麻牛膝TONSOKU基因克隆、生信分析及其抑菌作用
2023-08-25罗新何梦竹陈曦万健张浪李瑞陈稷田孟良
罗新?何梦竹?陈曦?万健?张浪?李瑞?陈稷?田孟良
摘要:目的 克隆編码麻牛膝(Cyathula capitata(Wall.) Moq.)CcTSK蛋白的基因全长,通过基因重组技术制备表达质粒,对表达产物进行生物学预测和抑菌分析。方法 以麻牛膝嫩叶为材料,完成CcTSK基因cDNA的克隆。采用生信软件分析该基因及编码蛋白的生物学特征。构建CcTSK基因原核表达载体,并转入大肠埃希菌BL21(DE3)感受态中,IPTG诱导表达,考察CcTSK-GST融合蛋白的表达菌株对低浓度抗生素的敏感性、自身生长变化及在较高浓度抗生素下的存活率。结果 CcTSK基因的gDNA为12613 bp,CDS为3855 bp,共编码1284个氨基酸。CcTSK蛋白为亲水性不稳定蛋白,没有跨膜结构域,属于没有信号识别功能的非分泌蛋白,亚细胞定位预测该蛋白可能位于细胞核,并且具有核定位信号,含有TPR重复序列与LRR重复序列结构域,二级结构主要有α-螺旋(占53.89%)、无规卷曲(占32.79%)、延伸链(占8.88%)和β-转角(占4.44%)。经IPTG诱导后,CcTSK-GST融合蛋白的表达抑制大肠埃希菌生长。低浓度抗生素测试结果显示,CcTSK-GST表达菌株对氯霉素(chloramphenicol, Cam)最敏感;高浓度抗生素处理下,CcTSK-GST的表达使得抑菌现象显著增强。结论 成功获得编码CcTSK蛋白的基因全长及原核表达质粒,并对其生物学特征进行了分析比较,该蛋白与拟南芥AtTSK蛋白有较高相似性。此外,CcTSK-GST融合蛋白的表达对大肠埃希菌增殖具有抑制作用,并且显著增强抗生素所产生的抑菌现象。
关键词:麻牛膝;TONSOKU;基因克隆;生物信息学;抑菌;抗生素;增强效应
中图分类号:R285文献标志码:A
Cloning, bioassay, and antibacterial activity of TONSOKU gene from Cyathula capitata (Wall.) Moq.
Luo Xin, He Meng-zhu, Chen Xi, Wan Jian, Zhang Lang, Li Rui, Chen Ji, and Tian Meng-liang
(College of Agronomy, Sichuan Agricultural University, Chengdu 611130)
Abstract Objective In order to predict the biological and antimicrobial analysis of the expressed product, the full length of the gene encoding CcTSK protein of Cyathula capitata (Wall.) Moq. was cloned and the expression plasmid was prepared by gene recombination. Methods The cDNA of CcTSK gene was cloned from tender leaves of Cyathula capitata (Wall.) Moq. Bioinformatics software was used to analyze the biological characteristics of the gene and its encoding protein. The prokaryotic expression vector of CcTSK gene was constructed and transferred into E. coli BL21(DE3) competent strain, and induced by isopropyl β-D-thiogalactopyranoside (IPTG), to investigate the sensitivity of strains expressing CcTSK-GST fusion protein to low concentration of antibiotics, and their own growth and cell survival rate under high concentration of antibiotics. Results The gDNA and CDS of CcTSK gene were 12613 bp and 3,855 bp, encoding 1,284 amino acids. CcTSK protein is a hydrophilic unstable protein with no transmembrane domain and belongs to a non-secretory protein lacking the ability to recognize signals. Subcellular localization predicts that it may be located in the nucleus and has nuclear localization signal, as well as two domains containing TPR repeats and LRR repeats. The main secondary structures were α -helix (53.89%), random coil (32.79%), extended chain (8.88%), and β-rotation (4.44%). Induced by IPTG, expression of CcTSK-GST fusion protein inhibited the growth of E. coli. The results of low concentration antibiotic test showed that CcTSK-GST expressing strain was the most sensitive to chloramphenicol (Cam), and the antibacterial effect was significantly enhanced at higher concentration of antibiotic. Conclusions The full length of the coding gene and prokaryotic expression plasmid of CcTSK protein were successfully obtained, and the biological characteristics of the protein were analyzed and compared. The protein had high similarity with the TSK protein of Arabidopsis(AtTSK). In addition, the expression of CcTSK-GST fusion protein could inhibit the proliferation of E. coli, and significantly enhanced the bacteriostatic phenomenon produced by antibiotics.
