塞拉利昂海域雨季深海声学散射层生物垂直迁移现象分析
2023-09-14屈泰春冯春雷李灵智范瑞良邵盛男张志超
屈泰春,冯春雷,,李灵智,,范瑞良,邵盛男,张志超
(1.中国水产科学研究院海洋捕捞工程技术研究中心,上海 200090;2.中国水产科学研究院东海水产研究所,农业农村部东海与远洋渔业资源开发利用重点实验室,上海 200090)
海洋水体,根据深度在垂向上可分为上层带(epipelagic zone,0~200 m)、中层带(mesopelagic zone,200~1 000 m)、深层带(bathypelagic zone,1 000~4 000 m)、深渊层带(abyssopelagic zone,4 000~6 000 m)和远洋海底带(hadopelagic zone,>6 000 m)[1]。伴随日出日落,中层带与上层带之间重复上演着地球上规模最大的生物昼夜垂直迁移运动(diel vertical migration,DVM)[2]。日落时分,海洋生物由约300~850 m的中层带上升至<200 m的上层带捕食,日出时分再下降到初始水层[3]。由于此生物层由声学探测设备发现,因此被称为深海声学散射层(deep scattering layers,DSL或DSLs)。
DSL最早于1942年在太平洋被发现[4]。随后的调查表明,世界各大洋均有DSL的分布[5-7],是中层带的一个普遍特征[8]。DSL生物由大洋中层鱼类(mesopelagic fishes),主要是灯笼鱼科(Myctophids)或圆罩鱼属(Cyclothone)鱼类,以及小鱿鱼、小虾和胶质浮游生物(gelatinous plankton)集群等组成[5]。中层带占据了约20%的海洋水体[9],其中很可能蕴藏着最大的海洋生物量[6]。DSL生物,特别是其中的游泳动物(micronekton)是食物网中连接浮游生物(zooplankton)和顶级捕食物种的关键纽带[5],是生物碳循环(biological carbon pump,BCP)的一个重要环节[6,10-11]。同时,DSL生物的活动规律和空间分布被认为与大尺度的环境条件有关[9]。因此,对DSL的探测可作为海洋相关学科研究和海洋环境监测的一种方法[4]。
