云南苍山西坡森林木本植物幼苗组成的海拔梯度格局及其年际动态*
2023-09-02冯哲乌俊杰于小莉孙继文王丽萍陈凤仙邓瑞民
冯哲,乌俊杰,于小莉,孙继文,王丽萍,陈凤仙,邓瑞民
(大理大学 农学与生物科学学院,云南 大理 671003)
群落物种多样性的研究对于认识群落的结构、功能和组成,了解群落的动态变化的内在机制具有重要意义[1],而植物群落的物种多样性沿环境梯度变化规律一直是生态学的研究热点[2]。在山地条件下,群落物种多样性对随海拔升高所导致的环境变化也同样敏感[3]。因此,了解山地植物群落分布、群落动态、物种组成对海拔梯度变化的响应,对于认识植物对环境的适应具有重要科学意义[4]。自20世纪以来,众多生态学家已开展物种多样性随海拔梯度变化的研究,如张璐等[5]对山地群落物种多样性的研究中发现物种多样性随海拔的升高呈递增趋势,较多的研究结果表明物种多样性随海拔的升高呈降低趋势[6-7],更多的研究者认为物种多样性在中等海拔地区,物种多样性达到最大值[3,8-9],极少研究者认为物种多样性与海拔梯度无关[10]。因此,对典型高山区域的物种多样性进行研究是十分必要的,可为高山植物物种多样性的垂直分布格局积累相应的基础资料。林下幼苗是森林生态系统的重要组成部分,在森林天然更新过程中发挥着重要作用[11]。对于大多数木本植物而言,幼苗阶段是植物个体生长最为脆弱,对环境变化最为敏感的时期[12-13]。而林下幼苗的动态和组成对森林更新、群落演替和群落组成具有重要作用,同时也是影响森林的物种多样性的重要因素之一[3]。因此,了解林下幼苗对环境因子的响应,可以预测森林群落的物种组成,为森林经管理提供科学指导。近年来,国内的生态学者们对幼苗进行了一系列研究,如群落幼苗组成与空间分布[14]、季节动态[3]以及月际动态[15]等。以上研究大多数基于中国中部或北部,而云南属低纬高原山地环境,拥有多种不同气候带和生态系统类型,素有中国“动植物王国”之称[16]。尽管前人针对苍山进行了多种研究,如杨涛等[17]对大理苍山东坡的种子植物多样性分布格局的研究,张萍等[18]对苍山西坡的植被垂直分布格局的研究等。但大部分研究只针对苍山东坡植物且并未考虑群落动态变化。本研究以云南苍山西坡3个海拔样地为平台,基于2020—2022年连续3年的木本植物幼苗调查数据,在群落水平上,对木本幼苗在不同海拔上的分布格局及其年际动态进行分析,旨在了解幼苗物种组成的海拔梯度格局及其年际动态,以期为该地区的植被恢复与生物多样性保护提供科研理论依据。
1 研究方法
1.1 研究区概况
研究地点位于云南省大理白族自治州境漾濞县苍山西镇苍山西坡(25.70°~25.72°N、100.04°~100.06°E)。该地区属于中亚热带气候类型,有明显的干、湿季之分,雨季为每年的5—10 月,年降雨量约 1 059.4 mm,年均温约 16.1 ℃[19]。
1.2 样地设置
2020年11月—2022年12月,在苍山西坡,分别在海拔2 503、2 612、2 895 m设立3块样地,样地规格均为30 m×30 m。将每个样方划分为9 个 10 m×10 m 的小样方,对每个小样方的4个端点的经、纬度分别记录后取均值为该样地的经纬度(表1)。

