不同遮阴环境对广西火桐幼苗生长及生理生化的影响*
2023-09-02陆奇丰骆文华
陆奇丰,骆文华
(广西壮族自治区中国科学院广西植物研究所,广西 桂林 541006)
广西火桐(Firmianakwangsiensis)为广义锦葵科(Malvaceae s.l.)梧桐亚科(Sterculioideae)梧桐属(Firmiana)落叶乔木,国家一级重点保护野生植物[1-2]。该物种对研究梧桐属植物、亚非大陆间的物种亲缘关系以及植物区系地理分布方面具有重要的科研价值[3]。广西火桐为中国特有种,其树形优美、花色艳丽,具有极高的观赏价值,是珍贵的园林观赏树种[4]。此外,广西火桐生境区域极为狭窄,破碎化分布于我国广西中部至南部喀斯特地区,且由于人类的生产经营活动和过度开发利用,导致其野外种群和个体数量不断萎缩,已处于极度濒危状态[5]。目前,对广西火桐的研究主要集中于资源分布状况[6]、育苗技术[7-9]、光合特性[10]、遗传多样性[11]、传粉媒介[12]和叶绿体基因组系统进化[13-14]等方面。尚未见有光照对该物种幼苗生长和生理影响方面的相关报道,而该方面相关研究对探讨广西火桐的濒危机制具有重要意义。
光照是植物进行光合作用的能量来源,其不仅影响植物生长发育,还是影响种群更新的重要环境因子[15-17]。幼树幼苗对光照极为敏感,过强或过弱的光照均会影响幼树幼苗的生长发育,导致其生长停滞甚至死亡[16,18-19]。因此,研究探讨珍稀濒危物种的光适应性对其保护具有重要科学意义。了解珍稀濒危植物的光照需求,有助于对其进行科学抢救和制定有效管理措施[20-21]。物种的光适应性可通过其对不同光照条件下的形态和生理响应进行评估,其不仅广泛应用于农业、生态、园艺等领域,还被广泛运用于濒危植物最适生境条件的评估[16,21-22]。目前,已有研究通过该方法分析珍稀濒危植物对光照的适应性,如梓叶槭(Acercatalpifolium)[16]、Boltoniadecurrens[20]、Linderamelissifolia[22]、地枫皮(Illiciumdifengpi)[23]、闽楠(Phoebebournei)[24]等。