Key words Cyathula capitata (Wall.) Moq.; TONSOKU; Gene cloning; Bioinformatics; Bacteriostatic; Antibiotic; Enhancement effect
在药用植物川牛膝(Cyathula officinalis Kuan.)及其伪品麻牛膝的分子鉴定中,田孟良等[1]基于RAPD标记筛选出分子标记SCAR_L2,利用川牛膝全基因测序技术将该分子标记定位到了TONSOKU基因。但在川牛膝与麻牛膝中,TONSOKU基因产生差异的原因,以及该基因对牛膝功能的影响尚未可知。
麻牛膝为苋科植物头花杯苋,目前有关研究主要围绕如何鉴定麻牛膝与川牛膝,包括形态鉴定[2-4]、分子鉴定[1,5]、化学成分和药理作用的比较[6-10]。在药理作用比较中发现,麻牛膝根部水煎药液具有更强的小鼠急性毒性、免疫抑制作用和抗炎作用[8-9],且麻牛膝多糖能够显著降低猪小肠上皮细胞炎症因子IL-6的表达[10]。麻牛膝更显著的抗炎作用为其成为区别于川牛膝的中药材提供了可能性。
而TONSOKU基因最初是为研究拟南芥顶端分生组织功能而被鉴定的[11]。TONSOKU不同位置突变造成的植株表型不同,故根据表型差异对该基因的突变体分别命名为MGOUN3[11] 、TONSOKU[12]、BRUSHY1[13],其中TONSOKU较为常用。目前,关于TONSOKU的研究主要以模式植物拟南芥(Arabidopsis thaliana)和烟草(Nicotiana tabacum)为材料,并关注于该基因对植物生长发育本身的调控作用[11-15]。
本实验以麻牛膝TONSOKU基因为研究对象,克隆编码麻牛膝CcTSK蛋白的基因全长,对该基因及所编码蛋白质的生物学特征进行分析,为川、麻牛膝基于RAPD标记的SCAR分子标记技术鉴定方法提供进一步参考。另外与川牛膝相比,麻牛膝具备更显著的抗炎作用[9],本实验通过抗生素敏感性实验及细胞生长实验,观察CcTSK蛋白表达而表现的抑菌性,为该基因的功能研究提供新参考。
1 材料与方法
1.1 材料
1.1.1 试验材料
本实验所用麻牛膝植株采自四川省雅安市宝兴县蜂桶寨乡(北纬30°东经102°)中药材种植基地(海拔约1400 m),经四川农业大学田孟良教授鉴定为苋科麻牛膝(Cyathula capitata(Wall.) Moq.)[16]。
表达载体pGEX-6t-1由四川农业大学农学院提供;大肠埃希菌BL21(DE3)菌株购自博迈德生物公司。
1.1.2 主要试剂盒及设备
EASYspin Plus Plant RNA Kit(北京艾德莱生物科技有限公司)、HiScript? Ⅲ RT SuperMix for qPCR (+gDNA wiper)(南京諾唯赞生物科技股份有限公司)、FastPure? Gel DNA Extraction Mini Kit(南京诺唯赞生物科技股份有限公司)、SMARTer@RACE5/3 Kit(日本TaKaRa公司)、One Step Cloning Kit(南京诺唯赞生物科技股份有限公司)。
超微量浓度测定仪(德国Implen公司)、恒温培养箱(郑州生元仪器有限公司)、恒温振荡器(江苏太仓市实验设备厂)、低温离心机(大龙兴创实验仪器股份公司)。
