佛手散改善谷氨酸钠诱导的PC12细胞氧化应激损伤的作用机制▲
2023-08-25冯丽旋郭学军张永斌
冯丽旋 郭学军 张永斌 陈 苑
(广州中医药大学实验动物中心,广东省广州市 510405)
阿尔茨海默病(Alzheimer′s disease,AD)是一种神经退行性疾病,可引起一系列的神经病变,导致记忆、认知、行为异常,甚至引发痴呆[1-2]。谷氨酸作为一种兴奋性神经递质,细胞外谷氨酸水平过高可导致神经元死亡和神经退行性疾病。谷氨酸诱导的兴奋性毒性一方面可通过释放大量的谷氨酸和Ca2+内流实现,另一方面也可通过氧化应激介导[3]。细胞外过量的谷氨酸受体可增加细胞内Ca2+浓度,高浓度的细胞内Ca2+可诱导活性氧簇大量产生,导致线粒体凋亡[4-6]。此外,胱氨酸是谷胱甘肽的前体,而细胞外的谷氨酸盐可通过抑制细胞对胱氨酸的吸收,导致细胞内谷胱甘肽水平下降,诱导细胞发生氧化应激并导致细胞死亡[3,7]。因此本研究推测氧化应激是谷氨酸诱导细胞凋亡的主要原因。
随着传统中医药在顽固性疾病治疗中的应用日益增多[8-10],中医药治疗AD的研究也得到快速发展,其中由当归和川芎组成的佛手散已被广泛应用于神经退行性疾病的治疗[11-14]。体外实验显示,佛手散中的当归、川芎可通过协同作用发挥防治帕金森病的作用[15]。佛手散水煎剂可改善实验性脑缺血所致的大鼠神经行为学障碍、脑水肿程度及脑梗死体积[16]。此外,佛手散可通过调节肠-肝-脑轴来改善表达嵌合小鼠/人淀粉样蛋白前体蛋白和突变的人早老素1的双转基因小鼠肠道微生物群失调,调节碱性磷酸酶活性、脂多糖和丙二醛水平,从而改善AD小鼠的认知功能[17]。而本课题组早期也通过系统药理学研究和初步的细胞实验验证了佛手散治疗AD的分子机制与其抑制细胞外Ca2+内流、减少细胞内一氧化氮含量有关[18]。然而,佛手散治疗AD的抗氧化和抗细胞凋亡机制还需要进一步研究。因此,本研究探讨佛手散改善谷氨酸钠诱导PC12 细胞氧化应激损伤的作用机制,为佛手散的临床应用提供实验依据。
1 材料与方法
1.1 材料
1.1.1 实验仪器:SW-CJ-1FD超净工作台购自苏州净化设备公司,Multiskan MK3酶标仪购自Thermo Fisher Scientific公司, FACSCalibur型流式细胞仪购自Becton,Dickinson and company公司,INC108型细胞培养箱购自Memmert公司。
1.1.2 实验药品与试剂:当归饮片(批号:161101)和川芎饮片(批号:170401)均购自广州中医药大学第一附属医院,并经广州中医药大学喻良文副教授鉴定分别为伞形科植物当归的干燥根和伞形科植物川芎的干燥根茎。