在前期野外调查中,项目组发现广西火桐不存在生殖障碍,种子能正常萌发,但幼苗保存率低,种群内幼树数量极少,且幼苗、幼树多分布于郁闭度较高的林下或岩隙内,其生境的透光率较低,幼苗生长状况较差。由此,引出如下相关问题:弱光对广西火桐幼苗生长及生理生化影响如何、光照是否是影响其更新的重要环境因子?对该问题的探讨不仅有助于深入了解广西火桐致濒的机制,亦可为广西火桐野外种群复壮和回归提供科学依据。
1 材料与方法
1.1 研究区概况
试验所用种子为2021年7月采自广西河池都安县金竹村弄盾屯(24°05′29″N、108°04′10″E)广西火桐成年健壮母树。种子采后及时脱粒,使用5‰高锰酸钾水溶液消毒,蒸馏水冲洗、晾干后贮藏于4 ℃冰箱中备用。同年8月,将种子置于培养皿中催芽,种子发芽后置于至高23 cm、口径20 cm的塑料盆中,基质均为采种地附近石山上的石灰土且重量一致,而后将其移至广西植物研究所试验用塑料大棚内。
1.2 试验方法及指标测定
将发芽2 d后的广西火桐种子栽植盆分别置于以下4种不同光照强度环境中:一层遮阴网(L1)、二层遮阴网(L2)、三层遮阴网(L3)和无遮阴网(CK)。遮阴网材料为市场上常售的3针黑色遮阴网(遮阳率约为60%),并利用照度计(testo 540,德国)测定晴朗天气1300时各处理的照度,分别为5 362、2 413、584、183 Lux。每个光照处理组均放置30盆(每盆含1粒发芽的种子),苗期保持一致的常规管护措施。
于2021年12月开展相关试验测定:每种遮光处理随机选择5株无病虫害、长势均匀的个体,利用MC-100叶绿素仪(Apogee,美国)测定叶片叶绿素含量指数(chlorophyll content index,CCI),然后计算叶片叶绿素含量(LC= 43.17e0.044CCI,μmol/m2)[25];测定其株高、基径和叶片数;整株收获,自来水冲洗根部,获取整体根系,测量主根长度及统计大于5 cm的侧根数;将叶片取下,利用叶面积仪(Scan Maker i800 plus,上海中晶科技有限公司)测量其面积及叶片长宽比;而后将所有样品置于烘箱干燥至恒重,计算不同光照条件下广西火桐幼苗的地上和地下部分生物量分配比。
另选择各处理组10株幼苗,取其上部成熟叶(1~3片),去除主叶脉后,用锡箔纸包裹并装入已编号的自封袋中,放入液氮中速冻,带回实验室后置于-80 ℃超低温冰箱保存,用于后续的生理生化指标测定。