1.2 gDNA、RNA的提取与cDNA的合成
称取麻牛膝健康幼嫩叶片0.1 g,使用CTAB法提取植物gDNA。再次称取0.1 g叶片并使用试剂盒提取植物总RNA,经浓度测定及琼脂糖凝胶电泳检测后,以RNA为模板合成cDNA。
1.3 麻牛膝CcTSK基因克隆和测序
依据川牛膝基因组从头测序组装拼接的川牛膝TSK基因(CoTSK)作为参考序列[17],进行引物设计,对cDNA序列扩增,并进行胶回收,经酶切检测后,将目的片段送往擎科生物公司测序获得核心序列。以核心序列为模板,使用试剂盒进行cDNA末端快速扩增克隆,并通过胶回收、测序、序列拼接获得该基因完整cDNA序列。设计gDNA特异引物对,进行PCR反应扩增,再进行胶回收及测序,通过序列拼接得到完整gDNA序列。引物设计软件为primer 3.0和DNAMAN(表1)[16]。
1.4 麻牛膝CcTSK基因的生物信息学分析
拼接后的基因序列开放阅读框由ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)完成,氨基酸序列由ExPASy(http://web.expasy.org/translate/)完成。信号肽分析由SignalP-6.0(https://biolib.com/DTU/SignalP-6/)网站完成。蛋白的亲水性与疏水性由ProtScale(https://web.expasy.org/protscale/)完成,并通过ProtParam(http://web.expasy.org/cgi-bin/protparam/protparam)在线预测其理化性质。利用SMART网站(http://smart.embl-heidelberg.de/)预测CcTSK蛋白结构域,跨膜区域由TMHMM-2.0(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)完成。CcTSK蛋白亚细胞定位预测通过WoLF PSORT Prediction(https://wolfpsort.hgc.jp/),SWISS-MODEL(https://swissmodel.expasy.org/)进行三级结构同源建模,核定位信号通过cNLS Mapper(https://nls-mapper.iab.keio.ac.jp/cgi-bin/NLS_Mapper_form.cgi),蛋白二级结构分析通过SOPMA(https://npsa-prabi.ibcp.fr/NPSA/npsa_sopma.html)。拟南芥AtTSK蛋白信息查询网站为UniProt(https://www.uniprot.org/)。
1.5 目的菌株构建
以pGEX6t(F)-BamHI-TSK和pGEX6t(R)-XhoI-TSK为引物,扩增麻牛膝CcTSK基因CDS序列,通过引物序列,使得扩增产物两端连上载体BamHI和XhoI酶切位点的碱基序列,再通过同源重组的方法,将其胶回收产物构建到用BamHI和XhoI过夜酶切后的pGEX-6t-1载体上。连接产物及空载通过热激法转化到大肠埃希菌BL21(DE3)感受态菌株中,挑选单克隆,用CcTSK基因特异引物3602F和H4389R进行PCR验证,验证成功后进行扩大培养,转入含有50 μg/mL氨苄西林(ampicillin sodium salt, Amp)的LB液体培养基中,37℃ 200 r/min过夜培养。挑选阳性菌落提取质粒后进行酶切验证,选择验证正确的片段送测序,将测序比对无误的菌株进行保存[16]。
