不同轮耕模式对豫东地区小麦—花生轮作田土壤质量、团聚体组成及稳定性的影响
2023-09-11袁亮
袁亮


摘要:为探明不同轮耕模式对豫东地区小麦→花生轮作田土壤质量、团聚体组成及稳定性的影响。于2018—2022年,通过田间定位试验,以连续旋耕(RT)处理为对照,设置旋耕-深翻-旋耕-深翻(RDRD)处理、旋耕-免耕-旋耕-免耕(RNRN)处理、深翻-免耕-深翻-免耕(DNDN)处理、旋耕-免耕-深翻-免耕(RDN)处理共4种轮耕模式,研究不同轮耕模式对土壤养分含量、酶活性、容重、孔隙度、团聚体组成及稳定性的影响。结果表明,与RT处理相比,不同轮耕模式均能提高土壤碱解氮、速效磷、速效钾、有机质的含量及土壤脲酶、碱性磷酸酶的活性;不同轮耕模式能够降低土壤容重,提高孔隙度,促进>0.25 mm粒级团聚体形成,提高团聚体平均质量直径 (MWD)、几何平均直径 (GMD)和>0.25 mm粒级团聚体含量(R0.25),降低团聚体分形维数(D)。其中,0~20 cm土层中,与RT处理相比,RDN处理容重显著降低6.25%(P<0.05),孔隙度、MWD、GMD、R0.25分别显著提高13.66%、15.22%、17.80%、10.90%。RDN处理>2 mm粒径团聚体比例最高,较RDRD、RNRN、DNDN处理分别显著提高16.50%、29.19%、7.16%。20~40 cm土层中,与RT处理相比,RDN处理容重显著降低7.75%,孔隙度、MWD、GMD值分别显著提高14.97%、9.80%、8.33%。DNDN处理>2 mm粒径团聚体比例最高,较RNRN处理显著增加8.18%,与RDRD、RDN处理相比均无显著性差异。相关性分析表明,土壤养分含量及酶活性的变化与土壤物理结构密切相关。综上所述,旋耕-免耕-深翻-免耕处理表现最优。
关键词:轮耕;养分;酶活性;容重;孔隙度;团聚体
中图分类号:S158.3;S152.3;S158.5 文献标志码:A
文章编号:1002-1302(2023)15-0245-08
基金项目:河南省软科学研究计划项目(编号:18200410429)。
作者简介:袁 亮(1982—),男,安徽亳州人,硕士,讲师,主要从事微生物应用研究。E-mail:sqzyyl@163.com。
豫东地区是河南省粮食主产区,其粮食产量常年稳定在60亿kg以上,被誉为“豫东粮仓”[1-2]。小麦→花生轮作是豫东地区常见的种植模式之一,其土壤耕作方式长期实行旋耕模式。然而,长期单一旋耕整地不仅会造成土壤耕层变浅、犁底层上升、土壤酶活性降低等问题,还会阻碍作物根系深层分布,影响根系对水肥的吸收利用能力[3-5]。因此,研究适宜的轮耕模式对小麦—花生轮作田土地持续生产力的提高具有重要意义。研究表明,频繁耕作不仅会破坏土壤团聚体结构,降低土壤团聚体稳定性与土壤容重,还会显著降低土壤蓄水能力与养分供应能力,造成作物产量下降,品质降低[6-7]。短期免耕能够减少土壤扰动,改善土壤微生态环境,提高土壤微生物群落功能多样性,而长期免耕会造成表层土壤养分富集、深层土壤养分匮乏、土壤容重变大、孔隙度降低等问题[8-9]。可见,长期单一的耕作模式不利于保持土壤结构的稳定性,对农作物生长发育及土壤质量的提高均有不利影响[10-13]。近年来,有专家学者提出,将不同耕作方式按照一定顺序进行合理组合与配置,能够消除长期单一耕作模式带来的问题[14]。陈宁宁等的研究表明,轮耕有利于改善土壤耕层物理结构,促进土壤大团聚体形成,对土壤蓄水保墒能力的提高具有积极的促进作用[15];龙潜的研究表明,合理轮耕有利于降低土壤容重,提高土壤孔隙度,提高土壤养分含量和作物产量[16]。于淑婷等的研究表明,深耕-旋耕-旋耕模式能够明显改善土壤耕层结构,提高土壤酶活性[17]。可见,合理的轮耕模式能够改善土壤耕层结构,提升土壤质量,提高土地生产力。