塞拉利昂海域属于热带海域,位于东大西洋三大强烈涌升流区域之一的毛里塔尼亚(Mauritania)涌升流势力范围以南,以高初级生产力著称[12],其专属经济区(exclusive economic zone,EEZ)面积104 850 km2,渔业资源丰富[13]。1963—1974年间,由海洋学家RICHARD主持的WHOI(Woods Hole Oceanographic Institution)项目的主要目标为发现大西洋中层鱼类分布与DSL之间的关系[12],调查范围遍及整个大西洋以及部分与大西洋毗邻的北冰洋海域。该项调查共进行了1 040次中层拖网,但在塞拉利昂海域附近仅实施了一个网次拖网,调查强度极低,不足以了解该海域DSL的组成和时空分布规律。此后,鲜见针对该海域DSL的报道。塞拉利昂海域DSL的研究有助于对中东大西洋生物碳循环规律的认知和海洋环境的监测,也对该海域渔业资源形成和发展的相关研究具有重要意义。本文利用2021年5月于塞拉利昂海域中上层渔业资源与环境调查中获取的船载Simrad EK60科学渔探仪(38 kHz)声学数据,对该海域DSL生物的垂直迁移过程和迁移量进行分析研究,以期为了解该海域DSL组成和分布规律提供参考。
1 材料与方法
1.1 数据来源
原始声学数据于2021年5月1日—5月8日西非塞拉利昂中上层渔业资源与环境调查时采集(图1)。声学探测设备为船载Simrad EK60型分裂式波束科学渔探仪,于4月22日在调查海域(8°30′06″N、13°34′17″W)进行校准[14]。38 kHz声学换能器发射功率2 kW,数据采集最大水深1 km,声脉冲收发间隔1 s。
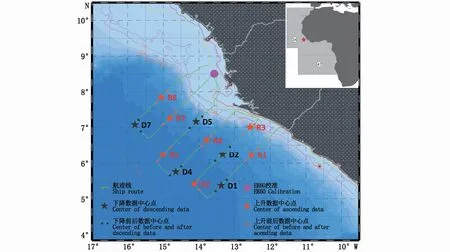
图1 调查海域及研究数据选取位置Fig.1 Survey area and locations of data used
选取走航中[4,15]底深大于800 m、垂直迁移前后1.5 h内的38 kHz声学数据进行分析,共有12段声学数据适用于垂直迁移现象的研究(表1)。但5月4日上升区域R4站位数据质量较差,未被采用;因此,共有6段上升和5段下降的可用数据。
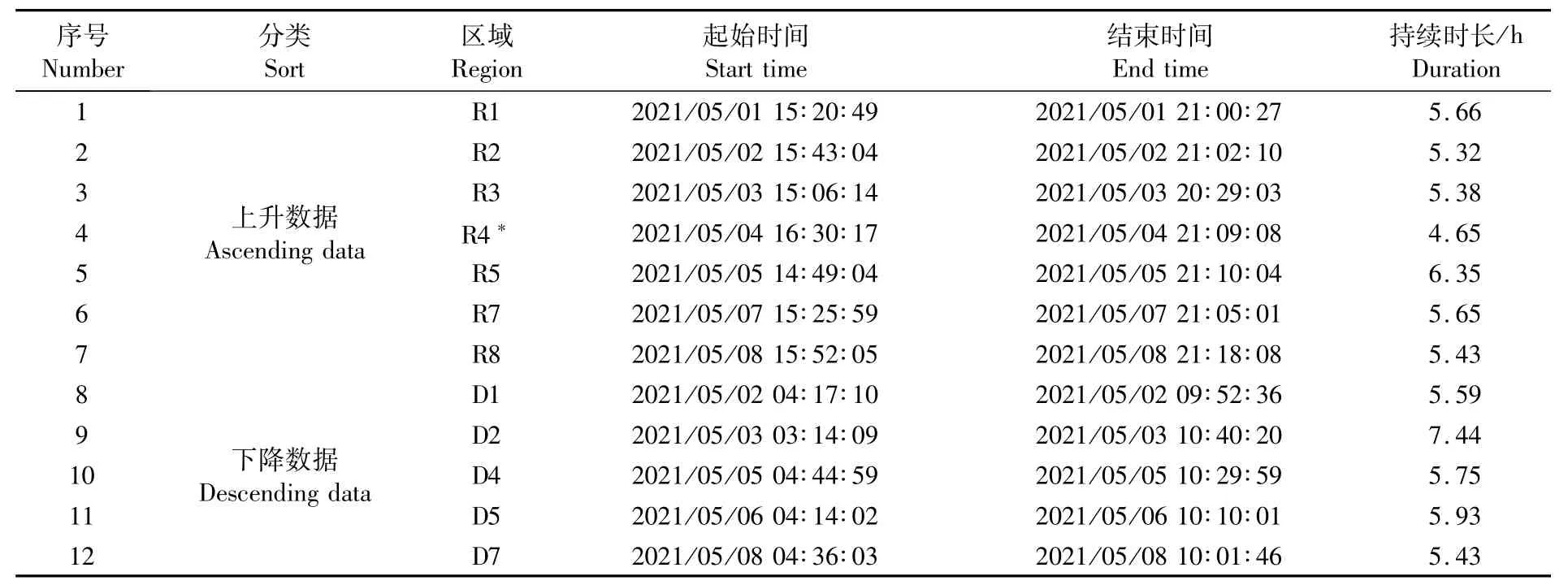
