西北太平洋公海日本鲭生物学特征研究
2023-09-14赵国庆陈俊霖唐峰华陈云云贺建文
赵国庆,陈俊霖,2,张 衡,唐峰华,陈云云,贺建文
(1.中国水产科学研究院东海水产研究所,农业农村部东海与远洋渔业资源开发与利用重点实验室,上海 200090;2.大连海洋大学船舶与工程学院,辽宁大连 116000;3.中国水产舟山海洋渔业有限公司,浙江舟山 316101)
日本鲭(Scomberjaponicus)为大洋暖水性中上层洄游性鱼类,广泛分布于太平洋、印度洋和大西洋大陆架海域,资源量大且食用价值高[1-3],是中国近海和远洋重要捕捞对象,主要被围网和拖网所捕获[4]。数十年来,随着捕捞技术的进步、捕捞力量的增加以及人类对水产品需求量的增大,人类对近海渔业资源的开发规模越来越大,渔业可持续性水平发生了显著变化[5]。研究发现,世界范围内,超过90%的近海生物资源被过度开发[6]。日本鲭作为中国近海的主要捕捞对象之一,自20世纪80年代即表现出渔获个体小型化、低龄化的特点,90年代即出现过度捕捞现象[1,7],因此,加强对西北太平洋公海日本鲭资源的探索和开发对保护近海资源有一定的积极作用。
西北太平洋隶属于联合国粮农组织(Food and Agriculture Organization,FAO)61区,该区域具有十分重要的渔业地位,不仅是FAO统计区中的高产区之一,且区域内的国家如中国、日本和韩国等都具有悠久的渔业捕捞和渔业消费历史[8]。近年来,我国逐渐加强对西北太平洋公海渔业资源的开发力度,并积极参加国际上对该海域的渔业管理工作。2012年2月24日,我国作为缔约方之一签署了北太平洋公海渔业资源养护管理公约(Convention on the Conservation and Management of High Seas Fisheries Resources in the North Pacific Ocean),其后各缔约方成立了北太平洋渔业委员会(North Pacific Fisheries Commission,NPFC),开始对北太平洋渔业进行科学管理,以期实现该海域渔业资源的可持续利用[9]。日本鲭作为西北太平洋重要捕捞品种之一,是NPFC的重点管理对象。据NPFC统计,该海域日本鲭2021年的产量超过50万t,日本捕捞产量占比63.5%,而我国仅占20.2%,具有巨大的发展空间。
目前,我国对于西北太平洋公海日本鲭的研究内容涉及渔场变动[10-11]、资源评估[12]、单位捕捞努力量渔获量标准化[13]、渔获组成[14]、摄食生态[15]等方面。生物学特征是进行诸多深入研究的基础,目前该海域的相关研究仍有待完善和补充[16-17],基于此,本文以2016—2021年西北太平洋公海捕捞的日本鲭样品为对象,对其生物学特征进行了研究,以期丰富西北太平洋公海日本鲭生物学内容,为进一步研究其资源评估、开发利用与科学管理提供基础资料。