土壤酶是土壤生态系统的重要组成部分,能够参与土壤养分分解与转化、微生物群落代谢活动等大多数生物化学过程,是评价土壤质量的重要指标[18]。土壤团聚体是土壤结构的基本单元,能够参与土壤中大多数物质交换与能量流动等过程,其团聚体粒级变化对土壤结构及肥力变化具有较大影响[19-20]。近年来,关于轮耕的研究主要单一地集中在旋耕结合免耕对土壤理化性状、酶活性、团聚体的影响方面[21-23]。关于深翻、旋耕、免耕轮耕模式对不同土层土壤质量、团聚体组成、稳定性变化及其相关性的研究很少,而关于豫东地区小麦—花生轮作区轮耕模式对土壤结构及质量的研究更是鲜有报道。且不同地区气候条件、土壤类型、种植制度均有所不同,不同轮耕模式对土壤质量、团聚体组成及稳定性的变化并不相同。因此,本试验以连续旋耕为对照,研究连续4年不同轮耕模式对土壤养分、酶活性、容重、孔隙度、团聚体组成及稳定性的影响。找到适宜的轮耕模式,旨在为豫东地区小麦—花生轮作田耕作模式的合理选择提供理论依据。
1 材料与方法
1.1 试验区概况
试验于2018—2022年在河南省商丘市睢阳区冯桥镇曹集村(116°43′12″E、39°28′11″N)进行。研究区为华北平原典型的小麦—花生一年两熟区(冬季小麦,夏季花生),气候类型属大陆性季风气候,冬季寒冷干燥,夏季湿热多雨,四季分明。年平均降水量695 mm,主要集中在6—9月,年平均氣温14.2 ℃,年平均日照时数1 944 h,无霜期约211 d。供试土壤为黄潮土,质地中壤。试验前0~20 cm 土壤理化性质:碱解氮含量62.35 mg/kg,速效磷含量48.19 mg/kg,速效钾含量135.89 mg/kg,有机质含量15.26 g/kg,容重1.46 g/cm3,pH值8.05;20~40 cm 土壤理化性质:碱解氮含量31.29 mg/kg,速效磷含量25.97 mg/kg,速效钾含量54.63 mg/kg,有机质含量9.68 g/kg,容重1.52 g/cm3,pH值8.11。
1.2 试验设计
试验采用冬小麦—夏花生生产体系,以2年4个生产季为1个周期,连续定点试验4年。试验采用单因素随机区组设计,分别设连续旋耕(RT)、旋耕-深翻-旋耕-深翻(RDRD)、旋耕-免耕-旋耕-免耕(RNRN)、深翻-免耕-深翻-免耕(DNDN)、旋耕-免耕-深翻-免耕(RDN)5个处理。每个处理3次重复,共计15个小区,小区面积56 m2。旋耕处理操作方法:前季作物收获后,秸秆直接粉碎还田,再通过旋耕机整地,旋耕深度为 15~20 cm;深翻处理操作方法:前季作物收获后,秸秆直接粉碎还田,然后通过铧式犁深翻1遍,翻耕深度为30~35 cm,再通过旋耕机整地;免耕处理操作方法:前季作物收获后,人工移走作物秸秆,通过粉碎机打碎后撒施在小区内,不进行旋耕处理。当季作物均按照常规播种方式进行。
小麦品种:周麦18(河南省周口市农业科学院),小麦播种量为375 kg/hm2,小麦季施肥为氮磷钾配方肥750 kg/hm2(配施比例N ∶P2O5 ∶K2O=8 ∶3 ∶7),小麦生育期为10月15日至次年6月8日;花生品种为豫花9326(河南省农业科学院经济作物研究所),种植密度为15万株/hm2,花生季施肥为复合肥(N ∶P2O5 ∶K2O=15 ∶15 ∶15) 900 kg/hm2,花生生育期为6月15日至10月10日。2季化肥均作为基肥一次性施入,生育期内不进行追肥。其他浇水、除草、杀虫等均与当地田间管理措施相同。
1.3 测试项目与方法
于2022年9月30日利用五点取样法采集0~20、20~40 cm土样,测定土壤养分含量、酶活性与团聚体比例。另外利用环刀采集0~20、20~40 cm土样,测定土壤容重、孔隙度。其中,土壤碱解氮、速效磷、速效钾、有机质的含量及pH值分别采用碱解扩散法、0.5 mol/L NaHCO3法、NH4OAc浸提-火焰光度法、重铬酸钾容量法-外加热法及水土比5 ∶1法测定[24];土壤脲酶、蔗糖酶、过氧化氢酶、碱性磷酸酶的活性分别采用苯酚钠比色法、3,5-二硝基水杨酸比色法、高锰酸钾滴定法、磷酸苯二钠比色法测定[25];土壤容重、孔隙度均采用环刀法测定[24];土壤团聚体组成采用干筛法测定,具体操作步骤详见惠超等的方法[26]。
