中药材黄精属的光合特性研究综述
2023-09-19杨丽姣涂海华孙佳佳邝先飞
杨丽姣,涂海华,孙佳佳,安 婷,邝先飞
(江西农业大学生物科学与工程学院,江西南昌 330045)
《中华人民共和国药典》(I部)中规定黄精、滇黄精和多花黄精是3种可以入药的中药材[1],其中多花黄精(Polygonatumcyrtonema)又名姜形黄精、南黄精、山姜、野生姜等,是百合科黄精属多年生草本植物,与其他品种相比,具有生长速度快、适应性强、药用成分高、经济价值高等特点。多花黄精以根茎入药,具有抗炎、抗肿瘤、抗病毒、抗真菌及解毒等功效及补气养阴、健脾益肾、降血糖、降血脂、保护心血管系统、调节和增强免疫力等保健功能[2],在新药研制和保健品开发等方面具有广阔的发展前景。随着市场需求量不断增加,以野生采集为主的多花黄精已不能满足市场的需求,开发优质高产的黄精人工栽培技术越来越被研究者关注。光照条件是植物生长发育最重要的外部条件,研究不同人工栽培条件下多花黄精的光合特性,提高多花黄精的产量,已成为高产高效栽培多花黄精亟待解决的问题,目前已有学者对林下栽培、大田栽培、盆栽种植3种生境条件下多花黄精的光合特性进行了探索。该研究综述不同地域、不同生境条件下的黄精属光合特性的研究进展,以期为黄精属的优质栽培提供技术参考。
1 研究文献的统计
以黄精、光合作用作为主题词,从CNKI数据库中检索到相关文献61篇,具体年度分布及2022年发文预测见图1。

图1 黄精光合作用文献统计及2022年预测
通过阅读筛选,选取密关的39篇文献进行分析。其中,高校发文量24篇,西北农林科技大学发文最多,为5篇;南京农业大学发文为4篇。研究院所等发文量为15篇,其中,安徽省林业科学研究院发文3篇。另外,发文中有18篇为多个异质单位跨区域合作完成。从地域上看,湖南发文6篇,陕西5篇,安徽、山东、江苏、浙江各4篇,重庆、福建各3篇,云南2篇,北京、吉林、河北、深圳各1篇。该研究对以上典型地域及单位作者对多花黄精光合特性的研究进行综述,既可以了解多花黄精光合特性的研究进展,也可以从一个侧面了解多花黄精人工栽培技术领域的研究动态和发展趋势。
2 不同区域黄精属的光合特性研究
2.1 湖南地区贺安娜等[3-4]测试了湖南溆浦和新晃2个黄精产区设置实验基地多花黄精及黄精的气体交换、叶绿素荧光参数,并比较其叶绿素含量和株叶形态特征。结果表明:①两地黄精的净光合速率、气孔导度、蒸腾速率、水分利用效率都高于多花黄精。②溆浦种植的黄精和多花黄精的Pn分别为5.82和2.65 μmol/(m2·s),都显著高于新晃栽培地,但溆浦种植的入药用的根茎产量明显低于新晃产地,由此得出了可多地种植高产、高光合黄精药材。