表1 苍山西坡3个样地概况
每个样地内横向建立8个2 m×2 m的幼苗样方(图1),每个幼苗样方间距为2 m,计8行,64个幼苗样方。幼苗小样方的四个角用PVC管固定,并用样方绳圈围。在3个大样地中共设192个幼苗小样方。在每个幼苗样方中,对所有地径<1 cm且苗高不低于10 cm的木本幼苗进行挂牌编号,并鉴定幼苗的物种名,记录存活状态。2020年11月进行第一次调查,此后每年11月复查1次。截至2022年12月已经完成3次调查工作。
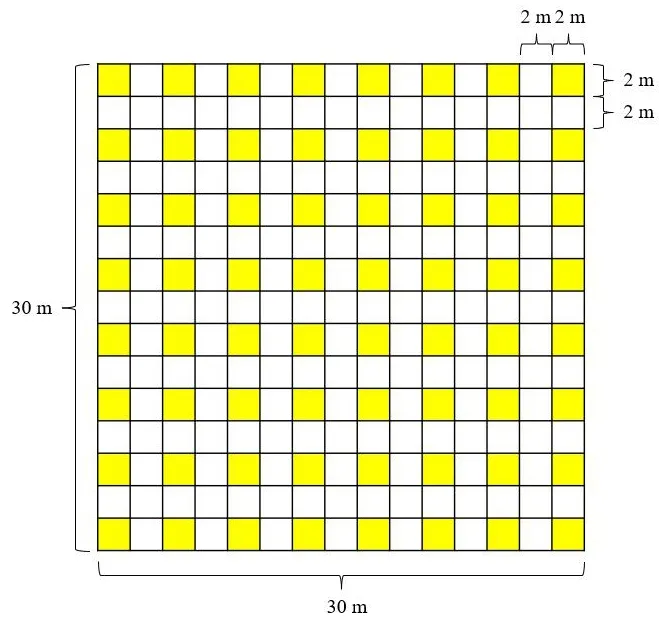
图1 幼苗样方分布
1.3 数据分析
(1)幼苗重要值(IV):
相对多度(RA)[20]
①
相对频度(RF)[20]:
②
重要值(IV)[21]:
③
式中:S和Si分别为某一样地幼苗总数和某一样地物种i的数量,Q和Qi分别为某一样地中样站总数和某一样地物种i出现的样站数。
(2)幼苗死亡率(m)[22]与更新率(r)[23]:
④

⑤
式中:N0和Nt分别为样地中第1次和第t次调查幼苗个体数,St为第t次调查时仍存活的个体数,T为两次调查的间隔时间。
(3)物种α多样性:
Simpson多样性指数[21]:
⑥
Shannon-Weiner多样性指数[21]:
⑦
Pielou均匀度指数[21]:
J=H/lnN
⑧
式中:N为物种数,Pi为第i种的个体数占全部物种总个体数的比例。
(4)Jaccard相似性指数反映不同海拔梯度之间样地物种的相似性程度,具体计算公式如下[24]:
⑨
式中:j为两个海拔样地群落共有的物种数;a、b分别为两个群落各自的物种数。
使用R软件vegan程序包计算幼苗物种α多样性与Jaccard相似性指数,其余分析与作图均在Microsoft Excel 2019和R 4.2.0(http://www.r-project.org/)中进行。
2 结果与分析
2.1 不同海拔梯度下的幼苗物种组成与数量特征
在2020—2022年期间共进行3次样地调查,3个样地共192个幼苗样方中共记录33种木本植物幼苗,分属19科28属。壳斗科(Fagaceae)为树种最多的科(5种)(表1)。主要木本植物幼苗为刺叶高山栎、金丝桃(Hypericummonogynum)、马缨杜鹃(Rhododendrondelavayi)和元江锥(Castanopsisorthacantha)(表2)。3个样方共有物种分别在相应样地中的占比为21.43%、18.75%、18.75%,特有物种分别于3个样地的占比为35.71%、25%、43.75%。调查期间,优势树种幼苗组成在年际间没有变化,刺叶高山栎始终具有明显优势地位,重要值均在前3位(表2),而不同树种的幼苗数量差异较大,样地重要值排名前6的物种数量之和分别占样地的84.1%、82.5%、79.8%(表2)。

表1 苍山样地物种名录

表2 不同海拔梯度木本植物幼苗重要值
2.2 幼苗物种的年际动态
