长江口浮游动物群落结构时空变化特征研究
2023-09-14杨阳阳杨杰青王云龙
杨阳阳,杨杰青,王云龙,晁 敏
(1.上海海洋大学海洋生态与环境学院,上海 201306;2.中国水产科学研究院东海水产研究所,农业农村部东海渔业资源开发利用重点实验室,上海 200090)
长江口是中国主要的河口之一,受长江冲淡水、沿岸流系、台湾暖流等多重因素的影响[1-2],水文环境复杂多变,且受人类活动影响显著[3],是典型的河口生态区[4-5]。长江口拥有优越的自然条件,其丰富的营养物质以及较高的初级生产力为浮游动物的生长提供了优良条件。浮游动物在海洋生态系统中有着至关重要的作用,它是生态系统完整性的生物指标[6],其种类组成、生物量和次级生产力的变化都会对生态系统的结构和功能产生影响[7],并可对水环境的变化产生响应[8]。
有关长江口浮游动物群落结构变化的研究已有不少报道[9-15],这些研究分析了浮游动物群落结构的变化规律,表明盐度和温度[10,12-14]是影响浮游动物群落结构的主要因子,高倩等[13]、郑金秀等[14]都研究表明,温度是影响群落结构季节变化的主要因素,盐度是影响群落结构区域变化的决定因素;王晓东和李超伦[15]研究表明,浮游动物不同聚类组群的地理分布与海域的盐度梯度分布相对应;徐兆礼[12]研究表明,群落结构受到不同性质水团以及长江冲淡水的交互影响。上述研究大多针对某个季节或某几个航次,在较长时间序列上的群落结构研究较少。国外对河口浮游动物的研究也很多[16-21],如BURDIS和HIRSCH[16]使用聚类、排序等方法研究密西西比河浮游动物群落结构的时空变化特征,通过ANOSIM程序分析群落结构间是否存在差异性,采用SIMPER程序分析导致群落差异的贡献物种;STEFANIDIS和PAPASTERGIADOU[17]采用Kruskal-Wallis分析方法检验希腊湖泊中浮游动物丰度的差异性。本研究基于2006—2008年、2016—2018年两个时间段每年春季和夏季的调查资料,探讨浮游动物群落结构的时空变化,确定较长时间序列基础上的浮游动物生态地理划分特点,明确浮游动物的季节及年间变化,以期为更好地了解长江口及其邻近海域的生态变化提供科学参考。