谷氨酸钠购自上海麦克林生化科技股份有限公司(CAS编号:142-47-2;批号:L810495-100g);超氧化物歧化酶(superoxide dismutase,SOD)检测试剂盒、谷胱甘肽检测试剂盒、丙二醛检测试剂盒购自南京建成生物工程研究所(批号:A001-3-1、A003-1-1、A006-2-1);活性氧簇检测试剂盒购自上海碧云天生物技术有限公司(批号:S0033S);Caspase-3抗体和B细胞淋巴瘤2(B-cell lymphoma 2,Bcl-2)抗体购自武汉博士德生物工程有限公司(批号:BA2142、BA0412);Bcl-2相关X蛋白(Bcl-2 associated X protein,Bax)抗体购自Abcam公司(批号:Ab32503);胎牛血清购自Thermo Fisher Scientific公司(批号:10099-141);RPMI-1640培养基购自HyClone公司(批号:SH30809.01B);胰蛋白酶购自GENVIEW公司(批号:9002-07-7);RIPA裂解液、脱脂奶粉、TWEEN®20、GAPDH抗体、辣根过氧化物酶标记山羊抗兔抗体、辣根过氧化物酶标记山羊抗小鼠、ECL液购自武汉赛维尔生物科技有限公司(批号:G2002、G5002、WGT8220、GB12002、GB23303、GB23301、G2014);二喹啉甲酸(bicinchoninic acid,BCA)蛋白定量试剂盒购自Thermo Fisher Scientific公司(批号:23227);其他试剂均为分析纯,水为蒸馏水。
1.1.3 实验细胞:PC12细胞购自美国菌种保藏中心。
1.2 实验方法
1.2.1 佛手散的制备:按照2010年版《中华人民共和国药典》[19]配制佛手散。具体煎煮方法:当归、川芎按3 ∶2的质量比称重,加入8倍体积的水煎煮2 h,收集药液,再加入6倍体积的水继续煎煮2 h,收集药液,混合两次药液,60 ℃浓缩药液至最终浓度为0.64 g/mL,-20 ℃保存备用。基于已有的研究方法[16],使用前再将浓缩药液从冰箱中取出,解冻后用蒸馏水稀释至所需浓度。