刘熙等[5]以移植栽培的湖南省安化县及长沙县高桥基地3年生的7种黄精属植物(多花黄精、点花黄精、滇黄精、湖北黄精、黄精、卷叶黄精和长梗黄精)为材料,通过选取净光合速率、气孔导度、蒸腾速率、胞间CO2浓度5个光合指标进行测试、对比分析,发现不同种质黄精属植物的叶片净光合速率差异显著,这种差异主要源于胞间CO2浓度和叶片水分利用率。通过光合指标的聚类分析得出多花黄精、黄精和滇黄精光合性能最优,适合推广种植。刘其武等[6]选取湖南南山国家公园白云镇黄伞村厚朴人工林地的多花黄精为材料,测试多花黄精光合特性指标,结果与溆浦县、长沙、重庆、歌乐山、浙江临安等地的多花黄精光合性能指标相比较,发现南山公园的多花黄精光补偿点[3.001 μmol/(m2·s)]、暗呼吸速率[0.217 μmol/(m2·s)]、水分利用率[5.140 μmol/(m2·s)]等多个特征指标均优于其他地方,说明该区域自然条件下具有人工栽培多花黄精的优势。但测试发现,南山国家公园多花黄精的最大净光合速率[4.560 2 μmol/(m2·s)]低于其他地点的对比值,分析原因可能与测定的季节、时间和不同的计算方法有关。吴媛媛[7]以湖南省及周边县(市、区)的22个多花黄精种质作为研究材料,在系统研究后发现不同多花黄精指标光合特性日变化均呈单峰曲线,Pn、Gs、Ci、WUE、Ls峰值均出现在10:00,Tr峰值出现在12:00,正午出现Pn下降。且通过不同多花黄精种质的Pn-Ci响应曲线比较发现,提升CO2浓度可以有效提高多花黄精的光合能力。
2.2 陕西地区徐晓蓝等[8-9]以陕西省略阳县步长集团黄精规范化种植基地的3年生黄精为试验材料,系统研究了黄精不同叶位叶片的光合速率的差异及环境因子对光合速率的影响,结果发现,黄精叶位不同,净光合速率不同,随着叶位升高呈现“低-高-低”的规律,即中位叶[3.71 μmol/(m2·s)]>上位叶[3.05 μmol/(m2·s)]>下位叶[2.26 μmol/(m2·s)];不同叶位黄精叶片的日净光合速率呈“双峰型”曲线变化,且出现了明显的光合“午休”现象。贾向荣[10]以陕西省略阳县步长集团黄精规范化种植基地2年生黄精作为供试材料,测试了黄精种苗在不同水分(75%、50%、35%)条件下的生长、干物质和黄精多糖积累规律,并研究了低温胁迫条件下5个地区(陕西略阳、浙江仙居、四川雅安、河南篙县、河南灵宝)黄精生理指标的变化规律。结果发现,不同的水分条件下,黄精种苗的叶片净光合速率、气孔导度、瞬时水分利用率的变化均呈现明显的双峰曲线,并具有“午休现象”;低温胁迫对黄精的生长、发育和生存都会造成严重的影响。其中低温胁迫4 h时为黄精幼苗离体叶片生理指标响应的关键点;对比5个区域的黄精低温胁迫过程中离体叶片丙二醛(MDA,是膜脂过氧化最重要的产物之一,它的产生能加剧植物膜的损伤)含量测试结果发现,略阳当地的黄精自身调控抗低温胁迫能力强,具有很好的适应性。