在3个样地中,木本植物幼苗数量的年际差异较大,在2020年达到最大,随调查期间,幼苗数量整体呈下降趋势(表3)。在3次调查期间,共记录到22株新增幼苗,样地A、样地B和样地C分别增加11、3和8株,样地C在2022年未发现新增幼苗。新增幼苗物种9个,其中野扇花(Sarcococcaruscifolia)新增幼苗数量最多为7株,其次为水红木(Viburnumcylindricum)和溲疏属a(Deutziaa)种,新增数量均大于等于3株。在3次调查期间,共记录死亡幼苗109株,样地A、样地B和样地C分别死亡幼苗41、38和30株(表3)。
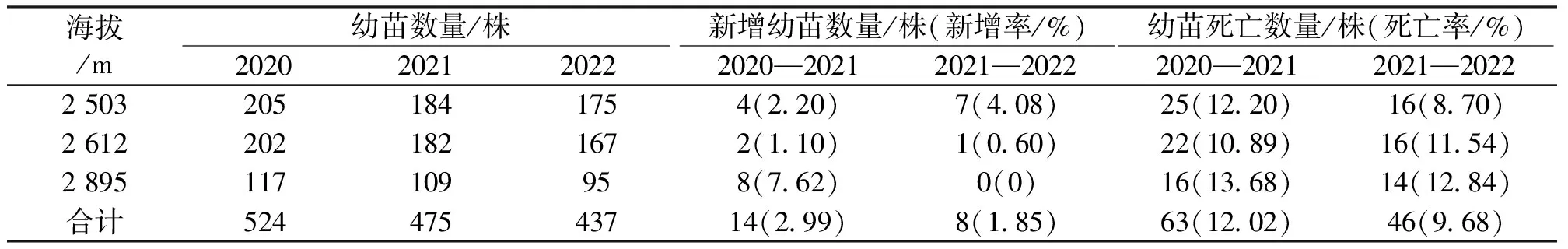
表3 不同海拔梯度幼苗动态变化
调查期间幼苗物种组成变化较小,仅样地C的第二次调查种新增物种白瑞香(Daphnepapyracea)。每个物种的重要值排名在不同年际间波动较小(表2)。3个样地的Simpson多样性指数、Pielou均匀度指数与Shannon-Weiner多样性指数随着调查时间的推移呈下降趋势,但在2021—2022年期间,样地B中Simpson指数与Pielou均匀度指数存在上升趋势(表4)。
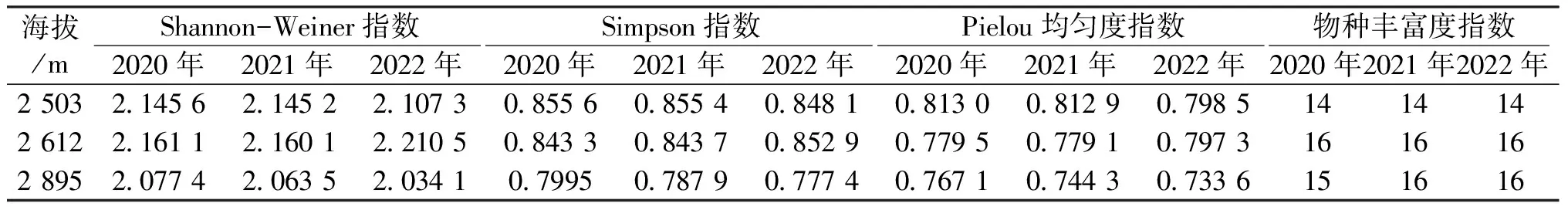
表4 不同海拔梯度物种多样指数
2.3 物种多样性随海拔的变化
幼苗物种丰富度随海拔上升呈现先增加后减少的趋势,在海拔2 612 m处达到最大值;Shannon-Weiner指数的变化趋势与物种丰富度变化基本一致,而Simpson多样性指数、Pielou均匀度指数及幼苗数量,均表现出随海拔升高而下降的趋势(表4)。
样地A与样地B的Jaccard相似性指数为0.363,样地A与样地C的Jaccard相似性指数为0.200,样地B与样地C的Jaccard相似性指数为0.333。Jaccard相似性指数结果显示随海拔升高不同海拔之间的共有种越来越少,物种组成的相似性越来越小。
3 讨论与结论
3.1 讨论
(1)幼苗树种组成
在2020—2022年研究期间内,幼苗数量累积记录物种共33种。样地内物种较丰富,大于长白山40 hm2样地中调查到的17个物种[25],但相比于西双版纳20 hm2样地的100多个物种还有很大差距[26],原因是长白山样地森林类型为阔叶红松林且环境较苍山西坡样地与西双版纳样地恶劣许多,而苍山西坡样地森林类型主要为中山湿性常绿阔叶林[27]较西双版纳样地的热带雨林森林类型缺乏更好的水热条件。在调查期间内,优势树种幼苗组成并未发生变化,群落内树种变化只有1株,但每个大样方之间差距很大,且随海拔升高,特有物种占比呈现先降低后增加趋势,并在样地C以43.75%占比最大,形成此现象的原因之一可能是由于不同海拔之间的环境异质性导致[3];另一个原因可能是因为特有物种大多数为稀有物种,由于稀有物种的幼苗的强扩散限制以及繁殖个体较少而导致此现象[28]。
在3年调查期间,累积记录幼苗共546株。不同树种的幼苗数量差异较大,样地重要值排名前6的物种数量之和分别占样地物种总数量的84.1%、82.5%、79.8%,说明在苍山西坡大样地优势物种的优势地位十分明显,且优势物种在之后的调查中均占很大比例,表明该林分更新良好。