根据李合生方法[26]测定广西火桐幼苗叶片的以下生理生化指标:愈创木酚法测定过氧化物酶(POD)活性、氮蓝四唑光还原法测定超氧化物歧化酶(SOD)活性、紫外吸收法测定过氧化氢酶(CAT)活性、硫代巴比妥酸比色法测定丙二醛(MDA)含量、磺基水杨酸法测定游离脯氨酸(Free proline)含量、考马斯亮蓝法测定可溶性蛋白质(Soluble protein)含量。可溶性糖(Soluble sugar)含量则采用硫酸-蒽酮法[27]测定。
1.3 数据分析
采用Microsoft Excel进行数据整理和作图,运用SPSS统计软件单因素方差分析(one-way ANOVA)及最小显著性差异法(least-significant difference,LSD)分析比较不同光照条件下广西火桐幼苗形态和生理指标的差异显著性(a=0.05)。图中的数据为平均值±标准差。
2 结果与分析
2.1 不同光照条件对广西火桐幼苗形态特征的影响
2.1.1 对生物量积累及分配的影响
光照对广西火桐幼苗的生物量积累有显著影响,见图1。
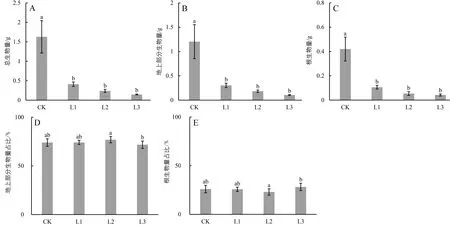
图1 不同光照条件对广西火桐幼苗生物量的影响
由图1可知,广西火桐幼苗的总生物量、地上部分生物量和根部生物量均随着光照的减弱呈降低趋势。与 CK相比,遮阴处理组的广西火桐幼苗总生物量、地上部分生物量和根部生物量均显著降低,分别下降了约75%(P<0.05)、85%(P<0.05)和91%(P<0.05)。表明在遮阴环境下,广西火桐幼苗的生长受到了极大的限制。然而,遮阴处理组的广西火桐幼苗地上部分和根的生物量分配比与CK无显著差异(P>0.05),在遮阴处理组内,除L2和L3呈现显著差异外(P<0.05),而其余遮阴处理组间无显著差异(P>0.05)。
2.1.2 对幼苗生长特征的影响
光照显著影响广西火桐幼苗的地上部分生长特征(图2)。株高和基径是反映植物生长发育的重要指标。遮阴处理组的广西火桐幼苗的株高和基径均显著低于CK(P<0.05)。在其他遮阴处理组中,株高和基径的变化趋势均为L1>L2>L3,其中L2的株高显著低于L1(P<0.05),但与L3无显著差异(P>0.05);在基径方面,L2与L1、L3无显著差异(P>0.05),而L1显著大于L3(P<0.05)。

图2 不同光照条件对广西火桐幼苗地上部分的影响