表1 适用于垂直迁移现象研究的原始声学数据时段Tab.1 Raw acoustic data suitable for the analysis of diel verticalm igrations
1.2 数据处理软件
Echoview®(V8.0)软件用于对声学数据进行处理,MATLAB(R2016a)用于对Echoview导出的数据做进一步处理,Surfer 16用于部分图形的制作。
1.3 数据处理
1.3.1 声学数据预处理
首先,提取、剔除原始声学数据的背景噪声(background noise,BN)[16-17],重采样间隔为20 Pings×5 m;之后手动剔除表层盲区、泡沫、海底回波及其他混响数据;最后,根据回波映像结合DSL生物的垂直迁移特征,将研究水体由上到下依次划分为浅水层(shallow layer,SL)、中间层(middle layer,ML)和深海声学散射水层(deep scattering layer,DSL)3个水层。
1.3.2 迁移速率的计算
采用净垂直速率(net vertical velocity,NVV)[18]表征DSL生物的迁移速率,即迁移速度在垂向上分量的绝对值,通过对迁移生物层的深度时间曲线求导获得。
首先,应用Echoview软件中可编辑线及相关位图运算模块,依据视觉辨别,粗略提取出迁移回波信号,回波信号分层的则分别提取。在提取出的各层回波信号范围内先后使用最大Sv拾取器(maximum Sv line pick)和平滑滤波器(smoothing filter)功能模块,即可获得各迁移生物层的原始深度时间曲线。
然后,应用MATLAB软件中的经验模态分解(empirical mode decomposition, EMD)工具箱[19-20]对原始深度时间曲线进行分解,对最后一个本征模函数(intrinsic mode function,IMF)求导得出迁移速率。当EMD最后一个本征模函数与原始深度时间曲线相差过大时,使用原始深度时间曲线求得迁移活动的相关统计数据。
1.3.3 迁移量的计算
由于较难确定迁徙生物的物种组成,所以无法给出直观的迁移生物资源量。但由于NASC(nautical area scattering coefficient)值与生物量呈线性关系[8],所以本文采用各水层生物迁移前、后1.5 h至0.5 h之间的1 h时间段内NASC值的变化代表对应水层生物量的变化。