1 材料与方法
1.1 数据来源及渔场区域
本文所用日本鲭样品来源于西北太平洋公海生产渔业企业,现场随机取样并冷冻保存,记录采样时间及经纬度,样品采集时间为2016—2021年,共4 417尾(表1),作业区域分布范围为35°~43°N、150°~162°E(图1),作业方式为灯光围网捕捞。由于气候原因及日本鲭的生长发育情况,我国在西北太平洋公海的主要作业时间集中于每年4—12月,季节划分为春季(4—5月)、夏季(6—8月)、秋季(9—10月)、冬季(11—12月)。
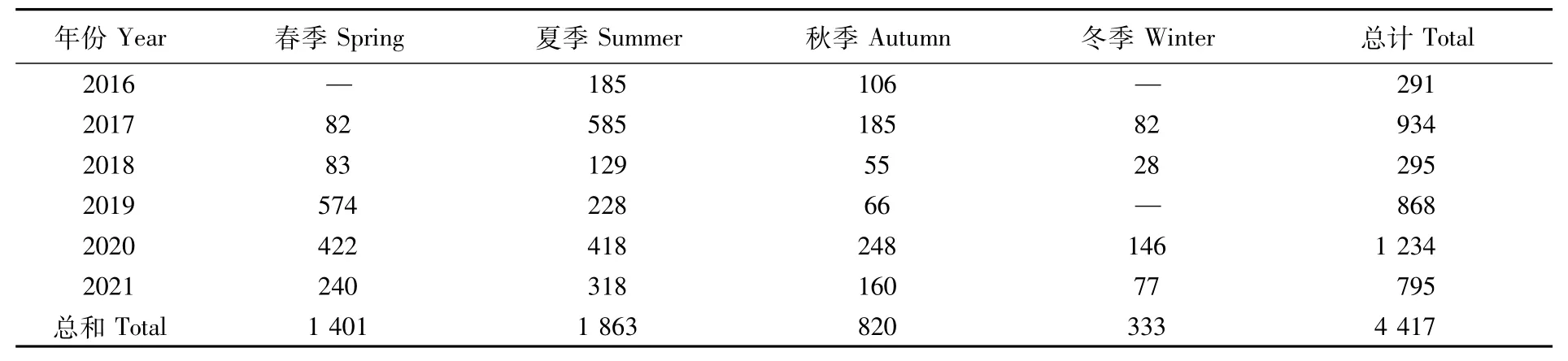
表1 西北太平洋公海日本鲭采样信息Tab.1 Sampling information of Scomber japonicus in the high seas of the Northwest Pacific(尾)

图1 西北太平洋公海日本鲭采样区域Fig.1 Sampling area of Scomber japonicus in the high seas of the Northwest Pacific
1.2 生物学测定
依据《海洋调查规范》[18]在实验室对样品进行生物学测量,测量指标包括性别、性腺成熟度、摄食强度、叉长(FL)和体质量(W),叉长与体质量分别精确到1 mm和0.1 g。性腺分为6期,分别用Ⅰ~Ⅵ来表示,摄食强度分为6级,用0~5表示,分期和分级标准参照殷名称[19]。在实验室进行生物学测量时,对所有样品的FL和W进行测量,单次样品不超过30尾时,全部解剖并对其性别、性腺成熟度和摄食强度进行辨认并记录,超过30尾时,随机抽取30尾进行解剖,另外,2016年未鉴定样品性别。
1.3 数据处理
1.3.1 叉长、体质量分布
利用频度分布法将体长和体质量进行分组并统计分析,体长组距10 mm,体质量组距20 g。分别利用K-S检验和单因素方差分析(one-way ANOVA)检测日本鲭雌雄群体叉长分布是否存在统计学差异及显著性。
1.3.2 叉长-体质量关系
利用幂函数拟合日本鲭叉长-体质量关系,其公式表示为:
式中,W为体质量(g),FL为叉长(mm);a、b为常数,分别代表生长条件因子和异速生长因子。b值通常在2.5~4.0之间,如果鱼类等速生长,则b=3,或接近3;b>3和b<3分别代表正异速生长和负异速生长[20-21]。同时,利用协方差检验性别对叉长体质量关系的影响。
1.3.3 性比、性成熟度和初次性成熟叉长
分别统计各年份日本鲭雌雄个体数量以及性腺成熟度,利用卡方检验分析判断雌雄比例是否符合1∶1。利用多因变量方差检验判别叉长和体质量与成熟度等级的显著性关系。一般将种群中50%的个体达到性成熟时的叉长作为该种群初次性成熟叉长,本文即将此时的叉长作为日本鲭的初次性成熟叉长[22-23],计算公式如下:
式中:Pi为日本鲭雌性成熟个体占组内样本比例;Li为各雌性叉长组上限(mm),L50为50%个体初次性成熟叉长(mm),c为模型参数。
1.3.4 摄食等级和肥满度
统计分析各年份日本鲭的摄食等级,利用多因变量方差检验判别叉长和体质量与摄食等级的显著性关系。采用Fulton状态指数计算日本鲭个体肥满度及平均肥满度,肥满度计算公式如下:
式中:K为肥满度系数,W为体质量(g),FL为叉长(cm)。采用单因素方差分析验证叉长组与肥满度的差异显著性。
本文数据分析采用Microsoft Excel 2019和IBM SPSS Statistic 19.0,图表绘制使用Origin 2018。
2 结果与分析
2.1 叉长、体质量组成
日本鲭叉长和体质量分布如图2所示。总体上,4 417尾日本鲭叉长范围为102~400 mm,分布呈双峰型,平均叉长236.50 mm,优势叉长组190~230 mm和270~300 mm,占比51.63%;除2019年、2021年和夏季外,其他年份或者季节,其叉长均表现出明显的双峰分布。总体上,体质量分布呈双峰型,第二个峰值低于第一个峰值,体质量范围为9.6~898.8 g,平均体质量165.0 g,优势体质量组50~150 g,占比46.20%;各年份和各季节体质量的分布峰型具有一定差异性,但多数仍为双峰型分布。
不同年份和不同季节的叉长和体质量分布表现出一定差异性,体质量差异性强于叉长。按年份分析,2020年叉长和体质量差异性显著强于其他年份,2018年叉长和体质量均值最高,2016年最低。按季节分析,夏季的叉长和体质量差异性强于其他季节,冬季的均值最高,夏季最低(图3)。
显著性分析结果显示,日本鲭雌雄群体叉长分布不存在统计学差异(P>0.05),而不同年份和不同季节叉长分布均差异极显著(P<0.01)。体质量分布与年份之间存在极显著差异(P<0.01),多重比较分析显示2019年与2020年差异不显著(P>0.05),其他年份之间均存在显著性差异(P<0.05)。
2.2 叉长-体质量关系
采用幂指数回归对日本鲭叉长和体质量进行拟合得到曲线,如图4所示,幂指数关系式为:
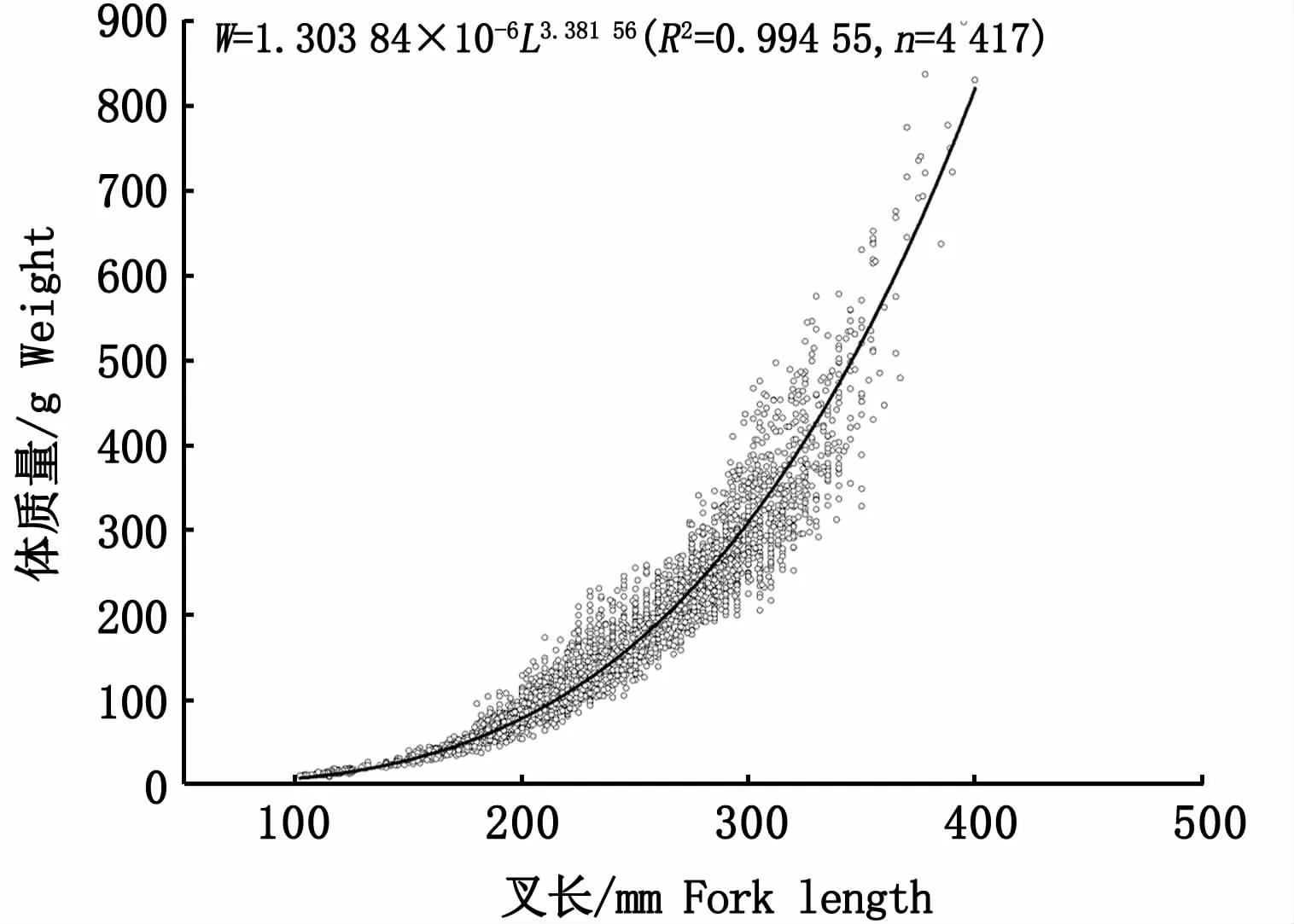
图4 西北太平洋公海日本鲭叉长-体质量关系Fig.4 Relationship between fork length and weight of Scomber japonicus in the high seas of the Northwest Pacific
按不同年份和不同季节对日本鲭的叉长-体质量关系进行分析,方程参数和拟合曲线分别如表2和图5所示。按年份分析,a值范围为7.10×10-7(2016年)~2.56×10-4(2021年),b值范围为2.44(2021年)~3.49(2016年),2021年变异程度强于其他年份。按季节分析,a值范围为1.09×10-6(冬季)~1.17×10-5(夏季),b值范围为2.99(夏季)~3.42(冬季)。年间和季节间a值的变化幅度分别为2.55×10-4、1.06×10-5,表明年间日本鲭叉长-体质量关系的变化强于季节。协方差分析(ANOVA)表明,性别对叉长-体质量关系影响不显著(P>0.05)。