1 材料与方法
1.1 资料来源及采样方法
数据来自农业农村部东海渔业生态环境监测中心2006—2008年和2016—2018年每年春季(5月)和夏季(8月)在长江口海域的调查资料。相比2006—2008年,2016—2018年站位设置有部分调整,因此只采用共有的11个站位资料,其中1~4站位位于口门区及最大浑浊带区域,5~11站位位于杭州湾(图1)。

图1 长江口调查海域及站位分布Fig.1 Investigation sea area and sam pling stations in the Yangtze Estuary
浮游动物标本采集使用浅水I型浮游生物网自底至表垂直拖拽,采集及标本处理均按照《海洋调查规范》(GB/T 12763.6—2007)[18]进行,经5%福尔马林溶液固定、保存,实验室内进行分类、鉴定和统计。本文浮游动物数据未统计水母类和海樽类,浮游动物丰度单位为:个·m-3。
1.2 统计分析
浮游动物优势种判别采用优势度公式:
式中,ni为第i种浮游动物丰度;N为浮游动物总丰度;fi为第i种浮游动物在各站位中的出现频率。优势度Y≥0.02的种为本文优势种。
以2006—2008年春、夏季和2016—2018年春、夏季各站每种浮游动物丰度均值数据为基础,用SPSS 26进行Kruskal-Wallis检验,分析种类组成和丰度差异[17];用PRIMER 5.0进行群落聚类、排序(nMDS分析)、ANOSIM和SIMPER分析,其中聚类和nMDS分析用于研究具有时空异质性的站位组[24],ANOSIM程序确认异质性[25],SIMPER程序确定对分组有主要贡献的种类[26]。用Surfer 14绘制站位图及浮游动物丰度空间分布图。
2 结果与分析
2.1 长江口环境概况