1.4 计算方法
土壤团聚体几何平均直径 (GMD)、平均质量直径 (MWD)、分形维数(D)以及>0.25 mm粒级团聚体含量(R0.25)是表征土壤团聚体稳定性的重要指标[27]。计算公式如下:
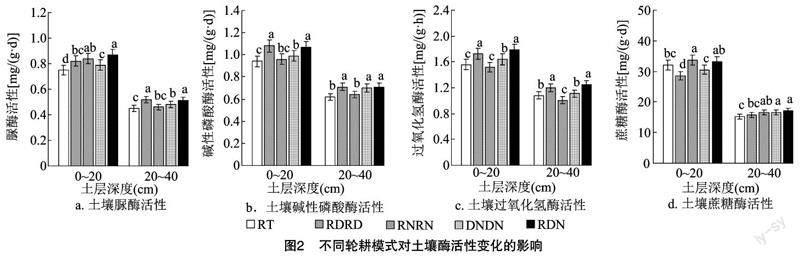
式中:Xi为某粒级团聚体平均直径;Wi为某粒级团聚体质量比值;Mi为某粒级团聚体质量;Mt为土壤团聚体总质量;W(δ 1.5 数据处理 本试验采用WPS Office软件进行数据整理、计算与作图,采用DPS 9.50数据处理软件利用Duncans新复极差法进行多重比较与相关性分析。 2 结果与分析 2.1 不同輪耕模式对土壤养分含量的影响 2.1.1 不同轮耕模式对土壤碱解氮含量的影响 由图1-a可知,不同耕作模式对不同土层碱解氮含量有不同的影响。0~20 cm土层中,与连续旋耕(RT)处理相比,不同轮耕(RDRD、RNRN、DNDN、RDN)处理土壤碱解氮含量均有不同程度的提高。其中,RNRN、RDN处理碱解氮含量较RT处理分别显著(P<0.05)提高6.24%、10.74%,RDRD、DNDN处理与RT处理差异不显著。不同轮耕处理中RDN处理的碱解氮含量最高,较RDRD、DNDN处理分别显著提高7.92%、7.77%,与RNRN处理相比无显著差异。20~40 cm土层中,与RT处理相比,RDRD、DNDN、RDN处理碱解氮含量分别显著提高9.85%、13.17%、17.55%,RNRN处理无显著性变化。不同轮耕处理中RDN处理碱解氮含量最高,较RDRD、RNRN处理分别显著提高7.00%、11.45%,与DNDN处理相比无显著性差异。 2.1.2 不同轮耕模式对土壤速效磷含量的影响 由图1-b可知,不同耕作处理土壤速效磷含量变化差异较大。0~20 cm土层中,与RT处理相比,不同轮耕处理土壤速效磷含量均有不同程度的提高。其中,RDRD、DNDN、RDN处理的速效磷含量较RT处理分别显著提高5.59%、5.04%、10.69%,RNRN处理无显著性变化。不同轮耕处理中RDN处理的速效磷含量最高,较RNRN处理显著提高5.74%,与RDRD、DNDN处理相比无显著性差异。20~40 cm 土层中,RDRD、DNDN、RDN处理的速效磷含量较RT处理分别显著提高7.00%、8.43%、11.45%,RNRN处理无显著性变化。不同轮耕处理中RDN处理速效磷含量最高,较RNRN处理显著提高10.36%,与RDRD、DNDN处理相比无显著性差异。RNRN处理速效磷含量显著低于其他轮耕处理。 2.1.3 不同轮耕模式对土壤速效钾含量的影响 由图1-c可知,0~20 cm土层中,与RT处理相比,不同轮耕处理土壤速效磷含量均有不同程度的提高。其中,RNRN、DNDN、RDN处理的速效钾含量较RT处理分别显著提高6.28%、6.99%、12.08%,RDRD处理无显著性变化。不同轮耕处理中RDN处理的速效钾含量最高,较RDRD、RNRN处理分别显著提高7.55%、5.46%,与DNDN处理相比无显著性差异。20~40 cm土层中,DNDN、RDN处理速效钾含量较RT处理分别显著提高6.38%、11.85%,RDRD、RNRN处理无显著性变化。不同轮耕处理中RDN处理的速效钾含量最高,较RDRD、RNRN处理分别显著提高6.99%、9.34%,与DNDN处理相比无显著性差异。 2.1.4 不同轮耕模式对土壤有机质含量的影响 由图1-d可知,0~20 cm土层中,与RT处理相比,RDN处理有机质含量显著提高6.28%,其他轮耕处理与RT处理相比均无显著变化。不同轮耕处理中RDN处理有机质含量最高,较RNRN处理显著提高5.56%,与RDRD、DNDN处理相比均无显著性差异,其他轮耕处理间也均无显著性差异。