党康等[11]采用叶绿素荧光分析技术,对取自贵州省嵋潭县米山林场,培养于中国科学院水利部水土保持研究所大棚的黄精属3个不同种黄精、滇黄精、多花黄精的光合特性进行研究,发现黄精净光合速率(Pn)最高[4.91 μmol/(m2·s)],多花黄精[3.49 μmol/(m2·s)]其次,滇黄精最低[3.09 μmol/(m2·s)]。
2.3 江苏地区梁永富等[12-14]以盆栽的多花黄精为材料,分别研究了高温强光胁迫下水杨酸(0.5、1.0、1.5、2.0 mmol/L)对多花黄精生理及光合特性的影响、Ca2+对高温强光胁迫下多花黄精抗逆生理和光合特性的影响以及遮阴对多花黄精生长及光合特性的影响。研究发现,适量浓度(0.5~1.5 mmol/L)的水杨酸可使多花黄精能够在逆境胁迫下维持较高的光学活性和净光合速率,其中浓度为1.0 mmol/L时效果最佳;适宜浓度(10 mmol/L)的Ca2+处理即可有效减轻多花黄精的高温胁迫伤害,依然具有较高的净光合速率;不同梯度遮光处理(80%、60%、35%)下多花黄精的光合特性研究发现,遮阴促进了多花黄精叶片叶绿素a、叶绿素b和总叶绿素的合成,提高了净光合速率(Pn),但过度遮阴会降低光合产物向根茎的分配比例,60%的透光环境下多花黄精根茎产量和品质最好。邓艳婷[15]系统研究了土壤中矿质元素如Ca、Si对多花黄精生长产量和生理特性的影响,研究发现,轻度钙肥处理CaCl2(0.2%)多花黄精净光合速率最高,中度钙肥处理有利于提高多花黄精生长产量,0.6%时株高、茎粗、根茎干重达到最大值,单一钙肥处理不能提高黄精多糖含量;重度硅肥处理(10 mmol/L)能提高多花黄精抗氧化酶活性、叶绿素含量及光合参数,轻中度硅肥处理(4、6 mmol/L)能提高黄精多糖含量,中度钙硅互施处理(Na2SiO36 mmol/L+CaCl20.6%)能提高多花黄精抗氧化酶活性、叶绿素含量、光合作用,能显著提高黄精多糖含量,重度硅、轻度钙组合处理(Na2SiO310 mmol/L+CaCl20.2%)则利于提高多花黄精生长产量。
2.4 山东地区王晶晶等[16]以山东润竹山农业科技有限公司试验基地的大棚温室内的多花黄精为材料,对其光合特性进行了研究。研究发现,在多花黄精光合作用中,老叶是主要功能叶,老叶比新叶具有更高的叶绿素相对含量(老叶59.51、新叶33.67)、净光合速率[老叶2.986 μmol/(m2·s)、新叶1.422 μmol/(m2·s)]、胞间CO2浓度(老叶222.391 2 μmol/mol、新叶306.093 4 μmol/mol),黄精不同叶位叶片的净光合速率日变化均呈“双峰型”曲线,且有明显的“光合午休”现象。因此,生产上应当保存老叶、适当摘除部分新叶加以开发利用。