2 结果与分析
2.1 声学数据预处理
依据声学数据预处理后的回波图(图2)和迁移生物分布规律,将水体划分为11~96 m的浅水层(SL)、96~277 m的中间层(ML)和277~563 m的深海声学散射层(DSL)(图3)。

图2 声学数据预处理前(a),后(b)对比图Fig.2 Contrast figure between before(a)and after(b)process of raw acoustic data
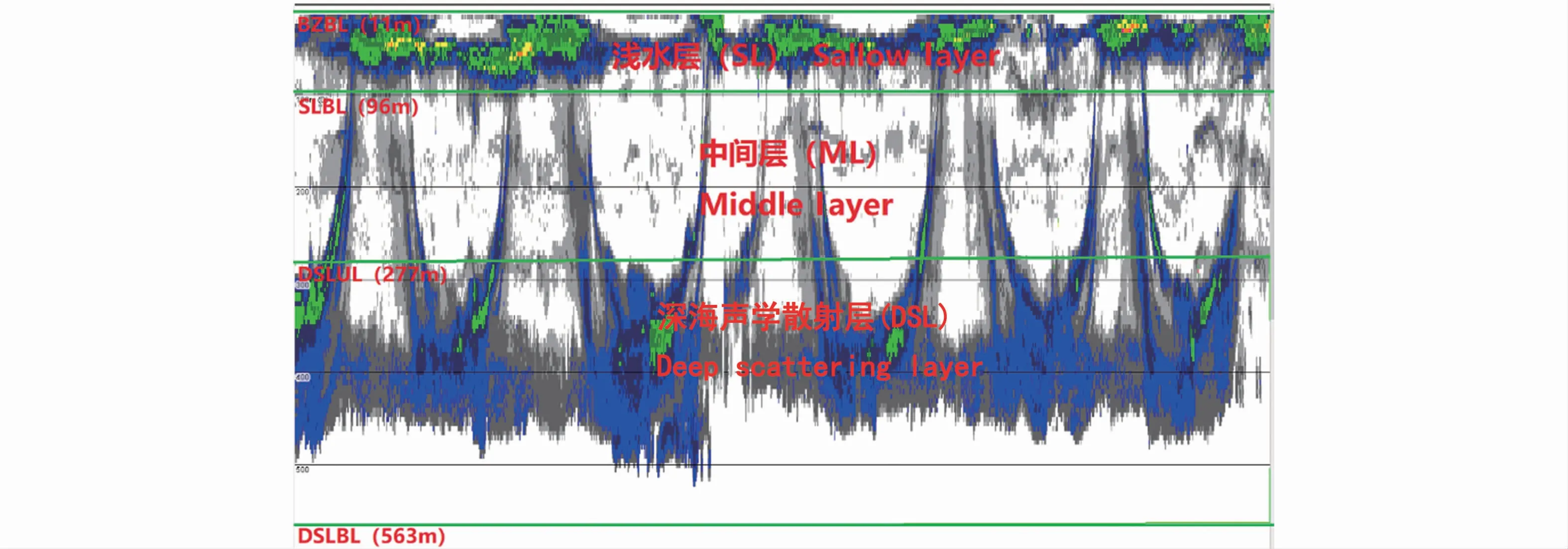
图3 水层和边界线的定义Fig.3 Definition of layers and their boundary lines
2.2 迁移速率
迁移生物日落时分上升,日出时分下降,且分层进行(图4)。各采样点生物分层数量和迁移规律存在差异。本文共获得38个生物分层,即38条深度时间曲线。6个上升区域曲线数量2~4条,共20条,平均每个区域3.3条;5个下降区域曲线数量2~5条,共18条,平均每个区域3.6条(表2)。

表2 各数据区域迁移曲线数量Tab.2 Number of depth-time curves in each region
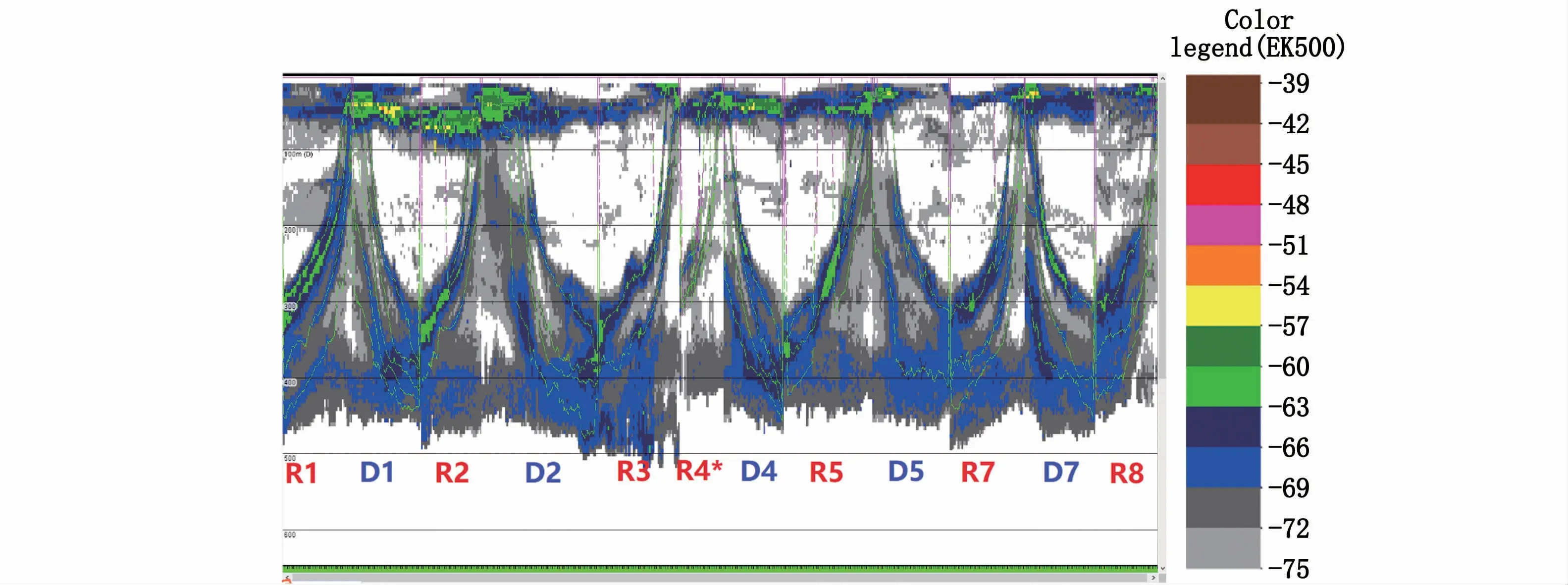
图4 Echoview软件中获得的垂直迁移原始深度时间曲线(绿色)Fig.4 Raw depth-time curves(green)acquired by Echoview
迁移速率集中分布在0~7.2 cm·s-1,上升速率峰值位置(1.6 cm·s-1)小于下降速率峰值位置(2.0 cm·s-1)(图5)。
各层生物上升开始时间介于16∶33~17∶31,约0.97 h;结束时间介于19∶26~19∶39,约0.21 h;上升持续时长介于1.95~3.10 h,上升速率均值为3.41 cm·s-1(表3)。
表3 各生物分层上升参数汇总Tab.3 Summary of ascending parameters for biological stratifications
各层生物下降开始时间介于5∶47 ~6∶18,约0.51 h;结束时间介于6∶43~8∶53,约2.18 h;下降持续时长介于1.98~3.06 h;下降速率均值为4.30 cm·s-1(表4)。