1.2.2 分组及细胞造模:PC12细胞用含有10%胎牛血清和1%双抗(链霉素+青霉素)的RPMI-1640培养基,在37 ℃、5% CO2、相对饱和湿度的条件下培养,每2~3 d更换一次培养基。当细胞长至融合度为80%时,用胰酶消化后制备成单细胞悬液。将细胞接种到6孔板,密度调整为每孔2×105个,置于37 ℃、5% CO2、相对饱和湿度的细胞培养箱中培养,并将细胞随机分为对照组、模型组、佛手散低剂量组、佛手散中剂量组、佛手散高剂量组,每组设置3个复孔。根据本课题组的前期研究结果[20],以及有关细胞活性和凋亡的研究结果[18],将佛手散低、中、高剂量分别设定为32 mg/mL、64 mg/mL和128 mg/mL。
在佛手散低剂量、中剂量、高剂量组细胞中分别加入500 μL终浓度分别为32 mg/mL、64 mg/mL和128 mg/mL的佛手散预处理1 h,对照组和模型组用等体积PBS预处理1 h,除对照组外,其余各组均加入80 μL终浓度为 40 mmol/L谷氨酸钠孵育24 h,构建PC12细胞氧化应激损伤模型,用于后续实验。
1.2.3 SOD、丙二醛、谷胱甘肽和活性氧簇含量的检测:收集各组细胞,根据检测试剂盒说明书的操作方法测定各组PC12细胞SOD、丙二醛、谷胱甘肽和活性氧簇含量。
1.2.4 Caspase-3、Bax和Bcl-2蛋白表达水平的检测:收集各组细胞,用PBS反复冲洗,采用RIPA裂解液(含蛋白酶抑制剂)在冰上裂解细胞30 min,随后4 ℃下以12 000 r/min离心10 min,收集上清液,即为细胞的总蛋白。采用BCA蛋白定量试剂盒检测蛋白浓度。将蛋白质样品煮沸变性后采用SDS-PAGE分离蛋白,然后转至PVDF膜上。用5%脱脂奶粉- TWEEN®20在室温下封闭PVDF膜1 h。使用TBST洗膜3次,5 min/次。然后分别加入3 mL稀释后的Caspase-3抗体(1 ∶1 000)、Bax抗体(1 ∶1 000)、Bcl-2抗体(1 ∶1 000),4 ℃孵育过夜。使用TBST洗膜3次,5 min/次。然后加入3 mL稀释后的辣根过氧化物酶标记山羊抗兔抗体(1 ∶5 000),在室温下孵育40 min,使用TBST洗膜3次,7 min/次。以GAPDH(1 ∶5 000)为内参,用ECL显色,暗室曝光显影,采用凝胶图像分析系统测定目的蛋白条带的灰度值。
1.3 统计学分析 采用SPSS 25.0软件进行统计学分析。符合正态分布的计量资料以(x±s)表示,多组间比较采用单因素方差分析,方差齐时两两比较采用 LSD-t检验,方差不齐时两两比较采用Dunnett′s T3检验。以P<0.05为差异有统计学意义。
2 结 果
2.1 5组PC12细胞SOD、丙二醛、谷胱甘肽和活性氧簇含量的比较 模型组PC12细胞的SOD、谷胱甘肽含量低于对照组,丙二醛和活性氧簇含量高于对照组(均P<0.05)。佛手散低剂量组、佛手散中剂量组、佛手散高剂量组PC12细胞的SOD和谷胱甘肽含量高于模型组,丙二醛和活性氧簇含量低于模型组,且上述指标的变化呈剂量依赖性(均P<0.05)。见表1。