高秋美等[17]选择4种不同透光率的皂角林地构造不同的遮阴处理(全光照、32.3%、52.6%、71.7%),研究了不同光照强度对多花黄精生长及光合特性的影响。结果表明,透光率为32.3%~71.7%的皂角林下环境比较适宜栽培多花黄精,有利于其产量和品质提高。李国清等[18-19]分别研究了不同遮阴处理(遮阴20%、40%、60%、80%,透光率19.4%、21.8%、40.0%)对泰山黄精光合作用、生长特性及产量的影响,发现遮阴40%和透光率40.0%条件下泰山黄精的光合作用表现最佳[最大净光合速率10.933 μmol/(m2·s),第11位叶9.60 μmol/(m2·s)],黄精产量最高。
2.5 其他地区骆绪美等[20-21]以安徽九华山地区采挖块茎所得的4种黄精属植物长梗黄精、多花黄精、黄精、玉竹为材料,研究了黄精属植物光合生理特性。结果发现,4种黄精属植物净光合速率(Pn)日变化均呈明显的单峰曲线,气孔导度、蒸腾速率与光合速率呈正相关,最大值均出现在13:00左右,表现为多花黄精>长梗黄精>黄精>玉竹,其中最大值多花黄精的净光合速率为7.8 μmol/(m2·s)。胞间CO2浓度与光合速率呈负相关;光合有效辐射、蒸腾速率及胞间CO2浓度均影响4种黄精属植物生长,其中光合有效辐射是主要因素;多花黄精为中性草本植物,长梗黄精、黄精、玉竹为中偏阳性草本植物。
童龙等[22-23]以重庆沙坪坝歌乐山盆栽多花黄精为材料,研究了不同层遮阴网光照下[全光照;1层遮阴网,透光率(83±5)%;2层遮阴网,透光率(61±5)%;3层遮阴网,透光率(38±5)%]多花黄精的光合特性差异,研究发现,遮阴提高了多花黄精光合色素质量分数,促进了叶片的光合作用,其中2层遮阴网光照处理的多花黄精净光合速率最大,适宜其健康生长。
袁名安等[24]对2层遮阴网环境下的多花黄精光合特性日变化进行研究。研究发现,多花黄精的净光合速率(Pn)、气孔导度(Gs)日变化曲线为“双峰”型,存在“午休现象”,峰值分别出现在11:00和16:00。影响多花黄精净光合速率的主要环境因子是大气光量子密度。李迎春等[25]研究了光照对多花黄精生长、光合和叶绿素荧光参数特征的影响,发现多花黄精在弱光环境下对蓝光利用能力强,具较强的耐荫特征,而且对光环境适应范围较宽,透光率45%~75%的条件下能良好生长。