表4 各生物分层下降数据汇总Tab.4 Summary of descending parameters for biological stratifications
将上升或下降的所有生物层的NVV按1 m间隔水层统计算术平均值,即得1 m间隔水层平均迁移速率(图6)。结果表明,200 m以浅,上升和下降的各水层平均迁移速率相差较小且明显大于200 m以深的水层;200 m以深,下降的各水层平均迁移速率远大于上升的。
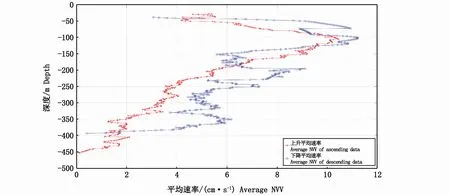
图6 1 m间隔水层平均上升和下降迁移速率Fig.6 Average net vertical rate of ascending and descending data by 1 m vertical separation
2.3 迁移层的分离与聚合
生物分层在迁移过程中会发生分离和聚合的现象。分离现象多发生在上升过程中,且表现为一分二(图7)。本研究中,6个上升区域有4个区域发生分离现象,分别为R2、R3、R5和R7区域;5个下降区域仅有D4区域发生分离现象(图8)。因此,11个迁移研究区域中共有5个发生分离现象,占比45.5%。
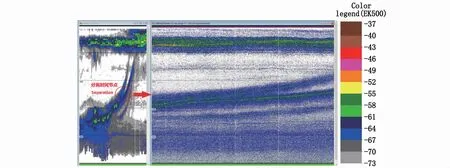
图7 R2区域发生的生物层分离示意图Fig.7 Separation of biological stratifications in region R2
聚合现象多发生在下降过程中,信号较弱且多发生在200 m以浅水域,表现为二合一(图9)。本研究中,6个上升区域中仅有R5区域发生聚合现象;5个下降区域有2个发生聚合现象,分别为D2和D4区域。因此,在11个迁移研究区域中共有3个区域发生聚合现象,占比27.3%。
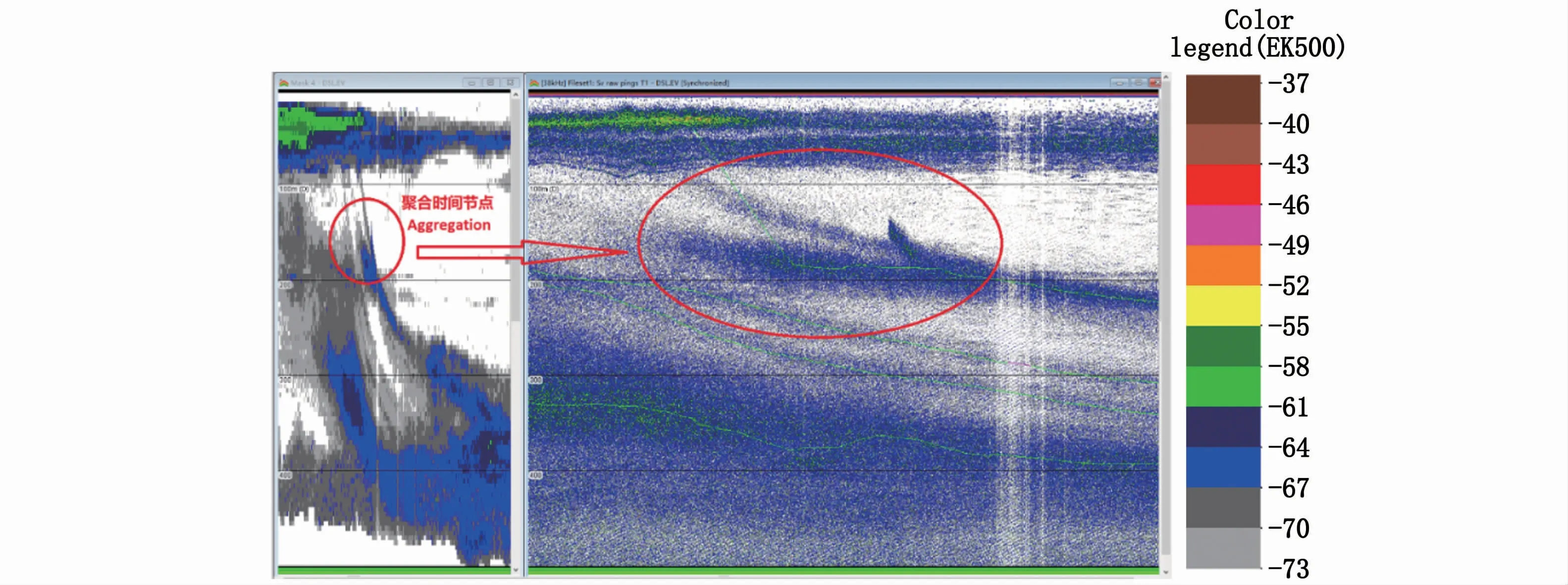
图9 D2区域发生的生物层聚合示意图Fig.9 Aggregation of biological stratifications in region D2
仅R5一个区域同时出现生物层的分离与聚合,表现为两个生物层在上升开始后逐渐汇聚为一个生物层,在水深约200 m时再次分离为两个生物层。