1.6 蛋白表达
将成功转入重组载体的BL21(DE3)菌株培养至A600=0.5,加入0.1 mmol/L IPTG,37℃摇床诱导5 h。取1 mL菌液,加入15 μL三氯乙酸(trichloroacetic acid,TCA),混匀后液氮冻融2~3次(液氮与37℃热水浴交替),再加入500 mL预冷的蛋白提取液(含有10% TCA与0.07% β-巯基乙醇的丙酮溶液),进行震荡悬浮,于-20℃静置过夜。4℃,12000 r/min离心20 min,向沉淀中加入1 mL含0.07% β-巯基乙醇的丙酮溶液,混匀后于-20℃静置30 min,12000 r/min,离心20 min并干燥,最后加入50 μL蛋白裂解液(8 mmol/L尿素,0.07% β-巯基乙醇),摇匀加入15 μL十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-Polyacrylamide gel electrophoresis,SDS-PAGE)缓冲液煮沸10 min,4℃冷却上样并进行电泳。将电泳后的凝胶在考马斯亮蓝溶液中染色5 h,转至蛋白洗脱液中脱色,条带清晰后在凝胶电泳成像仪中照胶记录[16]。
1.7 抗生素敏感性分析
选取0.5 μg/mL低浓度硫酸卡那霉素(kanamycin sulfate, Kan)、硫酸庆大霉素(gentamicin sulfate, Gen)、硫酸链霉素(streptomycin sulfate, Str)、利福平(rifampicin, Rif)和Cam进行试验。将转入pGEX-vector质粒的BL21菌株(对照菌株)与转入pGEX-CcTSK质粒的BL21菌株(过表达菌株)37℃摇床200 r/min培养至A600=0.5,分别加入终浓度为0.1 mmol/L的IPTG,诱导3 h后稀釋,将稀释倍数为10-3、10-4、10-5和10-6的菌液在无抗生素和添加不同种类抗生素的平板上进行点板,并以无IPTG诱导的两种菌株作为对照,37℃培养15 h后观察各组对抗生素的敏感性。
根据文献[18-20],设置浓度为0、0.025、0.05、0.075和0.1 mmol/L的IPTG进行细菌生长试验。将对照菌株与过表达菌株培养至A600=0.5后分别稀释,设置稀释倍数10-1~ 10-7的7个梯度,各梯度取2 μL菌液在含有不同浓度IPTG平板上点板,37℃培养12 h后,观察菌株的生长情况。
选取50 μg/mL较高浓度头孢噻肟钠(cefotaxime sodium salt, Cef)、Kan、Str、Rif、Gen和Cam进行试验。将对照菌株与过表达菌株培养至A600=0.5后,加入终浓度为0.1 mmol/L的IPTG诱导3 h,再分别加入不同种类的较高浓度抗生素诱导5 h,然后进行稀释,取稀释倍数为10-3菌液进行涂板,同时以不加抗生素的菌液作为对照,37℃培养13 h后统计菌落数(CFU),并比较不同抗生素处理下的存活率。