另外,光照也对叶片数量、叶形状和平均叶面积产生显著影响。随着光照的减弱,幼苗的叶片数显著减少(P<0.05),其中L3的幼苗仅有1片叶。遮阴处理组的平均叶面积显著小于CK(P<0.05),且随着光照的减弱而逐渐减小,其中L1显著大于L3(P<0.05),但L2与L1、L3间均无显著差异(P>0.05)。然而,随着光照的减弱,幼苗叶片长宽比逐渐增大,且CK显著小于遮阴处理组(P<0.05);L3的叶片长宽比与L2无显著差异(P>0.05),但两者均显著大于L1处理组(P<0.05)。
2.1.3 对根部特征的影响
由图3可知,广西火桐幼苗的主根长和侧根数(长度>5 cm)均随着光照减弱而减小。其中,CK的主根长和侧根数均显著大于遮阴处理组(P<0.05);而在遮阴处理组内,L3与L2的主根长和侧根数无显著差异(P>0.05),但显著小于L1(P<0.05)。虽然L2的主根长与L1无显著差异(P>0.05),但其侧根数显著小于L1(P<0.05)。
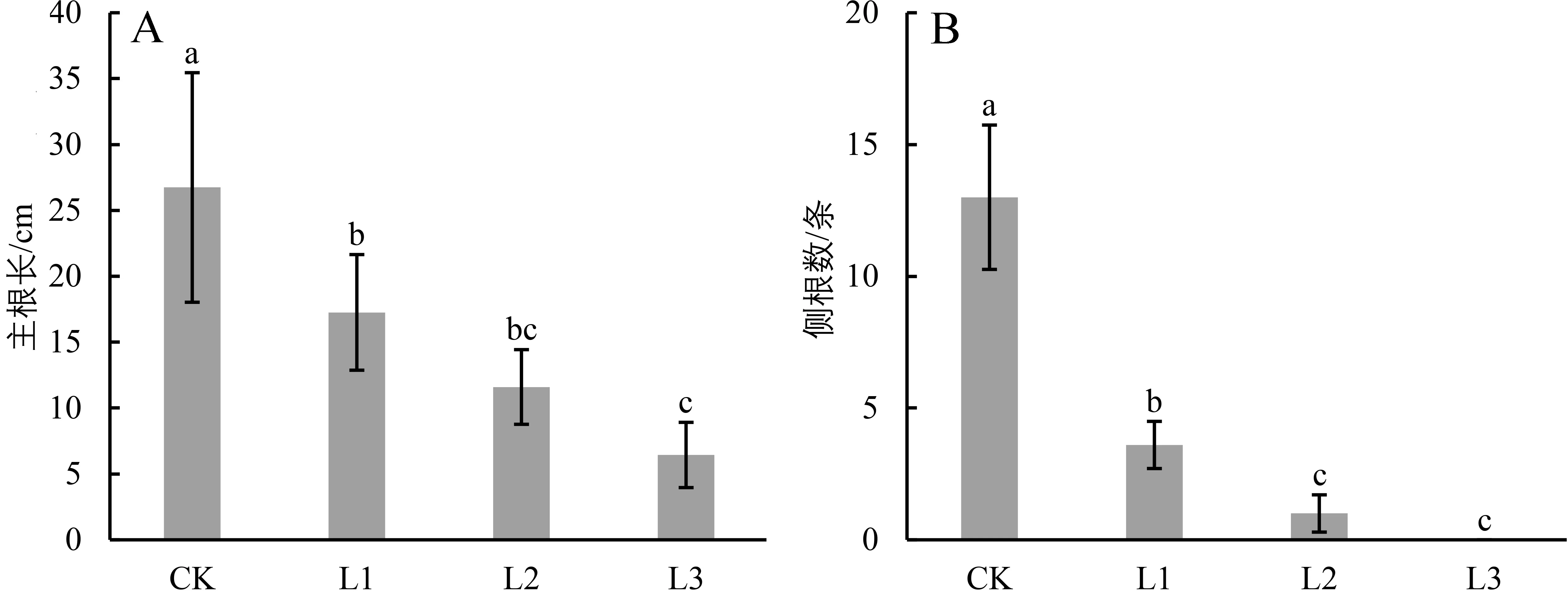
图3 不同光照条件对广西火桐幼苗根部特征的影响
2.2 不同光照条件对广西火桐幼苗生理的影响
2.2.1 对叶绿素含量的影响
广西火桐幼苗的叶绿素含量随光照的减弱呈现先上升后下降的趋势(图4)。其中L1的叶绿素含量最高,为(77.88±3.69)μmol/m2,显著高于CK(110.07%,P<0.05)和L3处理组(110.60%,P<0.05),而与L2无显著差异(P>0.05);其余组叶绿素含量变化则表现为L2>CK>L3,且三者无显著差异(P>0.05)。
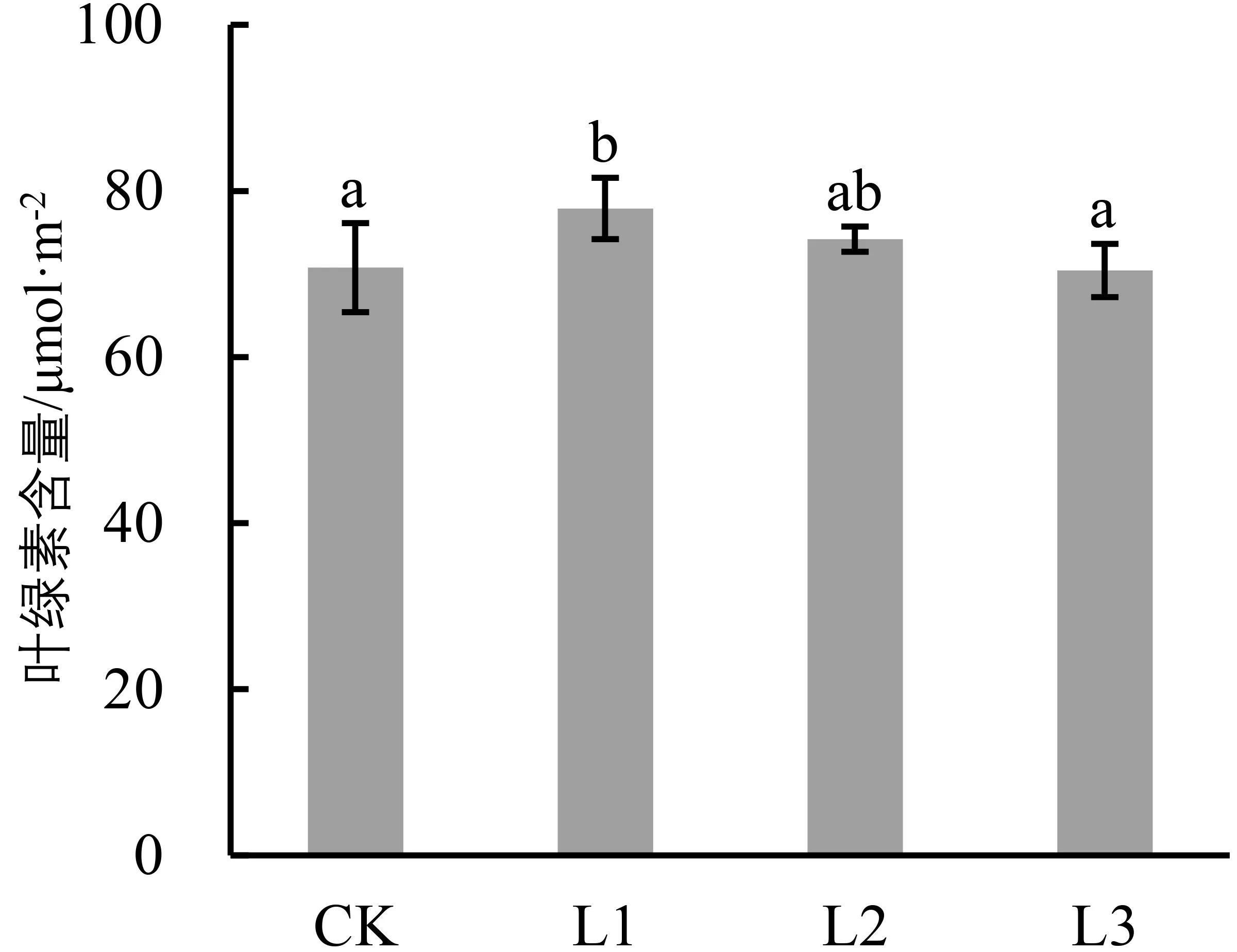
图4 不同光照条件对广西火桐幼苗叶绿素含量的影响
2.2.2 对叶片丙二醛积累的影响
由图5可知,CK的广西火桐幼苗叶片丙二醛(MDA)含量最低,为(11.23±0.24)nmol/g,显著低于各遮阴处理组(P<0.05)。在遮阴处理组中,L1和L2的MDA含量分别为(25.67±2.37)nmol/g和(24.06±3.23)nmol/g,且两者间无显著差异(P>0.05)。L3的MDA含量最大,显著高于L1和L2(P<0.05),为(35.53±1.48)nmol/g。表明在弱光环境下,广西火桐幼苗受到一定的胁迫作用。
2.2.3 对抗氧化酶活性的影响