2.4 迁移量
向上迁移过程中,生物从DSL水层进入并停留在ML和SL水层。调查期间6个上升活动前后各水层NASC值的变化如表5。DSL水层的NASC值上升前为1 323.23~2 829.13 m2·nmi-2,减少量ΔNASCDSL为982.72~2 575.70 m2·nmi-2,变化百分比为62.63% ~92.70%;SL水层NASC值增加量ΔNASCSL为984.12~2 633.73m2·nmi-2,占ΔNASCDSL的百分比为59.92% ~260.46%,其中R5、R7和R8区域约为100.00%;ML水层NASC值增加量ΔNASCML为-103.20~56.52 m2·nmi-2,占ΔNASCDSL的百分比为-8.31% ~2.95%;未进入SL和ML水层的ΔNASCother为-1 606.53~1 135.48 m2·nmi-2,占ΔNASCDSL的百分比为-158.83%~44.08%。
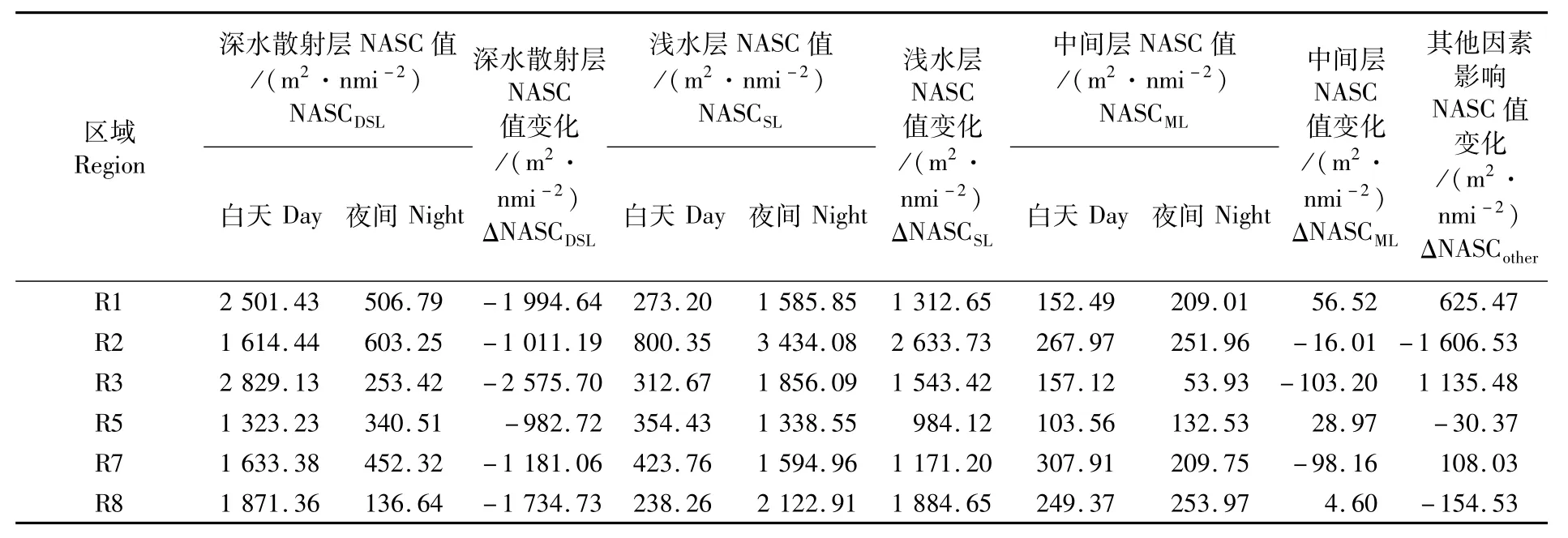
表5 上升前后各水层NASC值变化情况Tab.5 Variation of NASC in biological stratifications before and after ascending
向下迁移过程中,生物从SL水层向下运动进入并停留在ML和DSL水层。调查期间5个下降活动前后各水层NASC值的变化如表6。生物下降前后,SL水层NASC值减少量ΔNASCSL为907.25 ~2 468.54 m2·nmi-2;DSL水层NASC值增加后为946.90~2 203.13 m2·nmi-2,增加量ΔNASCDSL为431.73~1 757.91 m2·nmi-2,变化百分比为45.59% ~79.79%,占SL水层减少量ΔNASCSL的百分比为32.71% ~98.67%;ML水层NASC值增加量ΔNASCML为-319.97~-115.35 m2·nmi-2,占ΔNASCSL的百分比为-18.56%~-7.05%;未进入DSL和ML水层的ΔNASCother为149.25~1 889.58 m2·nmi-2,占ΔNASCSL的百分比为8.38%~87.68%。