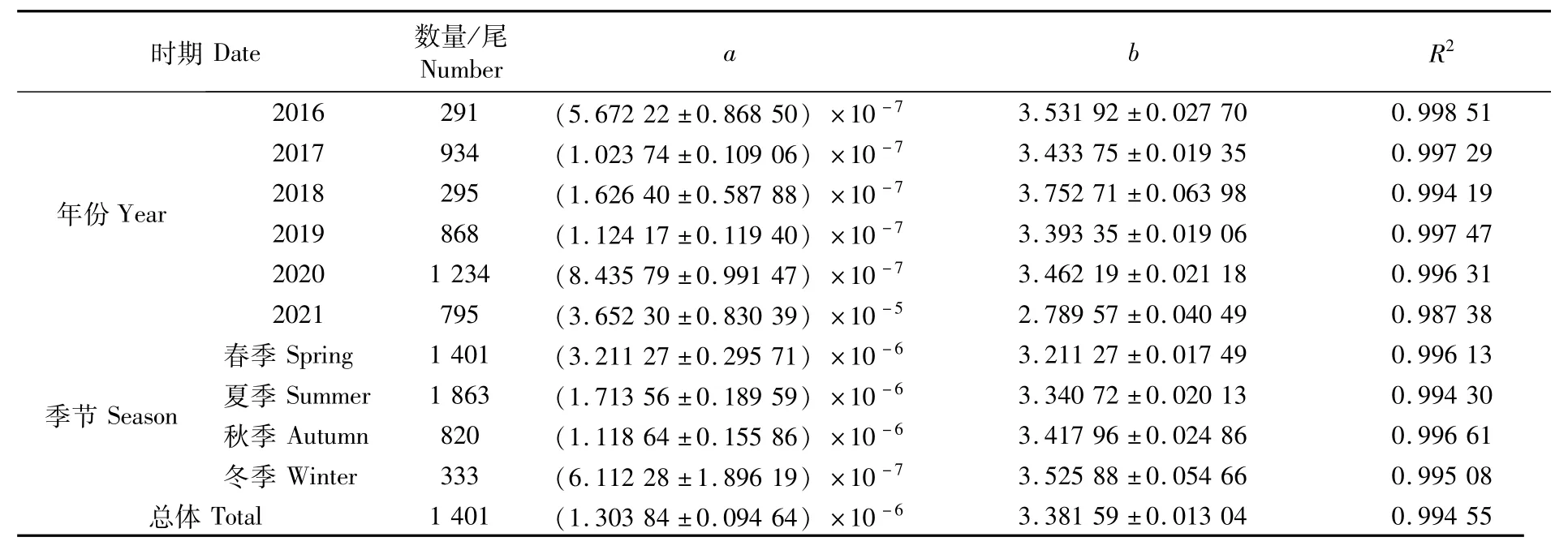
表2 西北太平洋公海日本鲭叉长-体质量幂函数参数Tab.2 Fork length-weight power function parameters of Scomber japonicus in the high seas of the Northwest Pacific

图5 西北太平洋公海日本鲭各年份和各季节叉长-体质量关系Fig.5 Fork length-weight relationship variations by years,seasons for Scomber japonicus in the high seas of the Northwest Pacific
2.3 性别分析
2.3.1 性比与性成熟度
共鉴定2 317尾日本鲭的性别和性腺成熟度,其中雌性1 472尾,雄性845尾,雌雄比为1.74∶1。卡方检验显示,雌雄比与1∶1差异显著(P<0.05)。
按照不同年份和不同季节进行分析,性成熟度均以Ⅱ期占比最高,其次为Ⅲ期,Ⅴ期和Ⅵ期群体均占比较少(图6);不同年份和不同季节各叉长组的雌雄比具有一定差异性,大个体中,雌性占比较高,当叉长超过390 mm时,雌性占比达到100%(图7);随叉长增大,达到性成熟的个体(Ⅲ和Ⅳ期)也随之增加,在性成熟个体中,Ⅴ期和Ⅵ期个体占比少(图8)。多因变量方差分析表明,叉长显著影响性腺成熟度(P<0.01)。
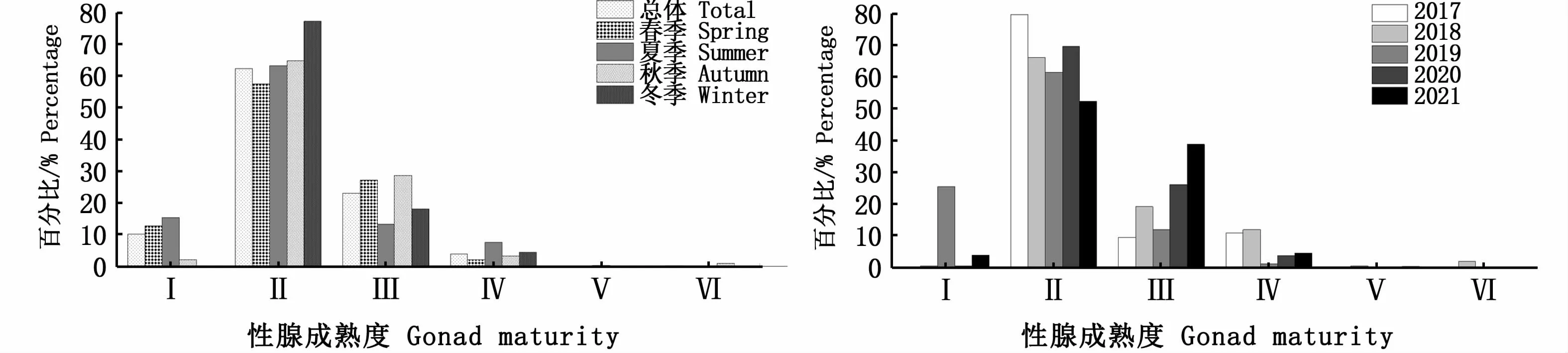
图6 西北太平洋公海日本鲭各年份和各季节性腺成熟度Fig.6 Gonad maturity of Scomber japonicus in the high seas of the Northwest Pacific by years and seasons

图7 西北太平洋公海日本鲭各年份和各季节性比与叉长的关系Fig.7 Relationship between sex ratio and fork length of Scomber japonicus in the high seas of the Northwest Pacific by years and seasons
2.3.2 初次性成熟叉长
根据不同叉长组中性腺成熟个体的百分比拟合日本鲭逻辑斯蒂曲线(图9),结果显示,雌性、雄性个体初次性成熟叉长分别为(298.08±4.32)mm、(287.82±4.26)mm,方程如下:
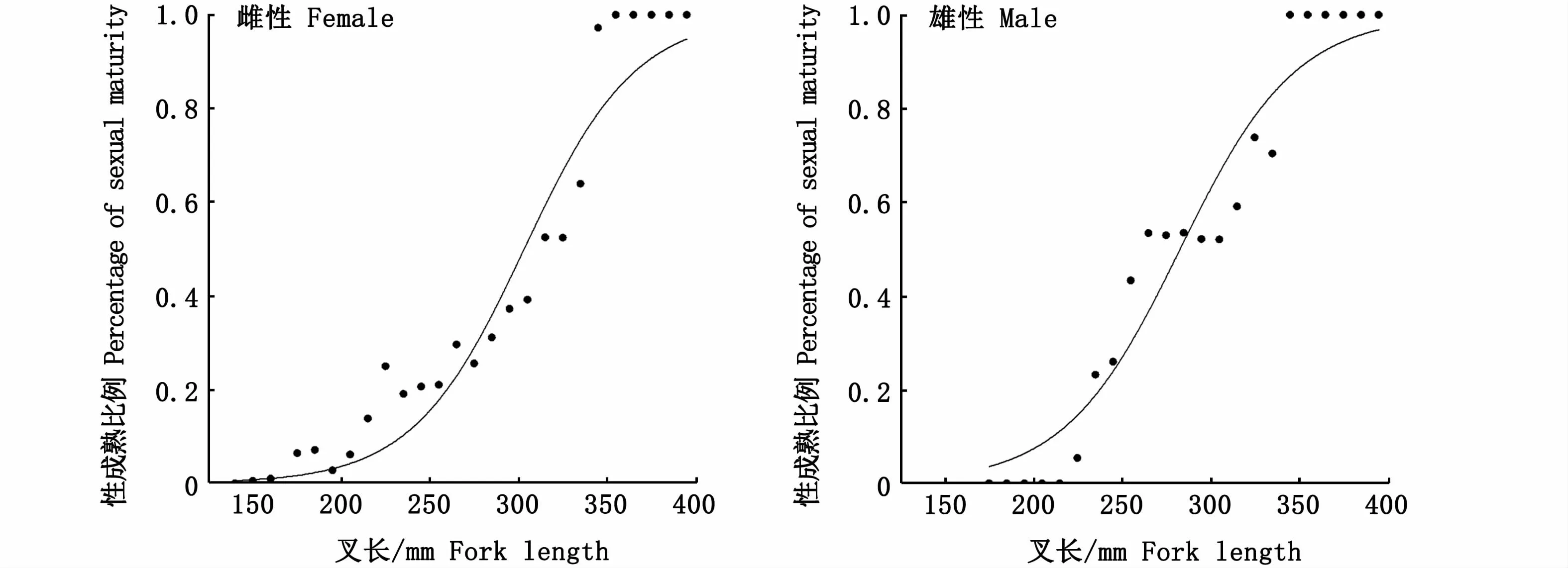
图9 西北太平洋公海日本鲭初次性成熟叉长Fig.9 Fork length at the first sexualmaturity of Scomber japonicus in the high seas of the Northwest Pacific
2.4 摄食等级和肥满度
2.4.1 摄食等级
总体上,摄食等级以1~3级为主,占比59.05%,5级占比最少,为0.52%;按不同季节分析,3~5级各自的占比随春夏秋冬基本上逐渐增加,年间变化较为随机,5级个体仅在2021年出现(图10)。各叉长组所对应的摄食强度组成如图11所示,空胃以及5级占比均较少,空胃在小于140 mm的小个体中占比高,2级占比最高,为33.55%。摄食强度与叉长显著相关(P<0.05),与性别的相关性关系不显著(P>0.05)。
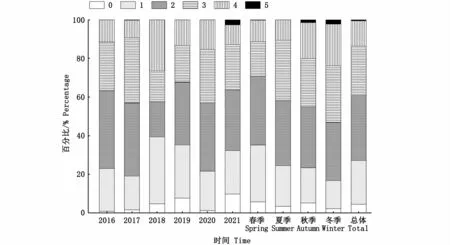
图10 西北太平洋公海日本鲭各年份和各季节摄食等级组成Fig.10 Feeding intensity of Scomber japonicus in the high seas of the Northwest Pacific by years and seasons

图11 西北太平洋公海日本鲭各年份和各季节不同叉长组与摄食强度百分比组成Fig.11 Percentage of feeding intensity in different fork length groups of Scomber japonicus in the high seas of the Northwest Pacific by years and seasons
2.4.2 肥满度
总体上,对所有个体分析结果显示,K值范围为0.64~2.45,均值1.30。年份和季节均显著影响其肥满度(P<0.05),年间K值变化范围为0.97(2019年)~1.14(2021年),季节间按照春夏秋冬依次增大,范围为0.97~1.19。叉长显著影响肥满度(P<0.05),随着叉长增加,日本鲭肥满度基本上均随之增大(图12)。
情况 3.2.1 C1中的集合都是Y中顶点色集合,4,5,6中至少有2种色同时包含在每个C(ui)中,不妨设4,C(ui), i=1,2,…,10,则C2中的集合都不是X中顶点色集合,且至多有3个不是Y中顶点色集合。
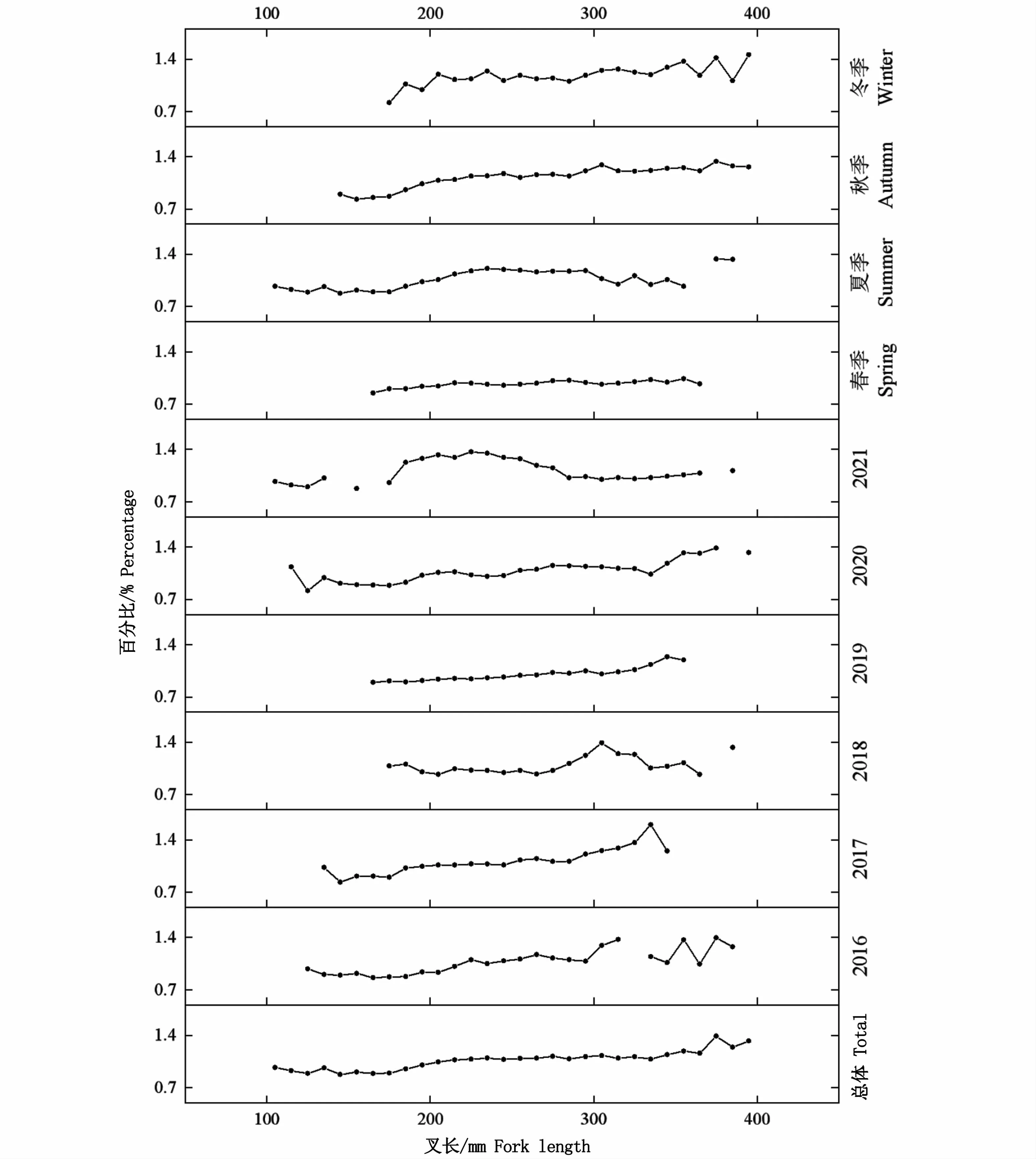
图12 西北太平洋公海日本鲭肥满度与叉长的关系Fig.12 Relationship between fork length and fatness index of Scomber japonicus in the high seas of the Northwest Pacific
3 讨论
3.1 叉长-体质量关系