20~40 cm土层中,RT处理有机质含量最低,DNDN处理有机质含量最高,但不同处理间土壤有机质含量均无显著性差异。 2.2 不同轮耕模式对土壤酶活性变化的影响 2.2.1 不同轮耕模式对土壤脲酶活性的影响 由图2-a可知,0~20 cm土层中,与RT处理相比,不同轮耕处理的脲酶活性显著提高5.33%~16.00%。不同轮耕处理中RDN处理的脲酶活性最高,较RDRD、DNDN处理分别显著提高6.10%、10.13%,与RNRN处理相比无显著性差异。20~40 cm 土层中,与RT处理相比,RDRD、DNDN、RDN处理脲酶活性分别显著提高15.56%、6.67%、13.33%,RNRN处理无显著性变化。不同轮耕处理中RDRD处理的脲酶活性最高,较RNRN、DNDN处理分别显著提高13.04%、8.33%,与RDN处理相比无显著性差异。 2.2.2 不同轮耕模式对土壤碱性磷酸酶活性的影响 由图2-b可知,0~20 cm土层中,与RT处理相比,不同轮耕处理碱性磷酸酶活性均有不同程度的提高。其中,RDRD、DNDN、RDN处理的碱性磷酸酶活性较RT处理分别显著提高14.89%、5.32%、13.83%,RNRN处理无显著性变化。不同轮耕处理中RDRD处理碱性磷酸酶活性最高,较RNRN、DNDN处理分别显著提高12.50%、9.09%,与RDN处理相比无显著性差异。20~40 cm土层中,与RT处理相比,RDRD、DNDN、RDN处理的碱性磷酸酶活性分别显著提高14.52%、12.90%、14.52%,RNRN处理无显著性变化。不同轮耕处理中RDRD和RDN处理的碱性磷酸酶活性均最高,均较RNRN处理显著提高10.94%,与DNDN处理相比无显著性差异。RNRN处理碱性磷酸酶活性显著低于其他轮耕处理。 2.2.3 不同轮耕模式对土壤过氧化氢酶活性的影响 由图2-c可知,0~20 cm土层中,与RT处理相比,RDRD、DNDN、RDN处理的过氧化氢酶活性分别显著提高10.90%、5.13%、14.74%,RNRN处理过氧化氢酶活性降低,但无显著性差异。不同轮耕处理中RDN处理的过氧化氢酶活性最高,较RNRN、DNDN处理分别显著提高17.76%、9.15%,与RDRD处理相比无显著性差异。RDRD处理过氧化氢酶活性也均显著高于RNRN、DNDN处理。20~40 cm土层中,与RT处理相比,RDRD、RDN处理的过氧化氢酶活性分别显著提高11.11%、15.74%,RNRN处理则显著降低6.48%,DNDN处理无显著性变化。不同轮耕处理中RDN处理的过氧化氢酶活性最高,较RNRN、DNDN处理分别显著提高23.76%、12.61%,与RDRD处理相比无显著性差异。RNRN处理的过氧化氢酶活性显著低于其他处理。 2.2.4 不同轮耕模式对土壤蔗糖酶活性的影响 由图2-d可知,0~20 cm土层中,与RT处理相比,RNRN处理的蔗糖酶活性显著提高5.05%,RDRD处理显著降低11.14%,DNDN、RDN处理无显著性变化。不同轮耕处理中,RNRN處理蔗糖酶活性最高,较RDRD、DNDN处理分别显著提高18.22%、10.17%,与RDN处理相比无显著性差异。20~40 cm 土层中,与RT处理相比,RNRN、DNDN、RDN处理的蔗糖酶活性分别显著提高8.01%、9.25%、11.48%,RDRD处理无显著性变化。不同轮耕处理中RDN处理蔗糖酶活性最高,较RDRD处理显著提高8.15%,与RNRN、DNDN处理相比无显著性差异。 2.3 不同轮耕模式对土壤容重的影响 由图3-a可知 0~20 cm土层中 与RT处理相比,不同轮耕处理土壤容重均降低。其中,RDN处理土壤容重较RT处理显著降低6.25%,其他轮耕处理均无显著性变化。不同轮耕处理的土壤容重均无显著性差异。20~40 cm土层中,与RT处理相比,不同轮耕处理土壤容重均降低。