表6 下降前后各水层NASC值变化情况Tab.6 Variation of NASC in biological stratifications before and after descending
3 讨论
调查海域DSL水层生物黄昏时分上浮,黎明时分下沉,规律明显(图2),属于常规类型的DVM[2]。迁移活动分层进行(图4),分层数量为2~5层,生物分层迁移是DVM 的一个普遍特征[2,4,21]。根据逃避捕食者假说(predator evasion hypothesis),生物受视觉捕食者的影响程度不同[2],小尺度物种更早到达而更晚离开表层[22],同种不同脂肪储备的个体在表层摄食时长也不同[23];此外,白天和夜间不同种类的物种聚集于不同水层[12],这些因素均可能是生物分层迁移及迁移层分离与聚合现象的诱因。塞拉利昂海域迁移生物分层的种类组成和分层诱因还有待进一步的调查研究。
本文对每个生物分层的迁移规律进行了分析,结果表明,各生物分层垂直迁移的起始时间、起始深度、抵达深度和持续时间等参数均不相同(表3,表4)。例如各生物层起始迁移时间在不同采样点或相同采样点均不相同。上升活动开始时间,不同采样点最大相差约58 min,相同采样点最大相差约104min(表3);下降活动开始时间,不同采样点最大相差约31 min,相同采样点最大相差约47 min(表4)。基于Gilliam’s rule的权衡(trade-off)模型说明浮游动物的DVM起始时间具有个体大小依赖性,在其他影响条件相同的情况下,个体越大,越容易被视觉性捕食者发现,因此会越晚上升而越早下降[22]。萨尼奇湾口中太平洋磷虾(Euphausiapacifica)的小个体相对于大个体提前30 min上升,落后45 min下降[21]。根据权衡模型的推论,个体尺寸相差较大的不同物种迁移活动起始时间可能相差更大。
本研究中下降速率均值4.30 cm·s-1大于上升速率均值3.41 cm·s-1(表3,表4),说明生物以更快的速度下降。最大上升速率为24.98 cm·s-1(约100 m),最大下降速率为29.15 cm·s-1(约200 m)。声学记录到的生物快速游动和高机动性行为被认为与鱿鱼行为特性相关[18]。对短鳍鱿鱼(Illexillecebrosus)游泳行为的研究表明,其经济游泳速度(economical speed)为34~50 cm·s-1(不考虑游泳角度)[24]。结合拖网样本,本研究记录到的高速迁移数据可能与鱿鱼有关。