2006—2008年长江口平均径流量为7 624.6亿m3,2016—2018年平均径流量为9 285.3亿m3,2016—2018年流量明显增加。盐度春季高、夏季低,2016—2018年低于2006—2008年,且空间分布格局呈近岸低、外海高的变化趋势。水温春季低、夏季高,口门区及沿岸站位高于外海站位,夏季水温的空间分布格局与长江冲淡水的扩散方向相一致。
2.2 种类组成
12次航次共鉴定出浮游动物109种,分别隶属于5门14大类,包括枝角类、桡足类、端足类、磷虾类、十足类、等足类、糠虾类、涟虫类、介形类、翼足类、异足类、多毛类、毛颚类和被囊类,桡足类的种类数占绝对优势(表1)。其中,2006—2008年春、夏季6个航次共采集浮游动物13大类群73种,2016—2018年春、夏季6个航次共采集浮游动物10大类群63种。春季浮游动物种类数70种,夏季83种,2016—2018年春、夏季物种数相比2006—2008年均有所减少(图2)。Kruskal-Wallis检验结果表明,同一调查阶段不同季节间(春季和夏季)、同一季节不同调查阶段间(2006—2008年和2016—2018年)的类群物种数变化均不显著(P>0.05),但同一季节不同调查阶段间的站位物种数有显著变化(P<0.05)。
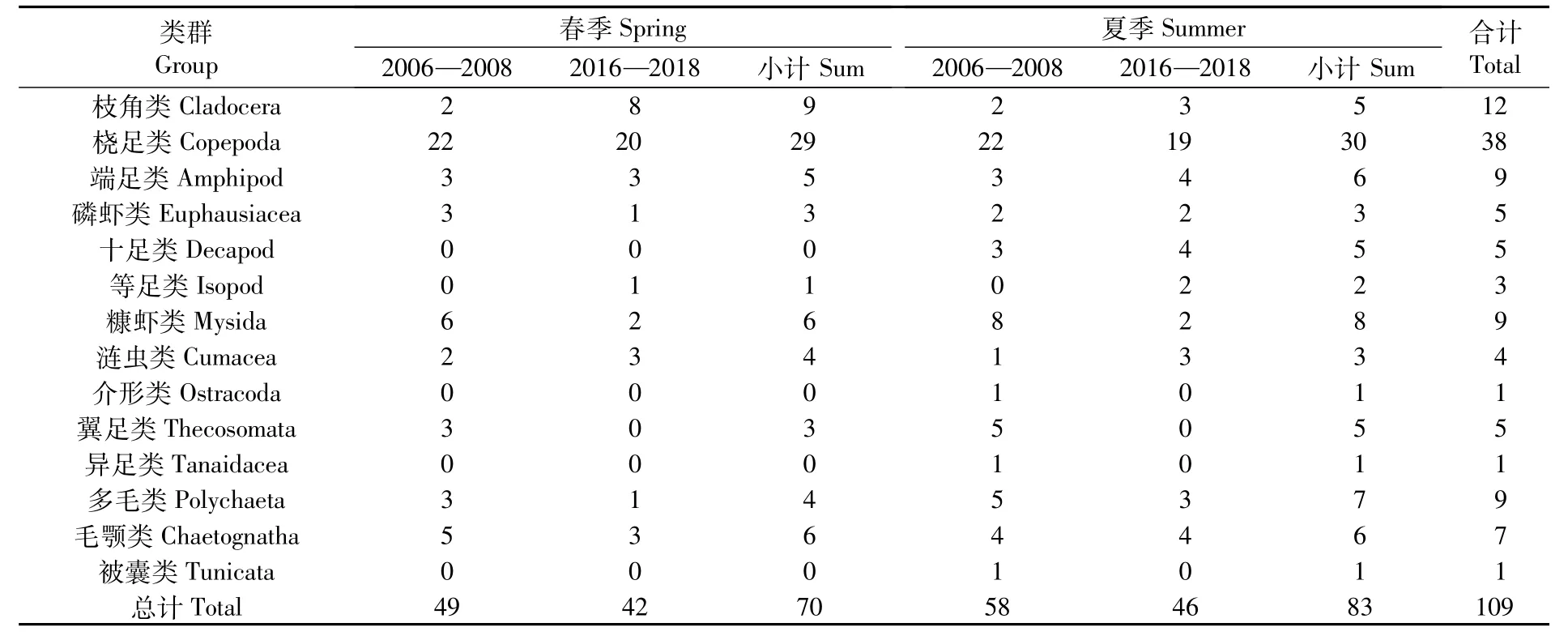
表1 长江河口浮游动物不同类群物种数Tab.1 Species of different groups of zooplankton in the Yangtze Estuary
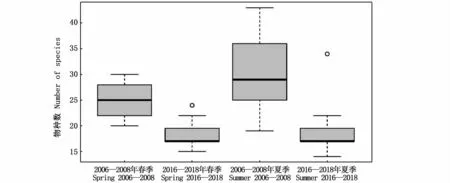
图2 长江河口各调查站物种数Fig.2 Number of species at each survey station in the Yangtze Estuary
2.3 浮游动物优势种
优势度Y≥0.02的优势种(表2)共计13种,分属3个类群,包括桡足类10种,糠虾类2种,毛颚类1种。
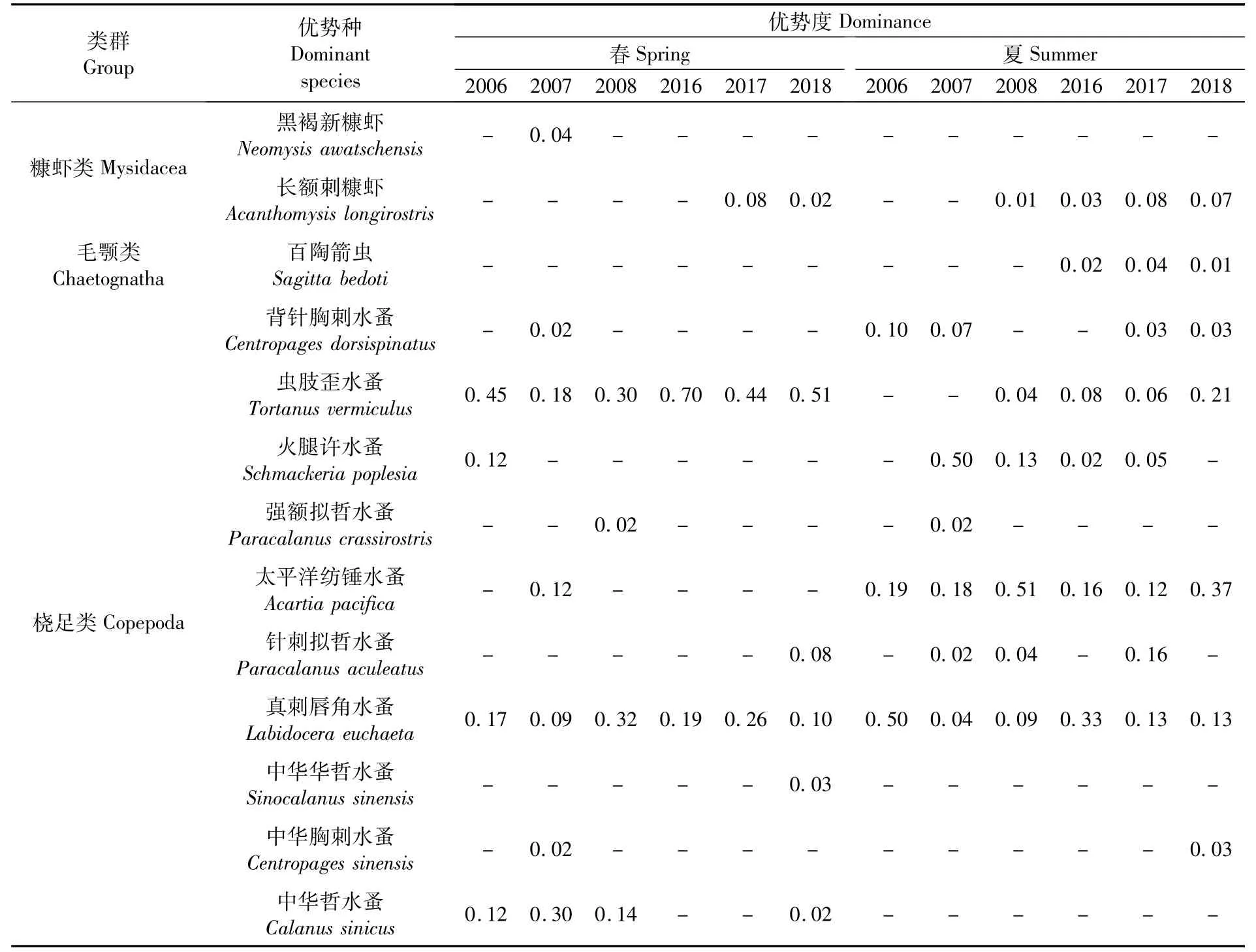