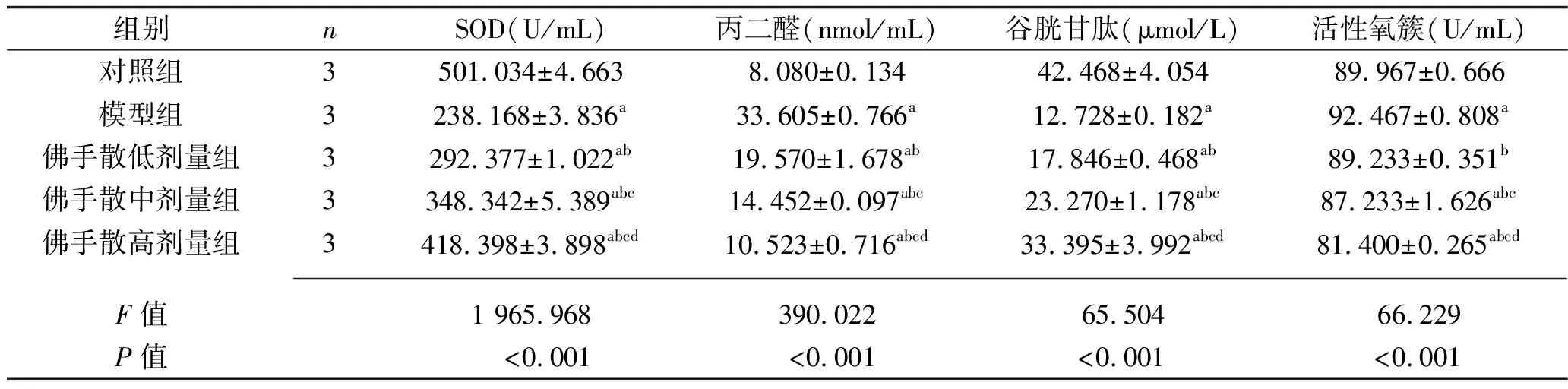
表1 5组PC12细胞SOD、丙二醛、谷胱甘肽和活性氧簇含量的比较(x±s)
2.2 5组PC12细胞凋亡蛋白表达水平的比较 模型组PC12细胞的Caspase-3和Bax表达水平高于对照组,Bcl-2蛋白表达水平低于对照组(均P<0.05)。佛手散低剂量组、佛手散中剂量组、佛手散高剂量组PC12细胞的Caspase-3和Bax表达水平低于模型组,Bcl-2蛋白表达水平高于模型组(均P<0.05),其中Caspase-3和Bax蛋白表达水平的变化呈剂量依赖性(均P<0.05),Bcl-2蛋白表达水平总体上随佛手散浓度的升高而升高。见表2和图1。
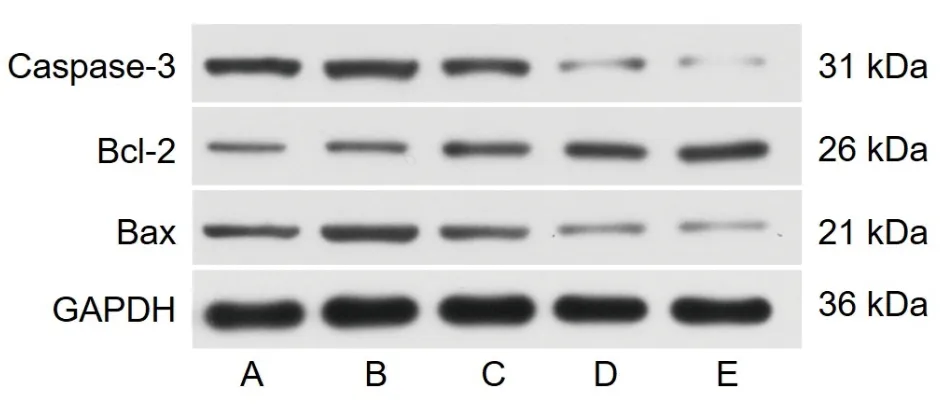
图1 5组PC12细胞中细胞凋亡相关蛋白表达的电泳图
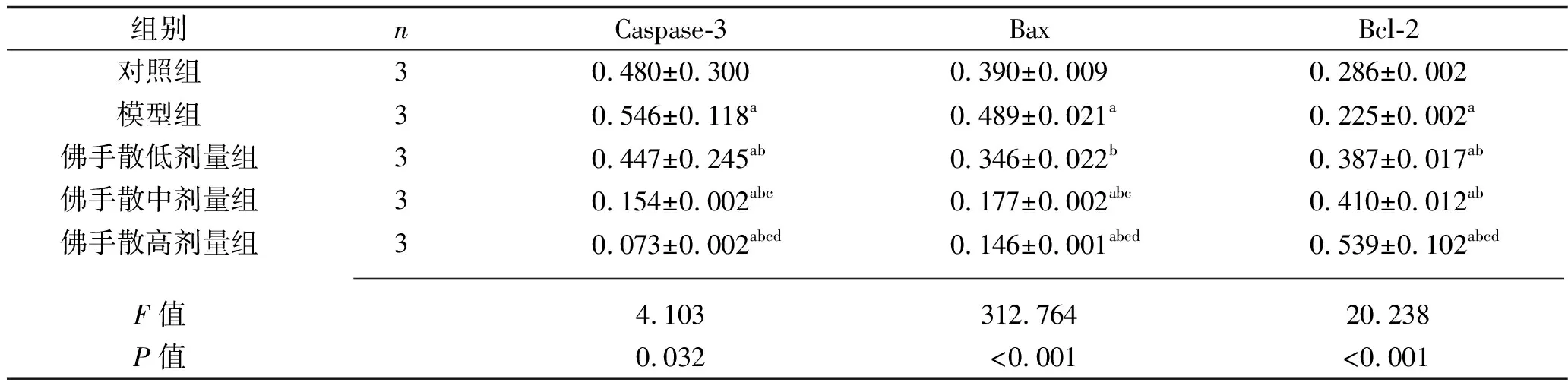
表2 5组PC12细胞凋亡蛋白相对表达水平的比较(x±s)
3 讨 论
研究显示,当归和川芎的主要成分对受损PC12细胞有保护作用[21-22],因此推测由当归和川芎组成的佛手散也可能对谷氨酸诱导的PC12细胞氧化应激损伤具有改善作用。本课题组前期的系统药理学研究结果显示,佛手散可通过抗氧化途径治疗AD[18],而且动物实验表明佛手散具有抗氧化作用[17],细胞实验也表明佛手散可通过抗氧化途径调节受损PC12细胞内的Ca2+和一氧化氮的浓度以起到细胞保护作用[18]。本研究进一步探讨佛手散改善谷氨酸钠诱导PC12 细胞氧化应激损伤的作用机制。
多项研究表明,AD患者大脑β淀粉样蛋白(amyloid beta-protein,Aβ)异常积累和神经纤维缠结沉积与氧化应激损伤有关[23-26]。Aβ的产生与氧化应激反应呈正相关,当大脑发生氧化应激损伤后,Aβ前体蛋白过表达,导致Aβ的形成和积累,同时伴随大量活性氧簇的产生,因此活性氧簇是AD发病的诱因之一[27],也是导致细胞结构受损、各种疾病状态和衰老的重要原因之一[28]。活性氧簇水平过高可能会导致自由基介导的连锁反应,如破坏细胞脂质、蛋白质、多糖甚至DNA[28]。活性氧簇诱发的氧化应激损伤使丙二醛水平升高,SOD和谷胱甘肽水平下降[29]。提高SOD和谷胱甘肽水平后,其可通过分解活性氧簇来降低细胞中活性氧簇的含量,抑制活性氧过量形成,减轻氧化应激损伤,同时抑制丙二醛的产生。