广西火桐幼苗叶片超氧化物歧化酶(SOD)活性随光照减弱而逐渐升高(图6)。其中,以L3的SOD活性最高,为(694.68±62.89)U/g,显著高于CK(222.75%,P<0.05)、L1(133.29%,P<0.05)和L2(133.63%,P<0.05)。另外,L1和L2的SOD活性较CK分别增加67.12%(P<0.05)和66.69%(P<0.05)。
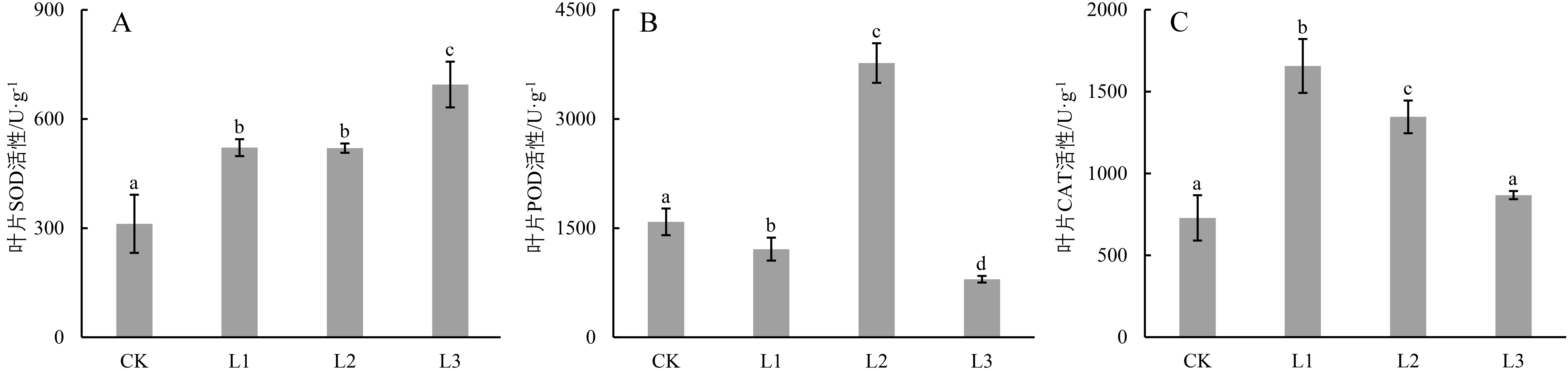
图6 不同光照条件对广西火桐幼苗抗氧化酶活性的影响
随着光照的减弱,过氧化物酶(POD)活性呈“下降—上升—下降”的趋势,以L3的POD活性最低,为(797.58±45.41)U/g,较CK、L1和L2处理组分别下降了49.71%、34.17%和78.84%(P<0.05)。另外,L2的POD活性上升急剧,较CK上升了2.37倍(P<0.05);而L1处理组显著低于CK,下降了23.61%(P<0.05)。
过氧化氢酶(CAT)活性随着光照的减弱呈现先升高后下降的趋势。其中以L1活性最大,为(1 656.40±164.66)U/g,显著大于L2(23.12%,P<0.05)、L3(90.92%,P<0.05)和CK(127.40%,P<0.05);L2次之,显著高于L3(55.06%,P<0.05)和CK(84.69%,P<0.05);L3较CK有所上升,但两者无显著差异(P>0.05)。
2.2.4 对叶片可溶性物质积累的影响
光照对广西火桐幼苗的可溶性物质积累有显著的影响,见图7。
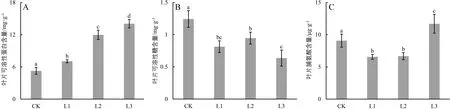
图7 不同光照条件下对广西火桐幼苗的可溶性物质含量的影响
由图7可知,随着光照的减弱,可溶性蛋白质、可溶性糖、游离脯氨酸含量呈现不同变化。可溶性蛋白随着光照的减弱而显著升高,与CK相比,L1、L2和L3的可溶性蛋白含量分别升高了34.14%(P<0.05)、127.55%(P<0.05)和167.07%(P<0.05),且各遮阴组间也存在显著差异(P<0.05)。遮阴处理组的叶片可溶性糖含量均显著低于CK(P<0.05),其中以L3降幅最大,达到48.79%(P<0.05);L1组次之(34.59%,P<0.05);而L2降幅较小(23.86%,P<0.05)。另外,游离脯氨酸含量随着光照的减弱呈先下降后上升的趋势,其中L1和L2的游离脯氨酸含量均显著低于CK,降幅分别为27.78%(P<0.05)和26.57%(P<0.05)。而L3显著高于CK,增幅为28.84%(P<0.05)。
