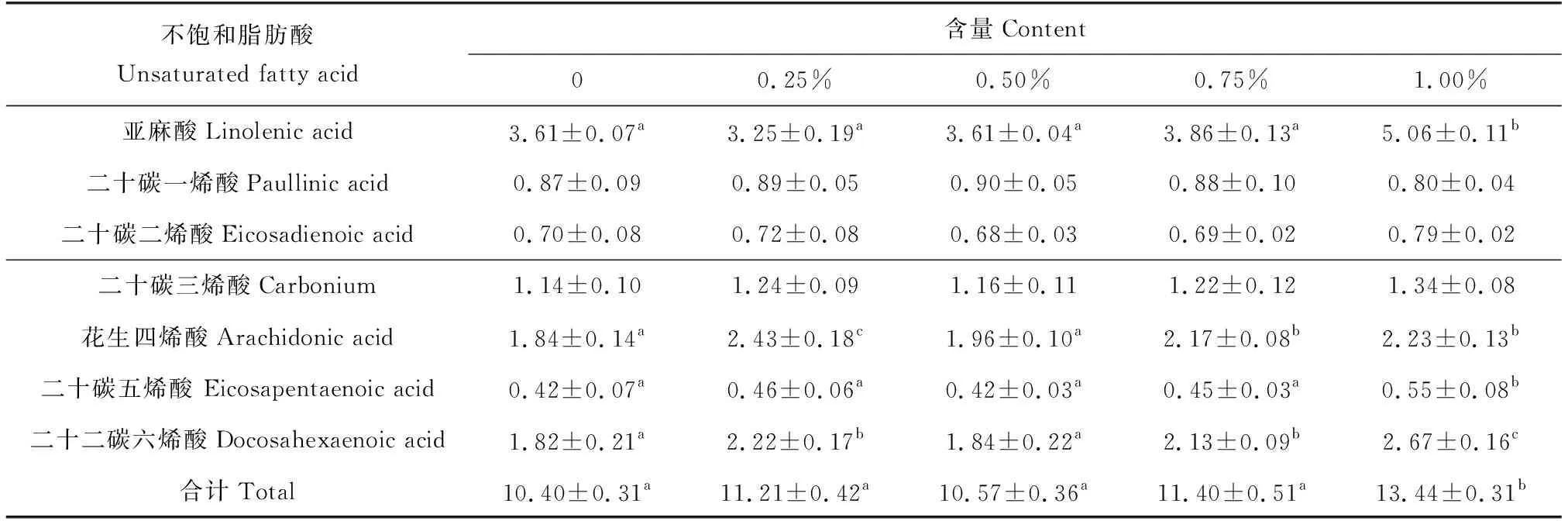
表7 小球藻对草鱼肌肉不饱和脂肪酸的影响 %
3 讨 论
3.1 小球藻对草鱼生长和成活的影响
添加组中试验鱼未出现死亡,与对照组一致,成活率均为100%,表明添加小球藻对草鱼的成活率无显著影响。
小球藻富含蛋白质,营养物质全面[10,17],可作为优质的添加剂应用于动物饲料中[18]。在饲料中加入一定量的小球藻,能增强仔猪对营养的消化吸收,促进生长[19],可增加鸡蛋中蛋黄色素含量,蛋壳变硬[20],肉鸡的质量增加率也会提高[21]。
水产饲料中,Bai等[22]在许氏平鲉(Sebastesschlegelii)饲料中添加0.5%的小球藻藻粉,发现其特定生长率、蛋白质效率明显提高。张宝龙等[14]在饲料中添加0%~3.2%的小球藻后,黄颡鱼质量增加率、特定生长率及蛋白质效率最高,且饵料系数最低;崔青曼等[23]也证实,饲料中添加小球藻能够显著促进凡纳滨对虾(Litopenaeusvannamei)生长。本试验结果显示,当饲料中小球藻粉添加量达0.50%以上时,草鱼的质量增加率、特定生长率显著高于对照组,饲料系数显著降低,与上述研究结果相似。表明饲料中添加适量小球藻可提高草鱼的生长率。
本试验中,小球藻添加组表现出良好的促生长效果,一方面可能是因为小球藻含较全面的营养物质,蛋白质中含有谷物类所缺乏的亮氨酸、精氨酸和赖氨酸[7],同时,破壁小球藻的蛋白质消化吸收率非常高(80%)[24]。饲料中添加小球藻,可解决植物蛋白营养价值低,达到“氨基酸的互补作用”的目的,满足草鱼生长要求。另一方面可能是小球藻所特有的促生长因子能增强机体免疫力,降低饲料消耗[25-26],促进草鱼的健康生长。
3.2 小球藻对草鱼血清非特异性免疫机能和抗氧化性能的影响
鱼体中溶菌酶和碱性磷酸酶是鱼类非特异性免疫的主要组成因子,可通过其活性的高低衡量机体免疫状态。溶菌酶是一类有溶菌活性的碱性蛋白,在水产动物抵抗病原微生物感染中具有重要作用[27-28]。与溶菌酶一样,碱性磷酸酶也是巨噬细胞杀菌的物质基础,它能形成水解酶体系,清除、破坏入侵的异物,在免疫中发挥重要作用[29]。Xu等[30]研究表明,小球藻添加于鲤(Cyprinuscarpio)的饲料中可提高其血清溶菌酶活性,溶菌酶活性随小球藻添加量的增加而升高。崔青曼等[23]在饲料中添加1%小球藻,发现小球藻能够显著或极显著提高凡纳滨对虾(Litopenaeusvannamei)血淋巴中的溶菌酶和碱性磷酸酶的活性(P<0.05),明显降低凡纳滨对虾的攻毒死亡率。本试验中,饲料中添加小球藻提高了草鱼血清的溶菌酶和碱性磷酸酶活性,当添加量达0.75%至1.00%时,提升水平显著(表3),表明饲料中添加小球藻能够增强草鱼的非特异性免疫机能。
超氧化物歧化酶是机体内重要的抗氧化酶,广泛存在于生物体的胞浆和线粒体基质中,能清除机体新陈代谢中产生的有毒物质,也是机体内部超氧自由基的清除因子[29-31]。Vijayavel等[32]研究表明,小球藻醇提物能提高小鼠超氧化物歧化酶活性和机体抗氧化能力。黄文庆等[15]对瓦氏黄颡鱼(P.vachelli)研究也得到了相同结论。崔青曼等[23]在凡纳滨对虾饲料中添加小球藻,血清中超氧化物歧化酶活性和抗氧化能力随添加水平的增加呈上升趋势。本试验结果显示,饲料中添加小球藻显著提高了超氧化物歧化酶活性和机体抗氧化能力,与这些研究结果一致。