陈芳软[26]采用福建泰宁县新垒生态农业有限公司的一年生多花黄精为材料,系统研究毛竹-杉木混交林下、大田栽培、盆栽种植3种生境条件下的多花黄精的生长特征及光合特性。结果发现,林下套种条件下多花黄精净光合速率在日变化进程中没有表现出“午休现象”,大田栽培与盆栽种植条件由于午间气温高,光合作用受到了明显的抑制,表现出较低的光合效率,比较而言,林下套种条件下的生长优于盆栽种植条件下的生长状况。章文前[27]分析了不同郁闭度和坡位的林下套种的多花黄精生长情况,认为郁闭度和坡位对多花黄精的株高、地径、根茎鲜重等生长指标均有影响,对根茎鲜重的影响最大。其中郁闭度对根茎鲜重影响为郁闭度0.6~0.8>0.4~0.6>0.8以上;坡位对根茎鲜重的影响为下坡位>中坡位>上坡位。因此,人工栽培多花黄精时要做好适度遮阳和水肥管理。
唐婧文等[28]通过盆栽控水试验,探讨土壤水分胁迫对多花黄精光合作用及叶绿素荧光参数的影响。结果表明,不同程度水淹胁迫对多花黄精的伤害明显,其中轻度水淹(1/4盆浸水)40 d后叶片开始变黄,中度水淹(1/2盆浸水)33 d后开始枯死,重度水淹(整盆浸水)14 d后全部枯死;不同程度干旱胁迫(土壤相对含水量20%、40%、60%、80%)对多花黄精的光合特性测试发现,多花黄精能忍受一定程度的干旱胁迫,土壤含水量60%~80%时多花黄精植物能够正常生长。
欧亚丽等[29]研究了遮阴对黄精光合特性和蒸腾速率的影响,得出了遮阴可以显著降低黄精的蒸腾速率,提高水分利用率的结论;闫莉等[30]研究发现,适度干旱和低养分投入可以促进黄精生长发育与黄精多糖累积,人工栽培黄精时适度的干旱及控制氮、钾等肥料用量,可以提升光合效能,保证黄精生长良好的同时提升黄精根茎生物量及多糖含量。
3 不同区域黄精属植物光合特性分析
3.1 不同区域多花黄精光合参数由表1可知,不同区域多花黄精净光合速率等光合参数存在差异。净光合速率(Pn)是植物光合作用强弱的重要指标,泰安市泰山林业科学院试验基地两层遮阳网(透光率40%)的泰山黄精净光合速率最大,湖南新晃无遮阴田间、山地的多花黄精净光合速率最小,极差达9.783 μmol/(m2·s)。气孔导度(Gs)是叶片对外界空气进入叶片细胞的能力指标,气孔导度越大,越有利于叶片细胞进行光合作用,泰安市泰山林业科学院试验基地两层遮阳网(透光率40%)的泰山黄精气孔导度最大,南山国家公园试验基地的多花黄精气孔导度最小,极差达0.102 3 mol/(m2·s)。胞间 CO2浓度(Ci)是叶片细胞内CO2的浓度,湖南溆浦无遮阴田间、山地的多花黄精胞间CO2浓度最大,南山国家公园试验基地的多花黄精胞间 CO2浓度最小,极差达225.616 7 mol/mol。蒸腾速率(Tr)是单位叶面积在一定时间内蒸腾的水量,湖南省农业科学院长沙县高桥基地栽培种的多花黄精蒸腾速率最大,山东润竹山农业科技有限公司试验基地的大棚温室内种植的多花黄精老叶(秋季叶)蒸腾速率最小,极差达1.67 mmol/(m2·s)。水分利用率(WUE)对光合作用有机物的积累具有重要影响,南山国家公园试验基地的多花黄精水分利用率最大,湖南新晃无遮阴田间、山地的多花黄精水分利用率最小,极差达4.246 mol/(m2·s)。