3 讨论与结论
3.1 讨论
自然条件下,植物的生长发育过程受到诸多的环境因子的影响。有研究表明,光照是影响林下幼苗幼树生长发育和繁殖的重要环境因子[16,28-29]。然而,过强或过弱的光照环境均会影响植物正常的光合作用,进而影响其株高、地径和生物量的积累等,导致植物生长量下降[30-31]。本研究结果显示,广西火桐幼苗遮阴处理组的株高、基径、根系发达程度、总生物量、地上部分和根部的生物量均显著低于对照组,且随着光照的减弱呈逐渐减小的趋势,表明广西火桐幼苗期的生长需较充足的光照。另外,光照对广西火桐幼苗的生物量分配影响较小,进一步证明“异速分配”理论,即植物生物量在各器官间的分配与环境变化无关,是仅与植物大小有关的幂函数。这与Liu等[32]对蒿属(Artemisia)植物的研究结果一致。
植物叶片是响应环境变化的敏感器官[33-34]。在弱光胁迫下,植物通过增加叶片面积以获取更多的光资源[35-36]。然而,本研究结果显示,广西火桐幼苗的叶片数量和平均叶面积均随着光照减弱而逐渐减少,且遮阴处理组显著低于对照组,进一步表明光照不足对广西火桐幼苗的生长产生抑制作用,这与王虹妍等[37]对米奴塔睡莲(Nymphaeaminuta)的研究结果一致。本研究还发现广西火桐幼苗的叶片长宽比随光照的减弱而显著升高,说明在弱光下,广西火桐幼苗改变叶片形状,使得其变得“细长”,这与闫旭等[24]复层林对闽楠幼树叶片形态影响的研究结果相似。
光胁迫还会引起植物细胞内氧自由基的快速积累,细胞膜损伤加剧,导致丙二醛(MDA)大量积累;而MDA会破坏细胞膜的稳定性,其含量变化可反映植株细胞质膜受胁迫的程度[38]。在本研究中,遮阴条件下的广西火桐幼苗MDA含量均显著高于对照组,且重度遮阴下的MDA表现最高,表明弱光环境加剧细胞膜的氧化分解,导致MDA大量积累,幼苗的细胞质膜受到显著胁迫。