表2 各航次长江口浮游动物优势种Tab.2 Dom inant species of zoop lankton in the Yangtze Estuary in each voyage
在12次调查中,真刺唇角水蚤(Labidocera euchaeta)均为优势种,虫肢歪水蚤(Tortanus vermiculus)有10次为优势种,太平洋纺锤水蚤(Acartiapacifica)有7次为优势种。
春季主要优势种群比较稳定,虫肢歪水蚤、真刺唇角水蚤为春季主要优势种,两者在6个航次中均为优势种,其中虫肢歪水蚤有4次为第一优势种、2次为第二优势种,真刺唇角水蚤有1次为第一优势种、4次为第二优势种。夏季主要优势种群亦比较稳定,主要优势种为太平洋纺锤水蚤、真刺唇角水蚤和火腿许水蚤(Schmackeria poplesia)。太平洋纺锤水蚤和真刺唇角水蚤在6个航次中均为优势种,其中,太平洋纺锤水蚤2次为第一优势种、3次为第二优势种,真刺唇角水蚤有2次为第一优势种;火腿许水蚤有4次为优势种,其中1次为第一优势种。
2.4 浮游动物丰度时空分布特征
2006—2008年春季航次丰度均值为333.87个·m-3,夏季为562.75个·m-3;2016—2018年春季航次丰度均值为119.30个·m-3,夏季为208.40个·m-3。浮游动物丰度呈夏季高于春季及2016—2018年春、夏季丰度均低于2006—2008年同期的特点。
春季,两个调查阶段中浮游动物丰度均呈现沿岸略高、外海略低的特点(图3-A,C),丰度高值区均分布在北导堤外侧(3号站)及杭州湾湾口(7号站)附近海域。Kruskal-Wallis检验结果显示,不同调查阶段间的丰度具有显著性差异(P<0.05);但在每一调查阶段中,相邻3个航次间的浮游动物丰度无显著性差异(P>0.05)。

图3 长江口浮游动物丰度的分布(单位:个·m-3)Fig.3 Abundance distribution of zooplankton in the Yangtze Estuary