2 结果
2.1 TONSOKU基因克隆
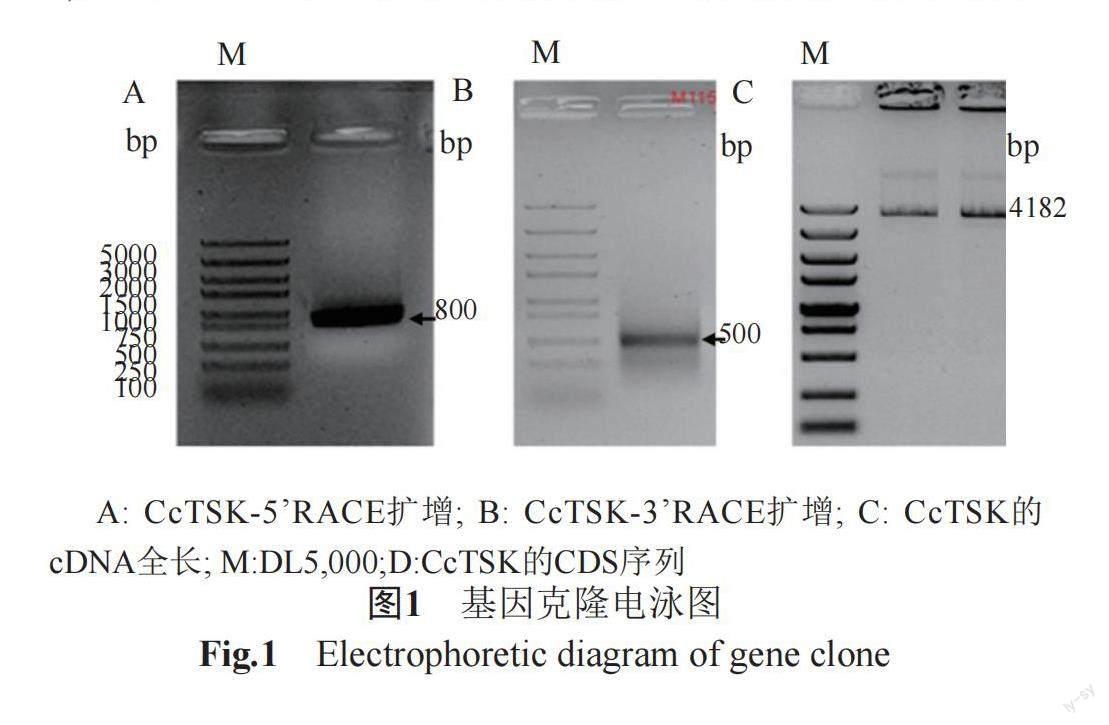
提取麻牛膝总RNA,并反转录合成cDNA。设计引物进行RACE反应,电泳检测5'RACE和3'RACE扩增条带(图1A和B),经测序分析,确定麻牛膝CcTSK基因5'UTR的149 bp序列和3'UTR的178 bp序列。设计特异引物,进行PCR反应扩增。通过琼脂糖电泳、胶回收及生物公司测序,确定麻牛膝CcTSK基因CDS全长3855 bp(图1C),基因登录号为OP858787。提取麻牛膝gDNA,设计引物,进行PCR反应,经琼脂糖电泳、胶回收及生物公司测序,确定麻牛膝CcTSK基因全长12613 bp。
2.2 麻牛膝TONSOKU蛋白的理化性质分析
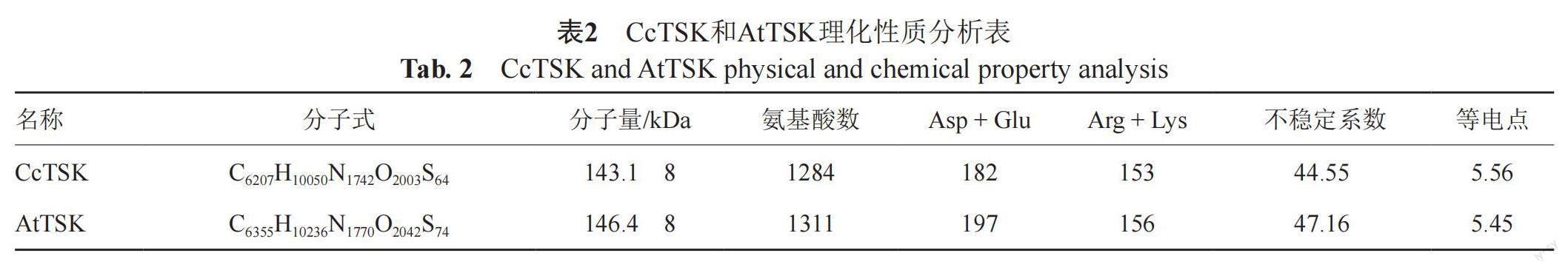
利用ORF Finder查找CcTSK基因的开放阅读框,并通过ExPASy在线工具将其转换为氨基酸序列,在ProtParam软件平台分析蛋白理化性质,预测结果显示CcTSK蛋白(C6207H10050N1742O2003S64)总分子质量为143.18 kDa,含有1284个氨基酸,是不稳定蛋白(表2)。ProtScale在线平台分析其在一级结构上的亲疏水性,结果显示在一级氨基酸结构上CcTSK蛋白表现为亲水性,其中第315位氨基酸分值最低为-3.044,该位置亲水性最强,第1084位氨基酸分值最高为2.522,该位置疏水性最强;AtTSK蛋白同样表现为亲水性,其中第607位氨基酸分值最低为-2.644,该位置亲水性最强,第1140与1142位氨基酸分值最高为2.311,该位置疏水性最强(图2),理化性质上CcTSK与AtTSK蛋白具有较高相似性。
2.3 麻牛膝TONSOKU蛋白跨膜结构域和信号肽预测
在TMHMM-2.0网站预测CcTSK蛋白的跨膜区域,该蛋白没有跨膜区域,为膜外蛋白(图3A)。新合成的多肽链进行跨膜转移或定位常常需要信号肽的指引,使用SignalP-6.0对CcTSK蛋白进行信号肽预测发现该蛋白没有信号肽结构,属于没有信号识别功能的非分泌蛋白(图3B),与AtTSK蛋白一致。
2.4 CcTSK蛋白结构域分析
使用SMART在线网站对CcTSK蛋白进行结构域分析,结果显示CcTSK蛋白主要由2个分别含有TPR重复序列和LRR重复序列的结构域构成,TPR重复序列包含5个TPR基序和2个TPR-like基序,LRR重复序列含有5个LRR基序,这与拟南芥AtTSK蛋白结构具有较高相似性[13](图4)。
2.5 CcTSK蛋白亚细胞定位预测
使用WoLF PSORT Prediction在线网站对CcTSK蛋白的亞细胞定位进行预测,该蛋白定位于细胞核的评分为11,定位于细胞质的评分为3,该蛋白位置最可能在细胞核中。再利用cNLS Mapper对CcTSK蛋白进行NLS预测,结果表明从N端起第595~603个氨基酸(VGRKRSRLV)构成该蛋白核定位信号,该结果评分为5。
2.6 CcTSK蛋白结构分析