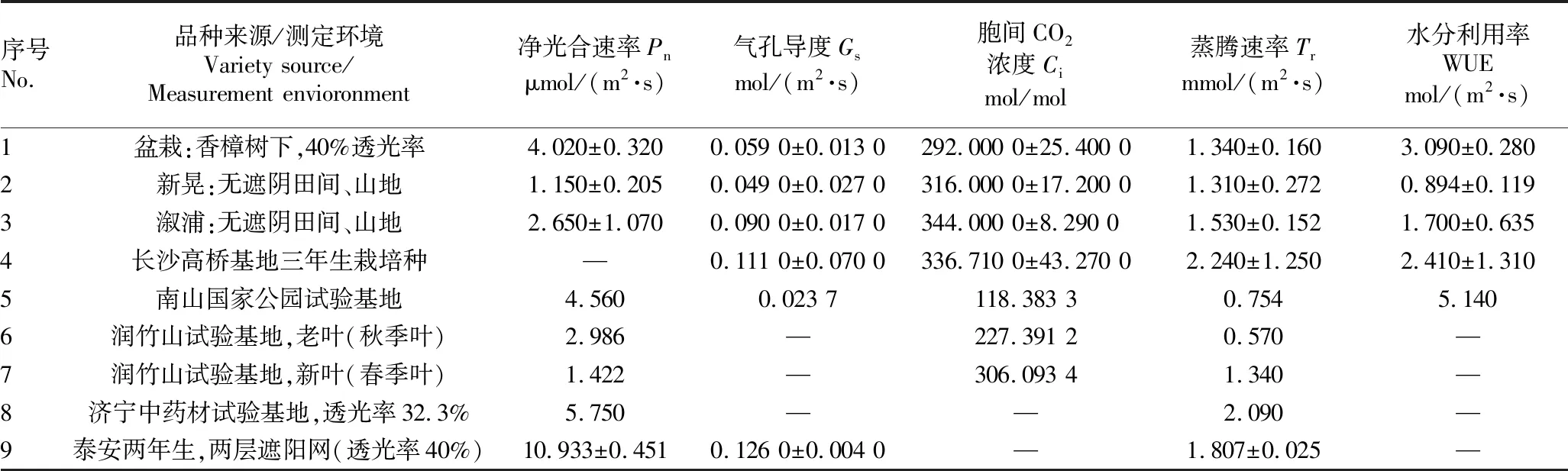
表1 不同区域多花黄精光合参数比较
3.2 不同区域黄精属植物(不含多花黄精)光合参数由表2可知,不同区域黄精属植物(不含多花黄精)净光合速率等光合参数存在差异。湖南溆浦无遮阴田间、山地的黄精净光合速率最大,湖南新晃无遮阴田间、山地的黄精净光合速率最小,极差达1.48 μmol/(m2·s)。湖南省农业科学院长沙县高桥基地栽培的点花黄精气孔导度最大,滇黄精气孔导度最小,前者为后者的5.8倍。湖南省农业科学院长沙县高桥基地栽培的湖北黄精胞间 CO2浓度最大,湖南新晃无遮阴田间、山地的黄精胞间 CO2浓度最小,极差达126 mol/mol。湖南省农业科学院长沙县高桥基地栽培的滇黄精蒸腾速率最大,长梗黄精蒸腾速率最小,极差达0.62 mmol/(m2·s)。湖南省农业科学院长沙县高桥基地栽培的湖北黄精水分利用率最大,长梗黄精水分利用率最小,前者为后者的2倍。
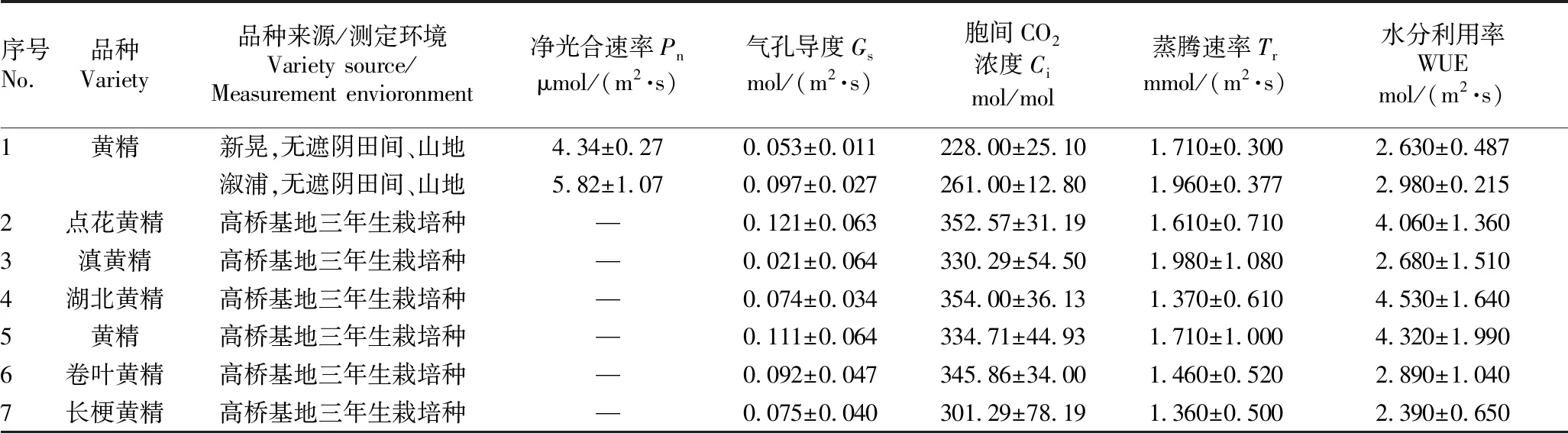
表2 不同区域黄精属植物(不含多花黄精)光合参数比较
3.3 不同区域多花黄精光合作用特征参数由表3可知,浙江省江山市盆栽的多花黄精在遮阳网(75%±5%)的条件下,最大净光合速率(Pmax)最高,南山国家公园试验基地内的多花黄精最大净光合速率最低;重庆市林业科学研究院内两层遮阴网(透光率61%±5%)下的多花黄精光饱和点(LSP)最高,浙江省江山市遮阳网[(45±5)%]条件盆栽的多花黄精光饱和点最低,二者光补偿点(LCP)相近,说明适宜的遮阴能提高多花黄精的光合作用效率;南山国家公园试验基地种植的多花黄精暗呼吸速率(Rd)最低,浙江省江山市盆栽多花黄精在遮阳网[(75±5)%]下暗呼吸速率最高,后者为前者的5倍,说明前者光合作用产物积累效率更高。

表3 不同区域多花黄精光合作用特性比较
4 结语
光是植物生长发育过程中必不可少的生态因子,光合作用直接影响植物的生长发育及生物量和有效物质的积累。综合已有研究发现,同种植地区、不同生境条件下人工栽培的多花黄精不同叶位的叶片,其光合速率存在差异,净光合速率日变化大多呈“双峰型”曲线,出现了明显的光合“午休”现象。而林下套种条件下多花黄精净光合速率在日变化进程中没有表现出“午休现象”,说明了大田栽培与盆栽种植条件下,光合作用由于午间气温高受到了明显的抑制,影响了光合效率,而林下套种条件下的生长则优于盆栽种植条件下的生长状况。
为缓解药用黄精品种短缺和野生资源枯竭现状,急需加快优质种质资源和培育研究。探索现有栽培条件下更利于调控多花黄精生长的光合因素,可为多花黄精的规范化、规模化栽培,为后续黄精属开发研究提供理论依据,助力中药材黄精属产业发展。