(2)幼苗的年际动态
物种组成的年际变化波动不大,原因是在调查期间3个样地的稀有物种的幼苗更新和死亡率都比较低。而幼苗数量的年际变化较大的原因是由于随着幼苗龄级的增长,种内和种间竞争加剧所导致[14]。另一种原因是幼苗的新增和死亡驱动幼苗数量的年际变化[29]。
3个样地中Simpson多样性指数、Shannon-Weiner 多样性指数和Pielou均匀度指数随着调查时间推移呈降低趋势但差异并不明显,但在2021-2022年间,样地B中3个指数呈上升趋势,可能原因是优势物种溲疏属a种大量死亡,导致其优势度及均匀度降低,Simpson多样性指数及Pielou均匀度指数随之升高。Shannon-Weiner 多样性指数则是对个体密度变化敏感,溲疏属a种的死亡导致其他物种的个体密度上升,从而导致Shannon-Weiner 多样性指数的升高。其他调查期间内,幼苗数量的减少是物种多样性指数呈降低趋势的根本原因。
(3)幼苗的海拔分布格局
苍山样地的幼苗物种丰富度与Shannon-Weiner 多样性指数随海拔的升高呈现先增加后减少的分布格局,李洁琼等[3]对哀牢山和玉龙雪山不同海拔梯度样地的林下幼苗调查结果也发现类似现象。中海拔地区可能具有最优的水热条件,使得资源利用和物种多样性更高形成此现象[30]。邓莉萍等[31]对辽东山区次生林物种多样性的研究发现Simpson多样性指数与物种丰富度变化高度一致。但本文研究结果显示Simpson多样性指数随海拔的升高而降低,可能因为Simpson多样性指数是反映群落优势度的指标[31],随着海拔的升高优势种的集中度过高,从而导致了多样性的降低,这与唐志尧等[32]对秦岭太白山83个样地木本植物物种多样性的梯度格局研究结果一致。幼苗数量也呈现随海拔升高而减少的情况,原因可能因为海拔的升高导致水热因子发生变化,从而致使幼苗的大量死亡。在物种多样性的研究中,通常采用的测度指标如物种丰富度、多度、Shannon-Weiner 多样性指数、Simpson多样性指数等对尺度变化十分敏感[34],王志高等[34]对鼎湖山季风常绿阔叶林物种多样性指数空间分布特征的研究中发现,物种多样性的测度指标在80 m×100 m的尺度上反映最为敏感,而本研究仅分析30 m×30 m样地,尺度较小,可能导致无法正常反映物种多样性指数的变化,有关物种多样性随尺度变化的研究还有待今后进一步深入探索。
3.2 结论
2020—2022年间,样地内共累计记录到33种木本植物幼苗,分属19科28属。物种组成的年际变化不大,幼苗数量随时间推移逐渐减少,物种α多样性随时间推移逐渐降低,但在2021—2022年期间,样地B中物种α多样性呈现升高趋势,形成原因可能是物种溲疏属a种的大量死亡。在不同海拔梯度下,幼苗物种丰富度与Shannon-Weiner 多样性指数随海拔升高呈现先增加后减少趋势,Simpson多样性指数、Pielou均匀度指数与幼苗数量呈现随海拔升高而降低趋势。Jaccard相似性指数显示,随海拔的升高,两个群落之间的幼苗物种组成相似性呈现降低趋势。