利用SOPMA对麻牛膝CcTSK氨基酸序列进行二级结构预测分析。结果表明,麻牛膝CcTSK二级结构主要有β-转角(beta turn 4.44%)、α-螺旋(alpha helix 53.89%)、无规卷曲(random coil 32.79%)和延伸链(extended strand 8.88%)(图5A),与拟南芥AtTSK二级结构构成比例相近[21]。
利用在线软件SWISS-MODEL蛋白质3D结构数据库对CcTSK蛋白质进行三级结构建模,选择编号为4peq.1牛核糖核酸酶抑制因子序列为模板,其QMEAN值为0.17(图5B)。
2.7 过表达菌株与抗生素抑菌作用产生增强效应
与川牛膝相比,麻牛膝具备更显著的抗炎作用[9],因此为了探究CcTSK基因的表达对细菌生长的影响,本试验选择了5种抗生素进行研究。将构建成功的pGEX-CcTSK重组载体转入大肠埃希菌BL21菌株体内,在0.1 mmol/L的IPTG诱导下,通过SDS-PAGE电泳检测分析重组蛋白表达。结果显示,GST蛋白表达的对照菌株成功表达26 kDa的GST标签蛋白,CcTSK-GST融合蛋白的过表达菌株在150 kDa左右出现明显的诱导条带,与CcTSK-GST融合蛋白预测分子量169.18 kDa一致(图6B)。
将成功进行蛋白表达的对照菌株与过表达菌株进行抗生素处理。培养15 h后结果显示,当未添加抗生素时,对照菌株与过表达菌株生长状况没有显著差异。当添加终浓度为0.5 μg/mL抗生素后,与对照菌株相比,在未经IPTG诱导时,过表达菌株生长均未受到显著抑制;在IPTG诱导3 h后,过表达菌株的生长仅在添加氯霉素的平板上受到显著抑制,说明经IPTG诱导后,在长时间低浓度抗生素处理下,CcTSK-GST融合蛋白的表达使得大肠埃希菌对氯霉素更为敏感(图7A)。
此外,通过上述实验可以发现IPTG诱导表达的CcTSK-GST融合蛋白对细菌生长产生了一定影响,为进一步探究其作用机制,在不同浓度的IPTG诱导下分析CcTSK-GST融合蛋白表达与细菌生长的关系。对照菌株在不同浓度IPTG诱导处理下其细菌生长未表现出显著差异,而过表达菌株生长显著受到IPTG浓度的影响,其生长受抑制情况随IPTG浓度增加而加剧,说明CcTSK-GST融合蛋白的表达对大肠埃希菌的生长产生了抑制作用(图7B)。
为了考察CcTSK蛋白的表达对细菌增殖的抑制性,和对抗生素敏感性的内在关联,对不同抗生素处理下,过表达菌株的存活率进行了分析。结果显示,加入终浓度为50 μg/mL抗生素处理5 h后,对照菌株的存活率均低于60%,且除头孢外,过表达菌株的生长相较于对照菌株均表现出更明显的抑制,说明CcTSK-GST融合蛋白的表达与较高浓度抗生素的叠加处理,使得抑菌现象更加显著(图7C)。
3 讨论
TONSOKU基因是为研究顶端分生组织功能而从分生异常的拟南芥突变体中分离出来[11],该基因主要在模式植物拟南芥和烟草中进行研究。在生信分析中发现,麻牛膝CcTSK蛋白与拟南芥AtTSK蛋白具有较高相似性,并且二者具有相同TPR和LRR保守结构域,推测二者具有相似的基因功能,现有研究已知拟南芥AtTSK蛋白在维持植物正常形态建成[14]、促进DNA损伤修复、细胞有丝分裂调控[21]、维持转录水平基因沉默[22]以及维护染色体结构[23]中发挥作用。