夏季,与春季相比,浮游动物丰度高值区向外海移动(图3-B,D),高值区多分布于崎岖列岛-鸡骨礁连线邻近海域。Kruskal-Wallis检验结果显示,不同调查阶段间的丰度具有显著性差异(P<0.05),2016—2018年相邻3个航次间的浮游动物丰度间有显著性差异(P<0.05),但2006—2008年相邻3个航次间的浮游动物丰度间无显著性差异(P>0.05)。
2.5 浮游动物群落聚类及nMDS分析
分别对2006—2008年春、夏季和2016—2018年春、夏季浮游动物丰度数据进行聚类和nMDS分析,结果见图4和图5。nMDS分析的stress值均低于0.2,说明聚类结果有一定的可信度[27-28]。

图4 长江口不同季节浮游动物群落聚类Fig.4 Clustering of zooplankton community in different seasons in the Yangtze Estuary
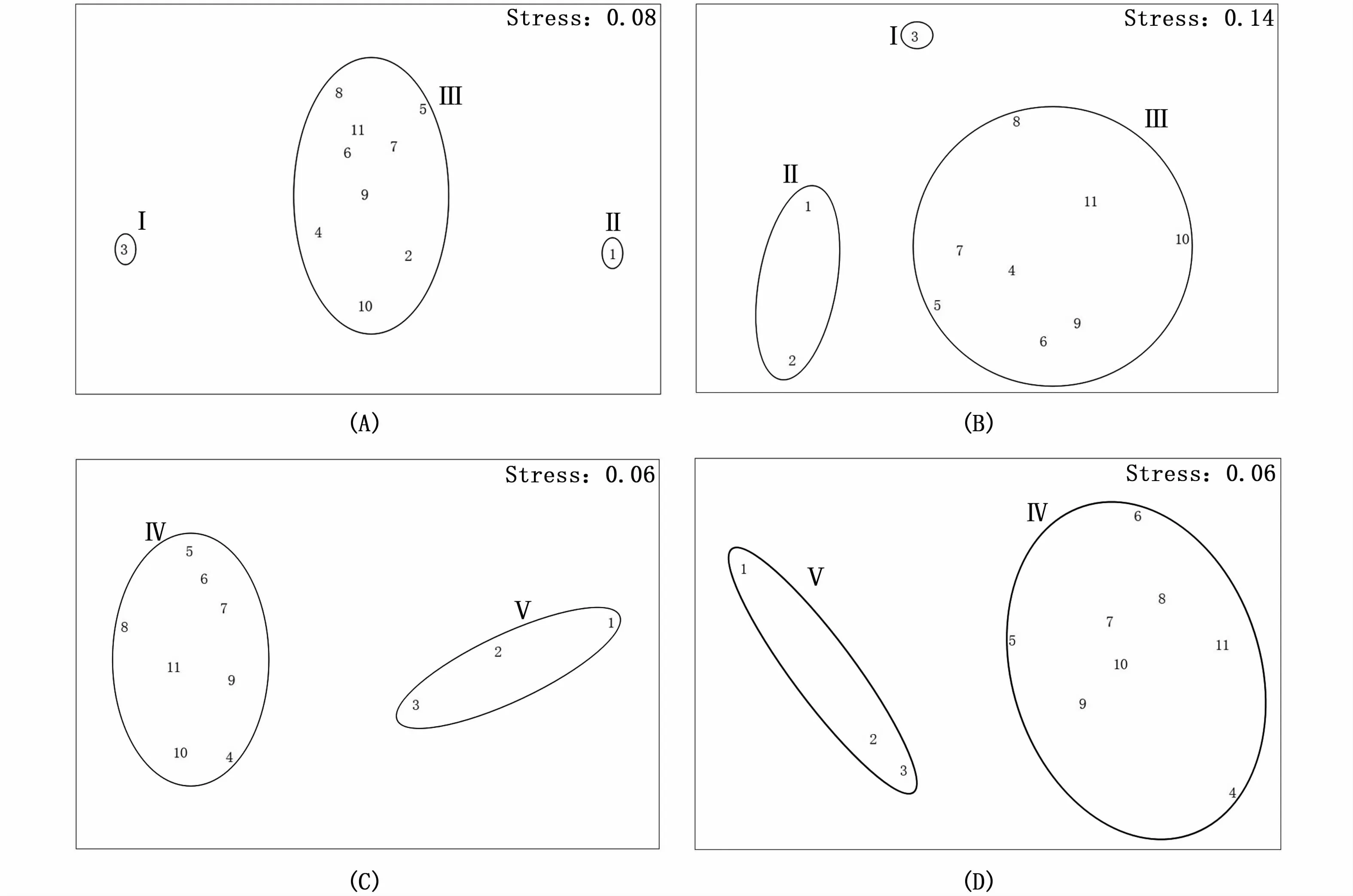
图5 长江口不同季节浮游动物群聚排序Fig.5 O rdination p lots of zoop lankton community in different seasons in the Yangtze Estuary
2006—2008年春、夏季浮游动物都可划分为3个类群:类群I仅包含3号站,为河口近海水域站位,主要受近海水团影响,属近海群落;类群II为河口水域站位,其中春季为河口口门区1号站位所形成的浮游动物群落,夏季则为河口口门区1号和九段沙附近2号站位形成的浮游动物群落,该区为最大浑浊带的核心区域,属长江口口门水域群落;类群III(共有站位有4、5、6、7、8、9、10、11)主要为崎岖列岛-鸡骨礁连线附近站位及杭州湾口各站,主要受枯季长江径流与口外近海水团的交错影响,属咸淡水交错水域群落。