其中,RDRD、DNDN、RDN处理土壤容重较RT处理分别显著降低5.63%、5.63%、7.75%,RNRN处理无显著性变化。不同轮耕处理中RDN处理的土壤容重最低,较RNRN处理显著降低5.07%,与RDRD、DNDN处理相比无显著性差异。 2.4 不同轮耕模式对土壤孔隙度的影响 由图3-b可知,0~20 cm土层中,与RT处理相比,不同轮耕处理的土壤孔隙度均提高。其中,RNRN、DNDN、RDN处理土壤孔隙度较RT处理分别显著提高10.64%、5.36%、13.66%,RDRD处理无显著性变化。不同轮耕处理中RDN处理的土壤孔隙度最大,较RDRD、DNDN处理分别显著提高8.81%、7.88%,与RNRN处理相比无显著性差异。20~40 cm土层中,与RT处理相比,不同轮耕处理土壤孔隙度显著提高5.00%~14.97%。不同轮耕处理中RDN处理土壤孔隙度最大,较RDRD、DNDN处理分别显著提高9.49%、7.39%,与RNRN处理相比无显著性差异。 2.5 不同轮耕模式对土壤团聚体组成的影响 不同耕作条件下,不同土层土壤团聚体组成存在显著差异。由图4可知,0~20 cm土层各处理土壤团聚体组成均以>2.000 mm(46.56%~64.80%)、>0.250~2.000 mm(20.46%~30.32%)粒径团聚体为主,>0.053~0.250 mm(9.15%~12.54%)、≤0.053 mm(5.59%~10.58%)粒径团聚体比例较少。与RT处理相比,不同轮耕处理中>2.000 mm粒径团聚体比例显著增加7.73%~39.18%。其中,RDN处理>2.000 mm粒径团聚体比例最高,较RDRD、RNRN、DNDN处理分别显著增加16.50%、29.19%、7.16%。不同轮耕处理>0.250~2.000 mm、>0.053~0.250 mm、≤0.053 mm 粒径团聚体比例较RT处理均不同程度降低。其中,除RNRN处理>0.053~0.250 mm粒径团聚体比例无显著性变化外,各轮耕处理 ≤2.000 mm 粒径团聚体比例与RT处理相比均显著降低。 随着取样深度的增加,20~40 cm土层中,>2.000 mm(35.85%~41.16%)粒径团聚体比例较 0~20 cm土层明显减少,>0.250~2.000 mm(35.94%~39.46%)、>0.053~0.250 mm(11.42%~13.24%)、≤0.053 mm(9.64%~12.02%)粒径团聚体比例均增加。与RT处理相比,不同轮耕处理>2.000 mm粒径团聚体比例显著增加6.14%~14.81%。其中,DNDN处理>2.000 mm粒径团聚体比例最高,较RNRN处理显著增加8.17%,与RDRD、RDN处理相比无显著性差异。不同轮耕处理 >0.250~2.000 mm、>0.053~0.250 mm粒径团聚体比例均较RT处理降低;而RDRD、RNRN处理 ≤0.053 mm 粒径团聚体比例较RT处理增加,DNDN、RDN处理≤0.053 mm粒径团聚体比例较RT处理降低。 2.6 不同轮耕模式对土壤团聚体稳定性的影响 不同耕作条件下,不同土层土壤团聚体稳定性表现出明顯差异。由表1可知,0~20 cm土层中,与RT处理相比,不同轮耕处理土壤团聚体平均质量直径(MWD)、几何平均直径(GMD)、>0.25 mm团聚体质量分数(R0.25)均提高,分形维数(D)降低。其中,RDRD、DNDN、RDN处理土壤团聚体MWD和R0.25分别显著提高12.50%、5.43%、15.22%和5.35%、9.01%、10.90% RDRD、 RDN处理土壤团聚体GMD分别显著提高14.41%、17.80%,RNRN处理土壤团聚体MWD、GMD、R0.25以及DNDN处理的GMD均无显著性变化。不同轮耕处理的D较RT处理均无显著性变化。不同轮耕处理中RDN处理土壤团聚体MWD、GMD、R0.25均最高,D最低。