最新研究发现,植物TONSOKU蛋白能够通过TPR结构域特异结合组蛋白H3.1变体,维护DNA复制过程中基因组的稳定性,并且这种特异性结合在动物同源TONSL蛋白中也是保守的[24],使得对TPR结构域的功能有了进一步发现。亚细胞定位显示CcTSK蛋白位于细胞核,AtTSK的分析中同样预测到了核定位信号肽,并在烟草细胞核中检测到了AtTSK-GFP信号,证实了这一预测[13]。此外本研究发现IPTG诱导CcTSK蛋白表达后,对大肠埃希菌生长产生了抑制作用,而对照菌株并没有明显影响,这可能是因为CcTSK蛋白同样具有维持转录水平基因沉默的功能,过表达后抑制了大肠埃希菌体内一些正常蛋白的表达,从而阻碍了细菌生长。另外通过抗生素敏感性实验发现,低浓度抗生素和IPTG对对照菌株并没有明显影响,而对过表达菌株来说,仅当氯霉素和IPTG同时加入时,大肠埃希菌生长受抑制现象加剧。王友如[25]、张海峰等[26]和陈绪美等[27]提出氯霉素能够扩增大肠埃希菌质粒,在低浓度氯霉素作用下,CcTSK基因得到扩增,并被IPTG诱导表达,从而阻碍了细菌生长。最后在较高浓度抗生素处理下,通过对菌株存活率的检测发现,CcTSK-GST融合蛋白的表达与较高浓度抗生素的叠加处理,使得抑菌现象更加显著,进一步提供了CcTSK蛋白在抑菌功能上的可能性,说明麻牛膝除多糖等活性成分,还可能通过抑菌方式帮助抗炎,同时这也补充了植物TONSOKU同源基因的生物学功能研究。
4 结论
本研究从麻牛膝嫩叶中成功克隆得到CcTSK的gDNA和CDS序列,并对其进行生物信息学分析。CcTSK基因的gDNA为12613 bp,CDS为3855 bp,共编码1284个氨基酸。CcTSK蛋白为亲水性不稳定蛋白,没有跨膜结构域,属于没有信号识别功能的非分泌蛋白,亚细胞定位预测该蛋白最可能处于核内,并且具有核定位信号。该蛋白含有TPR重复序列和LRR重复序列的两个结构域,二级结构主要有α-螺旋(占53.89%)、无规卷曲(占32.79%)、延伸链(占8.88%)和β-转角(占4.44%),生物学特征比较分析发现,CcTSK蛋白与AtTSK蛋白具有较高相似性。此外,通过抗生素敏感性实验发现,在低浓度抗生素处理下,CcTSK基因过表达后对氯霉素最敏感。而在细胞生长实验中发现,CcTSK-GST融合蛋白的表达对大肠埃希菌的生长产生了抑制作用,且随IPTG浓度的增加,抑制现象越明显。另外,当CcTSK-GST融合蛋白的表达与较高浓度抗生素叠加处理时,抑菌现象更加显著,这为CcTSK蛋白在抑菌功能上的研究提供了参考,但CcTSK蛋白与抗生素的抑菌作用之间的关联机制,还有待进一步研究确定。
参 考 文 献
田孟良, 官宇, 刘帆, 等. 基于RAPD标记的SCAR分子标记技术鉴定川牛膝及其混淆品[J]. 中国中药杂志, 2010, 35(8): 953-956.
梁欢, 何梦竹, 文青松, 等. 川牛膝与麻牛膝的形态鉴别[J]. 中国药学杂志, 2020, 55(5): 342-348.
趙磊. 川牛膝及其常见混用品鉴别研究[J]. 亚太传统医药. 2017, 13(16): 32-34.
吴向莉. 川牛膝与麻牛膝的鉴别[J]. 贵阳中医学院学报, 2000, 22(1): 62-64.
陈诗晴, 蒲沁琳, 陈小军, 等. 基于ITS序列分析技术鉴定川牛膝与常见伪品麻牛膝[J]. 中药与临床, 2014, 5(4): 4-6.