2016—2018年春、夏季浮游动物可划分为相同的2个类群:类群IV(站位有4、5、6、7、8、9、10、11)主要为崎岖列岛-鸡骨礁连线附近站位及杭州湾口各站,主要受夏季长江口冲淡水与外海水团的交错影响,属咸淡水交错水域群落;类群V(站位有1、2、3)主要为河口最大浑浊带水域,属夏季长江冲淡水控制区群落。
2.6 不同时空浮游动物群落相似性分析
聚类和nMDS分析结果表明,类群I、II和V在地理位置上接近,均为近河口浮游动物群落,可概括为同一组群,命名为近河口组群。类群III和IV位于河口口门南侧的杭州湾、崎岖列岛-鸡骨礁连线水域,受咸淡水控制,可概括为同一组群,命名为河口南侧组群。
ANOSIM相似性分析检验结果如表3所示,同一调查阶段同一季节的不同聚类组群间均具有显著性差异(P<0.05)。春季,虫肢歪水蚤、真刺唇角水蚤等是导致浮游动物群落聚群组内相似和组间分异的关键物种。夏季,太平洋纺锤水蚤、火腿许水蚤和真刺唇角水蚤是导致浮游动物群落聚群组内相似和组间分异的关键物种。
ANOSIM相似性分析结果如表4和表5所示,同一组群在同一季节不同调查阶段间均具有显著性差异(P<0.05),在同一调查阶段不同季节间亦具有显著性差异(P<0.05)。春季,导致不同调查阶段间浮游动物群落分异的关键物种有虫肢歪水蚤、真刺唇角水蚤和中华哲水蚤,但其丰度在2016—2018年均显著降低;夏季,导致不同调查阶段间浮游动物群落分异的关键物种有太平洋纺锤水蚤、火腿许水蚤和真刺唇角水蚤,其丰度在2016—2018年亦显著降低。季节差异的相似性分析结果表明,在两个调查阶段中,虫肢歪水蚤、太平洋纺锤水蚤及真刺唇角水蚤的丰度变化均是导致浮游动物丰度季节差异的最主要原因。
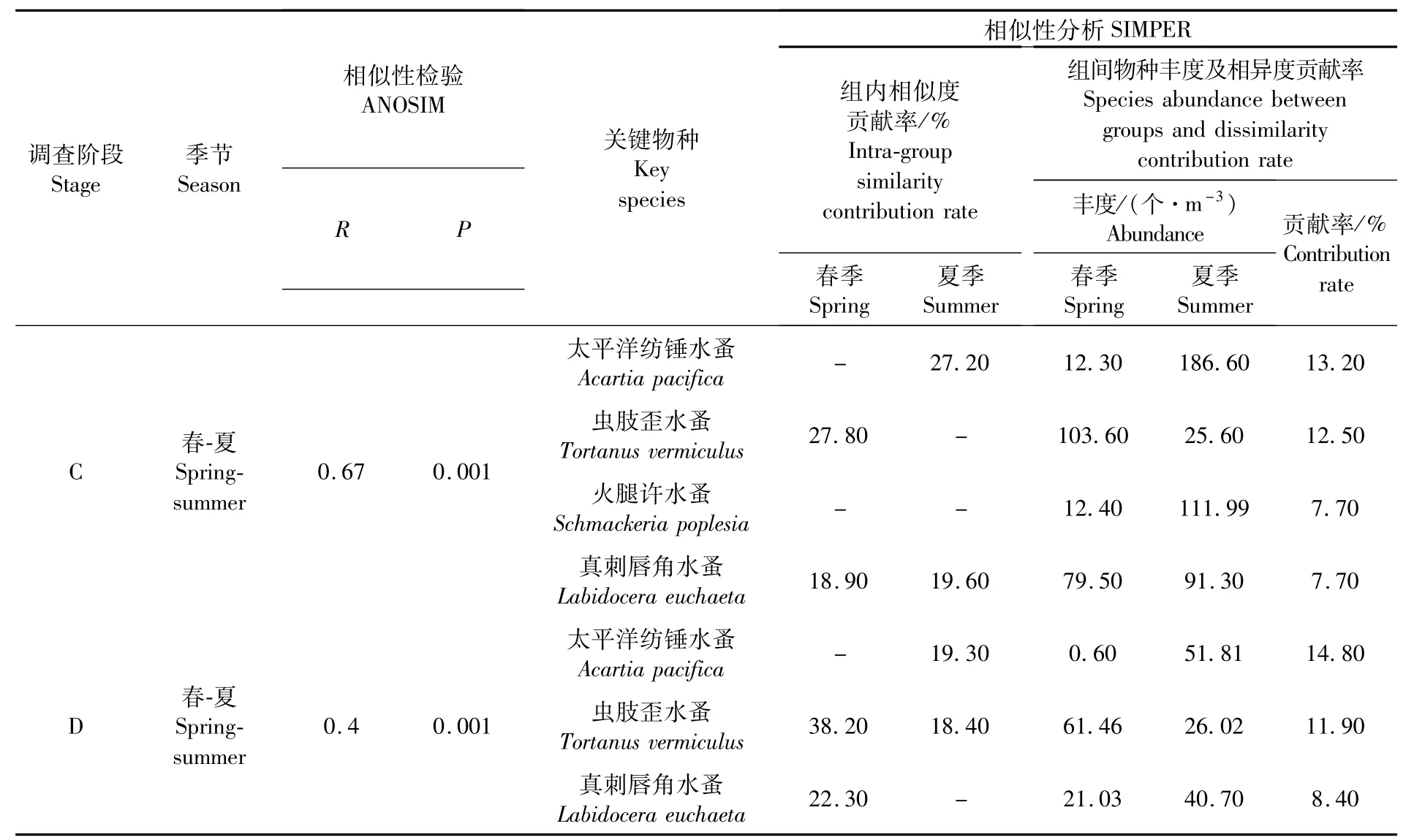
表5 不同季节相似性分析检验和相似性分析Tab.5 Sim ilarity analysis test and sim ilarity analysis in different seasons
3 讨论
3.1 浮游动物种类组成以及优势种的时空变化