体长和体质量是鱼类最基本的生物学特征,通常被换算为资源评估模型的基本参数用于渔业科学研究,也是鱼类个体低龄化、小型化最直观的表征[24],两者之间的函数关系一直被用于描述鱼类的生长特征。其中,幂函数公式W=aLb是用来描述鱼类生长最常用的关系式,式中a和b通过统计分析得出,分别代表条件因子和异速生长因子,是渔业资源评估的重要参数[25],能够反映鱼类生长的稳定性和适应性[26]。参数a代表种群生长的环境条件,易受水文、季节、栖息地、饵料等因素的影响[27-30];参数b反映鱼类生长状态,表示鱼类生长的不均匀性[30],这种不均匀性是由体质量(或体积)和体长的不均匀增长带来的,如果体长、体高和体宽为等速生长,比重不变,则b=3或接近3[20-21]。b值大小与鱼类生长发育状态密切相关,幼鱼的b值多小于3,呈强异速性生长,随鱼体生长发育,异速性减弱,发育趋于均匀,成鱼时b值接近或大于3,其生长状态可描述为:异速生长—等速生长—异速生长[27]。
本研究中,叉长-体质量关系式W=1.303 84×10-6L3.38156,b值大于3,表明西北太平洋公海日本鲭处于正向异速生长状态,即体质量增长速度快于叉长,能够较好育肥。受生长阶段、饵料水平、摄食效率、性腺发育程度和产卵频率等因素的影响[28],参数a和b的值也一直都处于变化中,本研究的参数a和b均处于表3所列举相关报道的a(6.55×10-7~1.66×10-5)和b(2.95~3.59)变化范围内。FROESE[28]认为,b值远小于或者远大于3(b<2.5或b>3.5)一般都不具代表性,可能是样品数量少或者长度范围小所致。据报道,日本鲭叉长-体质量关系中参数b的正常范围为2.7~3.7[4],而北太平洋日本鲭的b值大多时候均大于3(表3),表明该海域日本鲭的体质量增速快于叉长。与以往的记录相比较(表3),本研究中日本鲭叉长的范围更广(102~400 mm),时间跨度更长,样品量更大,研究所得的日本鲭叉长-体质量关系式应更为合理。因此,本研究结果对于西北太平洋公海日本鲭渔业资源的可持续利用和保护管理是一个十分有价值的基本参考。但是在日本鲭生物学的诸多研究中[1,4,16,31-35],人们很少捕获到小于100 mm的样品,假如小个体缺失,是否会对拟合的叉长-体质量关系产生影响?利用这些缺失小个体样品的叉长、体质量或者年龄作为参数的资源评估结果是否值得信任?这些问题尚需未来进一步研究解决。

表3 不同时期日本鲭叉长-体质量关系参数Tab.3 Parameters of relationship between fork length and weight of Scomber japonicus