迁移活动是变速运动,与中国南海相关研究结果一致[4]。本研究中上升活动一般是加速运动(图10),下降活动一般是减速运动(图11),说明生物在较浅水域的迁移速率大于较深水域的。本研究表明,200 m以浅水域生物层的迁移速度大于200 m以深水域(图6),造成这一现象的原因还需要进一步的研究论证。
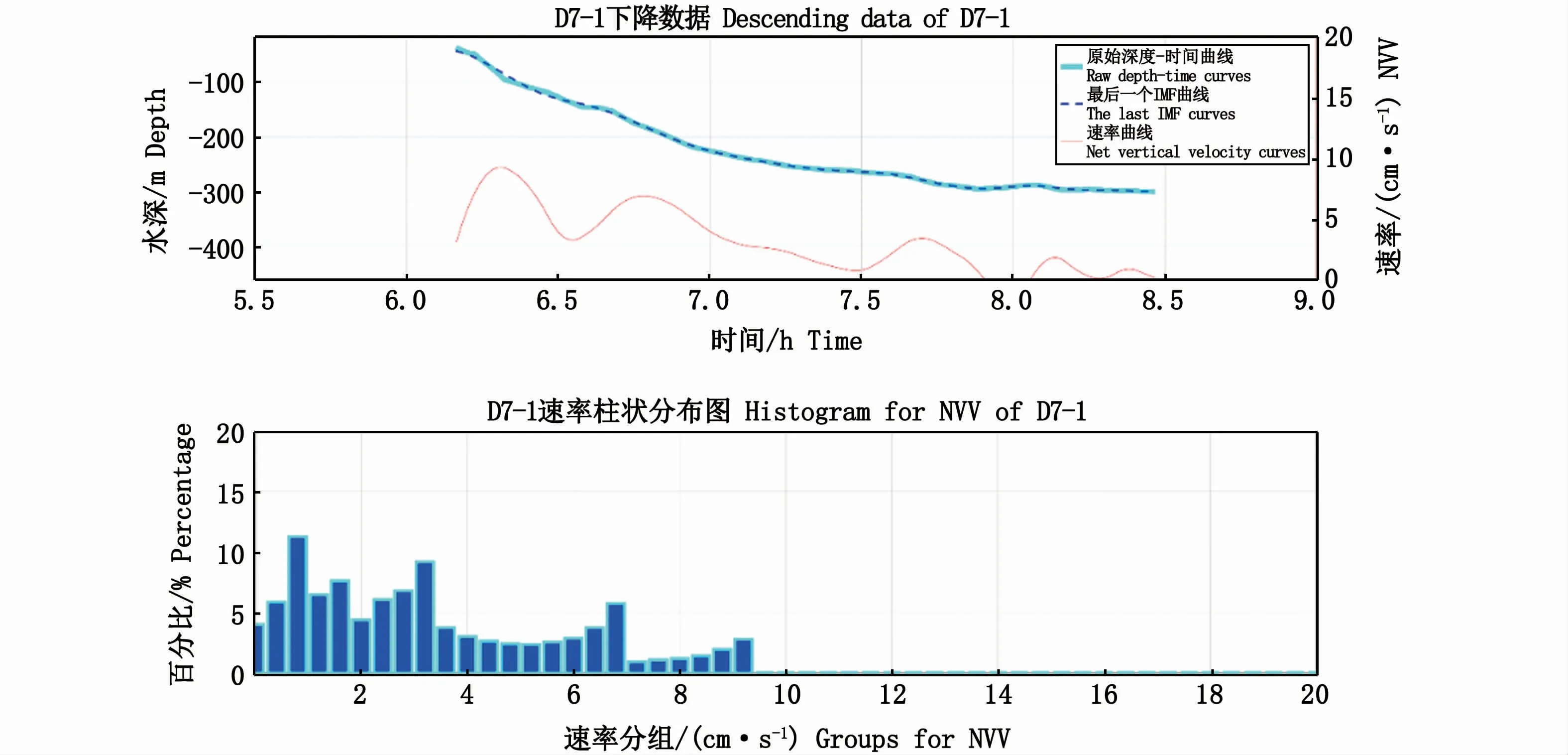
图11 D7-1下降曲线及下降速率Fig.11 D7-1 descending curve and net vertical velocity
DSL水层的NASC 值介于1 338.55~3 328.10 m2·nmi-2(表5,表6),处于热带和亚热带海域DSL值的记录范围158~7 617 m2·nmi-2内[8];DSL水层NASC值变化百分比的算术平均值为69.95%,为便于与KLEVJER等[3]的研究结果做对比,将本文数据应用到MP(proportion of animalsmigrating)公式中,得出本文的MP算术平均值为0.75,高于之前文献报道中北大西洋的MP值0.38,说明调查期间塞拉利昂海域有更多的深水散射层生物参与迁移活动。但由于DVM存在明显的地理和季节性差异[3,22],MP值仅做参考,不具普遍意义。本文不同地理位置的迁移生物量及迁移百分比就存在较大差异(表5,表6)。
短时间内,在不考虑生物交换和内部消耗时,同一水域内声学回波积分结果应保持不变。但是,由于探测设备性能[5]、生物时空分布差异[6]、垂直迁移前后生物体结构的改变[7,25-26]等都会影响到声学回波积分结果。因此,本文引入ΔNASCother变量来衡量引起声学积分结果异常的其他潜在因素的综合影响。
上升后,DSL减少的生物绝大部分进入SL,其他可忽略不计;下降后,SL减少的生物大部分进入DSL,小部分停留在ML,还有一部分受潜在因素影响消失在调查设备探测能力以外(表5,表6)。受潜在因素影响消失的信号,既可能是生物集群密度降低,造成集群信号淹没于背景噪声;也可能是上下水层洋流不同造成生物斑块的水平移动[2],或者可能是某些物种下潜到38 kHz换能器的探测范围之外[18]。具体诱因是哪种潜在因素还有待后续研究。