20~40 cm土层中,与RT处理相比,RDRD和RDN处理的土壤团聚体MWD、GMD分别显著提高6.94%、8.93%和9.80%、8.33%,RNRN、DNDN处理的土壤团聚体MWD、GMD均无显著性变化。不同轮耕处理土壤团聚体R0.25较RT处理均提高,D均降低,但均无显著性差异。不同轮耕处理中RDN处理的土壤团聚体MWD、R0.25最高,RDRD处理的土壤团聚体GMD最高,但与RDN处理相比无显著差异。DNDN、RDN处理的D最低。 2.7 土壤各指标间的相关性 土壤养分含量、酶活性与土壤团聚体稳定性、容重及空隙度的相关分析见表2。可以看出,碱解氮含量、脲酶活性与孔隙度呈极显著正相关关系(P<0.01),与D、容重呈负相关关系;速效磷含量与GMD、MWD、R0.25及孔隙度呈显著正相关关系,与D呈显著负相关关系,与容重呈极显著负相关关系;速效钾含量与R0.25、孔隙度呈显著正相关关系,与D呈显著负相关关系,与容重呈极显著负相关关系;有机质含量与R0.25呈极显著正相关关系,与D、容重呈极显著负相关关系;碱性磷酸酶、过氧化氢酶活性与GMD、MWD呈极显著正相关关系,与D值、容重呈不显著负相关关系;蔗糖酶活性与GMD、MWD、R0.25呈不显著负相关关系。可见,土壤养分含量及酶活性的变化与土壤物理结构密切相关,不同轮耕措施在改变土壤结构的同时,也改变了土壤生物学特性,进而影响土壤养分转化及酶活性的提高。 3 讨论 农业耕作措施能够改变土壤结构,影响耕层土壤养分分布[4,28]。研究表明,轮耕能够均衡耕层土壤养分分布,避免养分富集或分布不均[29]。有研究表明,与传统耕作模式相比,合理的轮耕方式能够显著增加耕层土壤养分含量,提高作物产量[30-31]。 本研究表明,与RT处理相比,不同轮耕处理均能够不同程度地提高0~40 cm土层碱解氮、速效磷、速效钾及有机质的含量,这与大多数研究较为一致[14,32]。其中,旋耕-免耕-深翻-免耕(RDN)处理效果较为显著,分析认为,频繁的耕作措施能够增加对土壤的扰动,加速有机质矿化分解,但不利于有机质累积;免耕措施能够减少土壤扰动,提高微生物代谢活性,促进土壤养分转化与利用,但长期免耕促使土壤容重变大,空隙度减小,从而不利于土壤养分利用能力的提高。而旋耕/深翻/免耕相结合既可以有效疏松改善土壤结构,防止频繁耕作扰动导致的土壤挤压紧实,又可以避免长期免耕导致土壤容重变大,养分利用能力降低。且深翻能够打破犁底层,促进养分下移,促使养分在不同土层间均衡分布,避免表土层养分富集与浪费,从而提高土壤养分利用能力。 有研究表明,土地利用方式可以显著影响土壤酶活性变化,进而改变土壤生物化学反应的发生速度和方向[33-34]。本研究表明,与RT处理相比,不同轮耕模式均能够提高土壤脲酶、碱性磷酸酶活性,这与朱长伟等的研究[21]较为一致。其中RDN处理效果较为显著,结合土壤养分含量变化分析认为,旋耕、深翻、免耕模式处理既能够减少土壤扰动,提高土壤微生物群落代谢活性,且深翻促使养分及秸秆下移,为微生物代谢活动提供充足的能量与碳源,促进土壤养分转化与利用,从而提高相关酶的活性;又能避免土壤压实,适宜的土壤紧实度能够为土壤微生物活动提供良好的生存环境,从而有利于土壤酶活性提高。 土壤团聚体与土壤结构密切相关,其粒级分布与变化可以明显影响土壤孔隙结构、蓄水保肥能力和微生物代谢活动[35]。有研究表明,适宜的轮耕模式不仅可以改善土壤团聚体结构,提高土壤团聚体稳定性,还能提高土壤透气性和蓄水保肥能力[36]。本研究表明,与连续旋耕(RT)处理相比,轮耕能够降低土壤容重,提高土壤孔隙度,促进>0.25 mm粒级团聚体形成,提高MWD、GMD、R0.25,降低D。其中RDN处理效果较为显著。分析认为,相比其他处理,RDN处理既能够减少土壤机械破坏,降低土壤容重,提高土壤孔隙度,提高根际微生物活力,又能够促使上层养分和秸秆下移,为微生物代谢活动提供充足能量,促使团聚体的胶结团聚与稳定,用于抵消耕作措施对土壤团聚体的部分破坏,延缓团聚体转化与分解,进而提高大团聚体比值。