在本研究中,叉长-体质量关系(图5)及参数a和b(表2)各年份间虽有差异,但是并无明显的变化规律,然而,其季节性变化特征十分明显。参数b随季节变换而变化,最小值和最大值分别为3.21(春季)和3.53(冬季)(表2),当个体叉长一定时,体质量由春季到冬季依次增大(图5),即体质量增速与叉长增速的比值随之逐渐增大。日本鲭具有产卵洄游、索饵洄游、越冬洄游等长距离洄游特性[36],这可能是导致参数b季节性变化的重要原因。西北太平洋公海日本鲭的产卵期为每年的3—6月[36],整个春季都存在繁殖个体。鱼类在繁殖期摄食强度降低[37],部分肌肉组织在性成熟过程中将会转变为性腺[28]。因此,鱼类体质量的增加在春季将会变缓甚至停止,b值降低。此外,渔场重心的季节性变化可能是影响b值变化的另外一个原因,有研究表明,西北太平洋公海日本鲭渔场重心具有明显的季节性变化特征[11]。
3.2 性比及性腺成熟度组成
鱼类的雌雄比受多种因素诸如个体大小、海域、年份、季节等因素制约[38]。在繁殖季初期,日本鲭雌雄个体数量基本相当,繁殖旺季雌雄比达到最大值,其后逐渐下降,产卵末期雌性个体明显少于雄性[39]。个体规格显著影响雌雄比,小个体雌雄比基本符合1∶1,大个体群体中雌性显著多于雄性[39],在鱼类中这种现象较为普遍,其他鱼种诸如东黄海的黄鮟鱇(Lophiuslitulon)[40]和蓝点马鲛(Scomberomorusniphonius)[41]也符合这个规律。日本鲭雌性个体在产卵期产卵数次,每次产卵数量为5万~9万粒,据推测,为了使自身体腔能够容纳更多数量的卵子,性成熟后雌性个体会保持更高的生长速率,以此来达到维持种群数量的目的[39]。本研究中,日本鲭雌雄比为1.74,庄之栋等[16]、戴澍蔚[17]得出的雌雄比也均大于1,表明该海域雌性个体数量多于雄性。在繁殖季节,雌性会通过多次产卵来达到补充群体丰度的目的,因此,据推测雌性个体多于雄性个体可能是一种维持种群数量的手段[42]。
鱼类性腺发育具有较强的季节性,春季是西北太平洋公海日本鲭的产卵期,因此春季性腺达到成熟的个体比例相对较高,但是笔者发现这种现象并不明显(图6)。西北太平洋公海日本鲭具有距离洄游特性,3—6月在日本大陆沿岸海域产卵,随后向西北进行索饵洄游[43-44],在生殖洄游过程中大部分产卵个体在日本近海被消耗,这可能是造成索饵场内性成熟度高的个体数量少的重要原因之一。本研究中,性成熟度Ⅱ期个体占绝对优势,Ⅳ期、Ⅴ期和Ⅵ期个体数量少,特别是Ⅴ期和Ⅵ期,占比仅为0.30%。庄之栋等[16]发现,2016年8—10月北太平洋公海日本鲭的性腺成熟度全部为Ⅰ~Ⅱ期。因此,西北太平洋公海海域内日本鲭鲜见繁殖行为。本研究中,叉长显著影响日本鲭性腺发育程度,随着叉长增加,Ⅰ期、Ⅱ期占比逐渐下降,Ⅲ期、Ⅳ期占比逐渐升高。
日本鲭初次性成熟时间取决于其叉长而并非年龄,性腺在其叉长达到一定值后即开始发育成熟[45]。日本鲭初次性成熟的叉长具有较大的时空差异[46-47],本研究发现,西北太平洋公海雌性个体初次性成熟叉长为(298.08±4.32)mm,与20世纪50年代到60年代我国近海结果接近[48-49](表4)。目前,近海海域内日本鲭性成熟提前及初次性成熟叉长减小已成普遍现象,但是笔者发现,西北太平洋公海日本鲭初次性成熟叉长与历史数据对比并没有减小趋势,可能显示该海域内日本鲭资源并未受到捕捞压力的显著影响。