通常植物在受到逆境的胁迫后,往往会通过升高可溶性蛋白、可溶性糖和游离脯氨酸含量,以稳定其细胞膜完整性,进而缓解胁迫所带来的损伤[39]。本研究结果发现,遮阴处理组的可溶性糖含量显著低于对照组,说明遮阴已引起广西火桐幼苗光合效率不足,导致其碳水化合物的积累下降。游离脯氨酸作为植物细胞中主要渗透调节物质之一,在逆境中,植物细胞内游离氨基酸含量升高有助于维持膜结构稳定和细胞渗透压平衡[40]。重度遮阴条件下的广西火桐幼苗游离脯氨酸含量最高,而其他遮阴处理组却显著低于对照组。本研究发现广西火桐幼苗的可溶性蛋白随光照的减弱而显著升高,在重度遮阴下含量达到最大,且幼苗在此条件下生长受到严重抑制,由此说明可溶性蛋白是广西火桐幼苗应对弱光胁迫的主要渗透调节物。因此,在光照严重不足的环境下,可溶性蛋白和游离脯氨酸含量上升可增加广西火桐幼苗细胞渗透势,进一步维持其细胞膜的结构。
此外,SOD、POD和CAT活性可以有效清除细胞内的自由基和过氧化物,提高植物抗逆性[41-42]。本研究结果表明,SOD、POD和CAT没有表现出一致性的变化,SOD活性随着遮阴程度的升高而升高,在重度遮阴处理组中其活性达到最大,而POD和CAT最大活性分别出现在中度遮阴和轻度遮阴处理组。这说明不同的酶对于光照强度的敏感性和适应性不同,但三者共同协助幼苗抵御不良的环境。重度遮阴条件下SOD活性最高、POD和CAT活性最低,表明该条件下广西火桐幼苗不能够及时清除细胞内产生的过氧化物和过氧化氢,导致膜结构进一步被破坏,MDA加速积累。结合广西火桐幼苗的生长性状,说明重度遮阴极不利于广西火桐幼苗的生长。
综合上述分析可发现,光照是影响广西火桐幼苗生长发育的重要环境因素。当幼苗处于光照不足的环境时,表现出相应受胁迫状况:一方面表现为生长受到抑制,光合效率下降,株高、基径、叶片数量、叶面积和生物量积累等生长指标下降;另一方面表现为细胞内过氧化作用加剧,MDA大量积累,渗透压平衡受到破坏。为应对弱光胁迫带来的损伤,幼苗细胞内的可溶性蛋白和抗氧化物酶活性均呈现不同程度升高。表明广西火桐幼苗耐阴能力较差,环境中的光照越弱,其生长受到的抑制就越严重。
3.2 结论
广西火桐为强阳性树种,其幼苗生长需较充足的光照,弱光会明显抑制生长,因而光照是影响广西火桐野生种群幼苗更新的重要环境因子。林下郁闭程度较高的广西火桐天然群落中,种子虽能大量萌发但幼苗保存率却极低,这与处于下层的幼苗获得光能较少密切相关。因此,在进行广西火桐种苗繁育时,除加强肥水管理外,适当增加透光率有助于苗木的生长;在开展广西火桐野生种群恢复时,通过人工干预减少林内郁闭度以增加林下透光率,促进幼苗幼树的生长,有助于种群复壮;在开展广西火桐野外回归时,宜选择无遮阴或林缘等生境。