相关性分析表明,不同轮耕模式下土壤养分含量及酶活性变化与土壤物理结构密切相关,说明适宜的轮耕模式在促进土壤养分含量与酶活性提高的同时能够促进土壤大粒级团聚体的形成,有利于提高土壤团聚体稳定性。 4 结论 连续4年定位试验表明,与连续旋耕模式相比,不同轮耕模式均能提高0~40 cm土层碱解氮、速效磷、速效钾、有机质的含量及土壤脲酶、碱性磷酸酶的活性;不同轮耕模式能够降低土壤容重,提高土壤孔隙度,促进>0.25 mm粒级团聚体形成,提高MWD、GMD、R0.25,降低D。其中旋耕-免耕-深翻-免耕处理表现最优。 相关性分析表明,不同轮耕模式下土壤养分含量及酶活性变化与土壤物理结构密切相关。由此可知,不同轮耕模式在改变土壤结构的同时,也改变了土壤生物学特性,进而影响土壤养分转化及酶活性的提高。 参考文献: [1]张正斌,段子渊,王丽芳,等. 黄淮南片粮仓现代农业发展战略[J]. 中国生态农业学报,2017,25(3):309-315. [2]秦文臣. 商丘地区农业科技创新问题与对策研究[D]. 南京:南京农业大学,2016. [3]韩 上,武 际,李 敏,等. 深耕结合秸秆还田提高作物产量并改善耕层薄化土壤理化性质[J]. 植物营养与肥料学报,2020,26(2):276-284. [4]郭书亚,尚 赏,汤其宁,等. 不同轮耕方式与生物炭对土壤酶活性、土壤养分及小麦和玉米产量的影响[J]. 作物杂志,2022(3):211-217. [5]王 幸,邢兴华,徐泽俊,等. 耕作方式和秸秆还田对黄淮海夏大豆产量和土壤理化性状的影响[J]. 中国油料作物学报,2017,39(6):834-841. [6]Yan L,Jiang X X,Ji X N,et al. Distribution of water-stable aggregates under soil tillage practices in a black soil hillslope cropland in Northeast China[J]. Journal of Soils and Sediments,2020,20(1):24-31. [7]李 榮,侯贤清,贾志宽,等. 北方旱作区土壤轮耕技术研究进展[J]. 西北农业学报,2015,24(3):1-7. [8]沈晓琳,王丽丽,汪 洋,等. 保护性耕作对土壤团聚体、微生物及线虫群落的影响研究进展[J]. 农业资源与环境学报,2020,37(3):361-370. [9]贺建华,王 平,陈 娟,等. 不同轮耕方式对旱作区土壤容重、水分及春玉米产量的影响[J]. 土壤通报,2018,49(2):415-422. [10]谷思玉,朱玉伟,郭兴军,等. 不同耕作方式下黑土物理性状及其对玉米苗期生长的影响[J]. 华北农学报,2018,33(4):226-231. [11]王淑兰,王 浩,李 娟,等. 不同耕作方式下长期秸秆还田对旱作春玉米田土壤碳、氮、水含量及产量的影响[J]. 应用生态学报,2016,27(5):1530-1540. [12]周文涛,毛 燕,唐志伟,等. 长期定位试验不同耕作方式与秸秆还田对水稻产量和稻米品质的影响[J]. 中国稻米,2021,27(5):45-49. [13]周鹏翀,沈 莹,许姣姣,等. 长期定位耕作方式下冬小麦田根系呼吸对土壤呼吸的贡献[J]. 农业资源与环境学报,2019,36(6):766-773. [14]杨佳宇,谷思玉,李宇航,等. 深翻-旋耕轮耕与有机肥配施对黑土农田土壤物理性质的影响[J]. 土壤通报,2021,52(6):1290-1298. [15]陈宁宁,李 军,吕 薇,等. 不同轮耕方式对渭北旱塬麦玉轮作田土壤物理性状与产量的影响[J]. 中国生态农业学报,2015,23(9):1102-1111. [16]龙 潜. 不同轮耕模式对潮土不同土层理化性状及作物产量的影响[D]. 郑州:河南农业大学,2019. [17]于淑婷,赵亚丽,王育红,等. 