分析比较不同调查阶段浮游动物的种类组成,可以发现与2006—2008年相比,2016—2018年浮游动物种类数呈减少趋势,这与杨颖等[29]在长江口的数据趋势相似,桡足类种类数比2006—2008年减少。浮游动物种类的变动与河口水沙条件的变化有很大关系[30],从大通站径流数据看,2016—2018年3年平均径流量比2006—2008年增加了21.8%,径流量的增加可能导致了强卷螺 (Agadina syimpsoni)、长轴螺 (Peraclis reticulata)等暖水性浮游翼足类在调查区的消失。
在两个调查阶段均观测到物种数量具有季节差异,夏季物种数多于春季,主要是由于夏季处于丰水期,长江冲淡水以及外海暖流的势力较春季均有所增强,不但为整个调查水域引入了更多的外海暖水种,而且不同水团间的剧烈交汇,形成了更为复杂的水域环境,可容纳更多种的生物类型生存,因此浮游动物类群变化范围被进一步扩大[10,31]。
在本研究鉴定的优势种中,真刺唇角水蚤、虫肢歪水蚤和太平洋纺锤水蚤广泛分布于调查海域各站,其中虫肢歪水蚤丰度高值通常出现在春季,真刺唇角水蚤在春夏季均有较高的丰度,太平洋纺锤水蚤在夏季有较高丰度,这些物种均为典型河口、沿岸低盐物种[23],可适应长江口剧烈变化的温盐环境。值得注意的是中华哲水蚤,其属近海广温广盐种,在2006—2008年春季航次的近河口海域大量出现(表2,表3),该海域受长江径流和海洋潮汐的影响,该种丰度与温盐呈显著正相关关系[10];当2016—2018年长江径流量相对增加时,该海域受低盐水控制,中华哲水蚤不再成为优势物种。
上述分析结果表明,真刺唇角水蚤、虫肢歪水蚤和太平洋纺锤水蚤等河口沿岸低盐种为长江河口区主要优势物种,其丰度会受到水温、盐度等环境因子的综合影响,某些物种如中华哲水蚤的分布会受到长江径流和海洋潮汐相互作用强度以及适温适盐性的影响,当长江入海流量减少时,近海广盐物种有可能进入河口区成为优势物种。
3.2 浮游动物丰度的时空变化特征