3.3 小球藻对草鱼肠道消化酶活性的影响
鱼类消化酶活性的高低能反映鱼类对饲料消化利用的能力,消化酶活性越高,表明鱼类的消化吸收能力越强,饲养效果越好[33]。本试验中,添加组中草鱼肠道脂肪酶和胰蛋白酶活性均高于对照组,小球藻添加量达0.50%时,淀粉酶活性显著低于对照组,说明此时脂肪和蛋白质为草鱼提供更多的能量,脂肪酶和胰蛋白酶活性升高,碳水化合物作为能量物质的转换相对减少,淀粉酶活性降低。
Radhakrishnan等[13]研究发现,小球藻粉适量地替代饲料中鱼粉可提高罗氏沼虾消化道蛋白酶、脂肪酶和淀粉酶的活性。而黄文庆等[15]研究结果表明,饲料中添加小球藻醇提物对瓦氏黄颡鱼肠道内消化酶活性均无显著影响。本试验饲料中添加小球藻,草鱼脂肪酶和胰蛋白酶活性升高,淀粉酶(除0.25%添加组)活性显著减弱,与Radhakri-shnan等[13,15]研究结果存在差异性。这可能是因为鱼类消化酶不仅与其生活的水环境(温度、盐度、pH等)密切相关[34],而且还受其生理状况及摄食饵料的影响[35]。小球藻对草鱼消化酶活性的影响机理有待进一步研究。
3.4 小球藻对草鱼肌肉成分和品质的影响
小球藻粉对鱼体成分的影响存在差异。Kim等[36-37]研究发现,饲料中添加小球藻粉可提高褐牙鲆(Paralichthysolivaceus)机体粗蛋白质含量,降低机体粗脂肪含量。而黄文庆等[15]研究表明,饲料中添加0.25%~2.00%小球藻醇提物,对瓦氏黄颡鱼体成分中粗蛋白质、粗脂肪、粗灰分和水分含量均无显著影响。本试验中,小球藻添加组中草鱼的肌肉氨基酸含量与对照组无显著差异,与黄文庆等[15]研究结果一致。鱼体肌肉成分变化的不同,除鱼品种间存在差异外,也可能与鱼体的代谢方式及饲料蛋白源不同有关[38],本试验小球藻添加量最高为1.00%,与Kim等[36-37]的研究中小球藻粉添加量达4%不同,添加量不同可能是导致试验结果存在差异的原因之一。
小球藻中含有丰富的不饱和脂肪酸,如以亚麻酸、二十碳五烯酸和二十二碳六烯酸为代表的ω-3高度不饱和脂肪酸[39],它们是磷脂生物膜、视觉和神经系统等的结构组分,具重要的生物功能[40-41],是人类和动物生长发育的必需脂肪酸[42]。鱼体内不能自行合成亚麻酸、二十碳五烯酸和二十二碳六烯酸,而是通过摄入饵料后富集在体内聚集[16]。已有研究表明,投喂小球藻能增加轮虫和卤虫(Artemia)体内的二十碳五烯酸和二十二碳六烯酸的含量[7],张跃群等[43]研究证明,小球藻可以显著提高卤虫幼体二十碳五烯酸含量。本试验中,1.00%添加组的草鱼肌肉亚麻酸含量显著高于对照组;二十碳五烯酸含量高于对照组,其中,1.00%添加组与对照组差异显著;添加组(0.50%除外)二十二碳六烯酸含量显著高于对照组。说明草鱼饲料中添加小球藻能有效提高鱼肉中高度不饱和脂肪酸含量。
4 结 论
饲料中添加小球藻对草鱼的生长性能、非特异性免疫机能、抗氧化能力和肌肉品质均有提升促进作用,以质量增加率和饲料系数为评估指标,草鱼饲料中小球藻适宜添加量为0.75%。