轮耕模式对黄淮海冬小麦—夏玉米两熟区农田土壤改良效应[J]. 中国农业科学,2017,50(11):2150-2165. [18]Singh G,Bhattacharyya R,Das T K,et al. Crop rotation and residue management effects on soil enzyme activities,glomalin and aggregate stability under zero tillage in the Indo-Gangetic Plains[J]. Soil and Tillage Research,2018,184:291-300. [19]Peng X H,Horn R,Hallett P.Soil structure and its functions in ecosystems:phase matter & scale matter[J]. Soil and Tillage Research,2015,146:1-3. [20]隋鹏祥,罗 洋,李瑞平,等. 不同耕作措施土壤结构特征及其影响因素的研究进展[J]. 玉米科学,2022,30(4):83-89. [21]朱长伟,孟威威,石 柯,等. 不同轮耕模式下小麦各生育时期土壤养分及酶活性变化特征[J]. 中国农业科学,2022,55(21):4237-4251. [22]霍 琳,杨思存,王成宝,等. 耕作方式对甘肃引黄灌区灌耕灰钙土团聚体分布及稳定性的影响[J]. 应用生态学报,2019,30(10):3463-3472. [23]张 琦,王 浩,王淑兰,等. 深松轮耕模式对黄土旱塬春玉米土壤理化性质和作物产量的影响[J]. 应用生态学报,2020,31(2):459-466. [24]鮑士旦. 土壤农化分析[M]. 3版.北京:中国农业出版社,2000. [25]关松荫.土壤酶及其研究法[M]. 北京:农业出版社,1986. [26]惠 超,杨卫君,宋世龙,等. 生物炭施用对麦田土壤团聚体机械稳定性及春小麦产量的影响[J]. 土壤通报,2022,53(2):349-355. [27]孙 雪,张玉铭,张丽娟,等. 长期添加外源有机物料对华北农田土壤团聚体有机碳组分的影响[J]. 中国生态农业学报(中英文),2021,29(8):1384-1396. [28]王玉玲,李 军,柏炜霞. 轮耕体系对黄土台塬麦玉轮作土壤生产性能的影响[J]. 农业工程学报,2015,31(1):107-116. [29]李 娟,李 军,尚金霞,等. 轮耕对渭北旱塬春玉米田土壤理化性状和产量的影响[J]. 中国生态农业学报,2012,20(7):867-873. [30]侯贤清,李 荣,韩清芳,等. 轮耕对宁南旱区土壤理化性状和旱地小麦产量的影响[J]. 土壤学报,2012,49(3):592-600. [31]龙 潜,董士刚,朱长伟,等. 不同耕作模式对小麦—玉米轮作下潮土养分和作物产量的影响[J]. 水土保持学报,2019,33(4):167-174,298. [32]王 平,陈 娟,谢成俊,等. 轮耕条件下土壤改良及春玉米增产增收效果研究[J]. 干旱地区农业研究,2018,36(5):59-67. [33]张会慧,王世标,王均睿,等. 三江平原湿地不同利用方式对土壤理化性质及团聚体组成的影响[J]. 生态学杂志,2019,38(6):1679-1687. [34]徐莹莹,王俊河,刘玉涛,等. 耕作与秸秆还田方式对连作玉米田根际微生物及酶活性的影响[J]. 黑龙江农业科学,2018(7):1-3. [35]周 泉,王龙昌,邢 毅,等. 秸秆覆盖条件下紫云英间作油菜的土壤团聚体及有机碳特征[J]. 应用生态学报,2019,30(4):1235-1242. [36]胡钧铭,陈胜男,韦翔华,等. 耕作对健康耕层结构的影响及发展趋势[J]. 农业资源与环境学报,2018,35(2):95-103.