春季,虫肢歪水蚤和真刺唇角水蚤等优势种在长江口沿岸海域大量聚集,导致其在该区域的丰度值偏高,使浮游动物丰度均值呈现沿岸高外海略低的特点,这与之前的研究结果相似[31-32]。夏季,受长江冲淡水势力增大影响,丰度高值区相比春季向外海移动,集中在咸淡水交错水域。夏季浮游动物群落构成受到高温高盐的台湾暖流与长江冲淡水的影响,沿岸流带来的太平洋纺锤水蚤、火腿许水蚤等适应河口低盐生境的优势种大量繁殖。
浮游动物丰度均值呈现出夏季高春季低的现象,主要是由于夏季水温较高,咸淡水交错区营养物质丰富,浮游植物数量充足,有利于浮游动物的生长繁殖,导致夏季相比春季丰度增加[33]。浮游动物丰度在2016—2018年春季和夏季均比2006—2008年同季节显著下降,从相似性分析结果看,优势物种丰度的降低是差异的主要原因,推测与长江口近年来的生态环境变化密切相关[34],2016—2018年长江径流量的增加使得调查区域的盐度相比2006—2008年时低,限制了真刺唇角水蚤、虫肢歪水蚤、太平洋纺锤水蚤、中华哲水蚤等在本区域的大量繁殖。由此可见,水团的季节波动、外海高温高盐水和长江冲淡水交汇的位置以及潮汐的涨落等因素均有可能影响长江口浮游动物丰度空间分布[35]。
3.3 浮游动物群落结构的时空变化特征
基于较长时间序列的浮游动物丰度数据、群落聚类和nMDS分析,可以发现,不论2006—2008年还是2016—2018年,春、夏季的浮游动物群落空间分布趋势基本相同,聚类类群位置相对稳定。浮游动物类群I、II和V属于近河口区域群落,类群III和IV在长江冲淡水和咸水相互作用较强的地区,受咸淡水控制,属于河口南侧组群。本调查海域盐度在春、夏季均呈现长江口口门和近岸向外海扩散增高的趋势,因此两个区域水盐条件有一定差别,盐度分布是决定长江口浮游动物群落划分及其空间分布的主要决定性因子[36]。
浮游动物群落相似性分析结果表明,在长江口及邻近咸淡水交错区,春季浮游动物群落可归纳为虫肢歪水蚤-真刺唇角水蚤群落,夏季可归纳为太平洋纺锤水蚤-真刺唇角水蚤群落,在长江径流影响力减弱的年份,中华哲水蚤等广盐种有可能随高盐水进入调查区域成为优势物种。