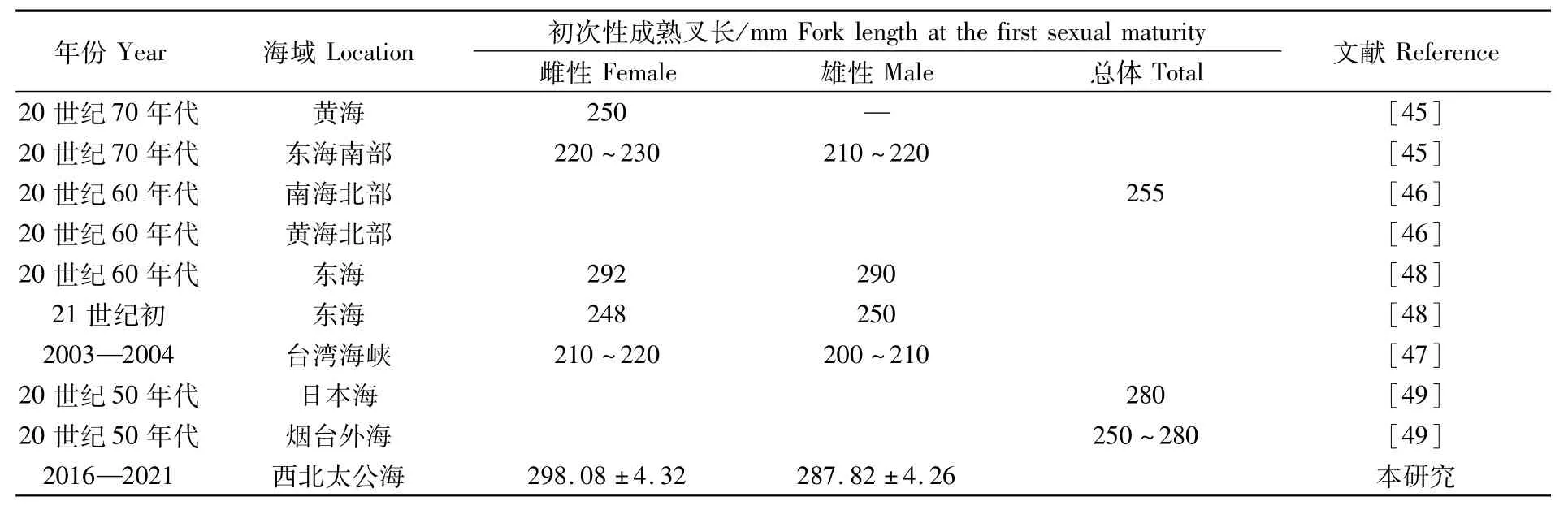
表4 日本鲭初次性成熟叉长Tab.4 Fork length at the first sexualmaturity of Scomber japonicus
3.3 摄食等级与肥满度
日本鲭的摄食饵料随个体增大而变化,种类先增加后减少,并由小个体逐渐向大个体转变[15]。这种现象符合“最佳摄食理论”,可以让摄食者得到最大的食物净值。在鱼类摄食中,摄食强度与叉长存在多种关系:正相关[50]、负相关[51]或没有显著差异[52]。日本鲭活动能力强,洄游路线长,其摄食等级和摄食种类具有明显的季节性变化。这种变化往往与水温有直接关系,水温对于鱼类的摄食具有重要影响,摄食强度随着水温降低而减小[53],温度降低时,一部分能量转为脂肪用来保持体温,此时摄食强度低[15]。但是在本研究中,笔者发现日本鲭的摄食强度从春季到冬季依次增强,空胃和1级的个体占比逐渐减少,其他摄食等级个体占比逐渐增加,这可能与日本鲭的洄游有关,本研究海域内日本鲭渔场重心在每年的8—9月由东北向温度较高的西南移动[11],说明日本鲭在此范围内洄游。且本研究显示日本鲭摄食强度以1~4级为主,空胃个体少,此前有研究指出,西北太平洋公海日本鲭的摄食等级以2~4级为主[16],本文结果与其相似,表明西北太平洋公海可以基本满足日本鲭的饵料需求。肥满度是用来表征鱼类肥瘦程度和生长情况的指标,可用来分析鱼类生长速度、成活率和总体增重程度,同时也可用来间接评价生活环境状况[54]。本研究发现,叉长显著影响日本鲭肥满度,随叉长增加肥满度逐渐增大,体质量增加快于叉长,表明日本鲭在该海域能够较好的育肥。结果表明,该海域能够为日本鲭提供良好的觅食及育肥条件,是其生长发育的优良渔场。
3.4 不足与展望
1)鉴于目前存在关于西北太平洋公海存在多种日本鲭种群的猜想,本研究仅初步鉴别了研究海域的样品种群,未能结合更多方法做出有效认定,这也是后续有待深入研究的问题。
2)本研究发现,大个体中雌性占比较高,未来可针对此现象进一步探讨原因。
鱼类的生物学特征是进行深入研究的基础,由于我国对西北太平洋公海捕捞的规范化管理起步较晚,同时科学调查存在成本高、采样困难等问题,该海域诸多鱼种的生物学特征及其诸如年龄与生长、生活史、种群划分等方面都有待深入探讨。在后续研究中,笔者将首要明晰西北太平洋公海日本鲭年龄结构组成,进一步地进行资源评估,并分析其生活史,对其生长发育、繁殖产卵、摄食、洄游等进行深入研究,为合理开发利用该海域日本鲭资源提供科学指导,并为我国参与NPFC的相关工作提供理论支撑,提高我国在NPFC中的话语权。