童凯, 李昭玲, 闫燊, 等. 基于HPLC指纹图谱比较分析川牛膝及其混淆品的化学成分特征[J]. 时珍国医国药, 2015, 26(10): 2424-2427.
高必兴, 齐景梁, 耿昭, 等. 基于皂苷类成分研究鉴别川牛膝及其伪品[J]. 中药材, 2020, 43(9): 2118-2123.
何光星, 舒光明, 宾雪英, 等. 川牛膝及其混淆品活血化瘀作用比较[J]. 时珍国医国药, 2015, 26(10): 2336-2339.
宾雪英, 舒光明, 何光星, 等. 川牛膝及其混淆品的抗炎作用比较[J]. 资源开发与市场, 2013, 29(4): 341-384.
赵磊, 冯鑫, 陈容, 等. 川牛膝、麻牛膝及杂交牛膝多糖的分离、纯化和对猪小肠上皮细胞活性的比较研究[J]. 饲料工业, 2019, 40(8): 31-36.
GuyomarcH S, Vernoux T, Traas J, et al. MGOUN3, an Arabidopsis gene with tetratrico peptide-repeat-related motifs, regulates meristem cellular organization[J]. J Exp Bot, 2004, 55(397): 673-684.
Suzuki T, Inagaki S, Nakajima S, et al. A novel Arabidopsis gene TONSOKU is required for proper cell arrangement in root and shoot apical meristems[J]. Plant J, 2004, 38(4): 673-684.
Takeda S, Tadele Z, Hofmann I, et al. BRU1, a novel link between responses to DNA damage and epigenetic gene silencing in Arabidopsis[J]. Genes Dev, 2004, 18(7): 782-793.
GuyomarcH S, Benhamed M, Lemonnier G, et al. MGOUN3: Evidence for chromatin-mediated regulation of FLC expression[J]. J Exp Bot, 2006, 57: 2111-2119.
Suzuki T, Nakajima S, Morikami A, et al. An Arabidopsis protein with a novel calcium-binding repeat sequence interacts with TONSOKU/MGOUN3/BRUSHY1 involved in meristem maintenance[J]. Plant Cell Physiol, 2005, 46(9): 1452-1461.
何梦竹. 川牛膝形态鉴定与TSK基因功能初步研究[D]. 成都: 四川农业大学, 2018.
闫燊, 池旭, 童凯, 等. 中药植物川牛膝基因组从头测序及分析[J]. 中国农业科技导报, 2016, 18(3): 32-37.
Feng B Y, Mandava C S, Guo Q, et al. Structural and functional insights into the mode of action of a universally conserved Obg GTPase[J]. Plos Biol, 2014, 12(5): 866-880.
Kobayashi G, Moriya S, Wada C. Deficiency of essential GTP-binding protein ObgE in Escherichia coli inhibits chromosome partition[J]. Mol Microbiol, 2001, 41(5): 1037-1051.
Chen J, Deng F, Deng M S, et al. Identification and characterization of a chloroplast-targeted Obg GTPase in Dendrobium officinale[J]. DNA Cell Biol, 2016, 35(12): 802-811.
何夢竹, 黄进, 杜丽娥, 等. 拟南芥TONSOKU基因研究进展[J]. 分子植物育种, 2019, 17(4): 1175-1185.
Roudier F, Ahmed I, Bérard C, et al. Integrative epigenomic mapping defines four main chromatin states in Arabidopsis[J]. EMBO J, 2011, 30: 1928-1938.
Schubert I, Shaw P. Organization and dynamics of plant interphasechromosomes[J]. Trends Plant Sci, 2011, 16: 273-281.
Davarinejad H, Huang Y, Mermaz B, et al. The histone H3.1 variant regulates TONSOKU-mediated DNA repair during replication[J]. Science, 2022, 375: 1281-1286.
王友如. 大肠杆菌质粒DNA提取方法的优化[J]. 生物技术, 2006, 16(5): 42-44.
张海峰, 张登波, 马大龙. 氯霉素对大肠杆菌质粒的扩增作用[J]. 首都医学院学报, 1988(4): 295.
陈绪美, 尹志勇, 檀军, 等. 不同浓度氯霉素对大肠杆菌质粒扩增的影响[J]. 山地农业生物学报, 2020, 39(6): 81-84.