因此,可通过检测这三种氧化应激副产物的水平来评价氧化应激损伤的程度[28,30]。本研究结果显示,模型组PC12细胞的SOD和谷胱甘肽含量低于对照组,丙二醛和活性氧簇含量高于对照组(均P<0.05)。这提示PC12细胞氧化应激损伤模型构建成功。高浓度的谷氨酸钠可能通过抑制细胞对胱氨酸的摄取,使细胞无法维持一定的谷胱甘肽水平以保护细胞免受氧化损伤,导致细胞产生过多的活性氧簇,从而促使细胞内丙二醛水平升高,SOD和谷胱甘肽水平下降[7]。而给予不同浓度佛手散预处理的PC12细胞丙二醛和活性氧簇含量低于模型组,SOD和谷胱甘肽含量高于模型组(均P<0.05),且指标变化呈剂量依赖性,表明佛手散可以减轻谷氨酸钠诱导的PC12细胞氧化应激损伤,且浓度越高,佛手散的抗氧化作用越显著,与既往研究[17,31]结论相似,提示佛手散或可对AD具有很好的抗氧化作用。
氧化应激可引起线粒体功能紊乱,促进细胞凋亡[32]。Bax和Bcl-2是两种与细胞凋亡相关的蛋白,二者均属于Bcl-2家族蛋白,Bcl-2家族蛋白与氧化应激密切相关,在调控线粒体凋亡通路中发挥重要作用。促凋亡蛋白Bax与Bak形成寡聚体后作用于线粒体外膜,导致线粒体外膜的通透性降低,使线粒体中的细胞色素C释放到细胞质中,激活凋亡因子Caspase-3,促进细胞凋亡;抗凋亡蛋白Bcl-2可与Bax竞争结合于线粒体细胞膜上,形成Bcl-2/Bax二聚体,使线粒体通透性降低,细胞色素C释放减少,从而抑制细胞凋亡[33-34]。因此,当Bax和Caspase-3表达增加,Bcl-2表达降低时,细胞凋亡增加[35]。研究显示,在哺乳动物和异体模型中Bax的表达升高可导致活性氧簇的产生和氧化损伤[36],而Bcl-2的表达升高可抑制由地塞米松和生长因子缺失引起的脂质过氧化[37]。当谷胱甘肽耗尽时,Bcl-2可通过增加或者改变细胞中谷胱甘肽的分布间接降低细胞中活性氧簇水平[36]。因此,Bcl-2家族蛋白可通过氧化应激通路调控细胞的氧化损伤。本研究结果显示,模型组PC12细胞的Caspase-3和Bax表达水平高于对照组,Bcl-2蛋白表达水平低于对照组(均P<0.05),说明谷氨酸钠可通过调控Bax、Caspase-3和Bcl-2的表达来调控细胞凋亡,这种凋亡可能与谷胱甘肽水平下降引起的氧化应激有关[7]。而给予不同浓度佛手散预处理的PC12细胞Caspase-3和Bax表达水平低于模型组PC12细胞,Bcl-2蛋白表达水平高于模型组PC12细胞(均P<0.05),其中Caspase-3和Bax表达水平的变化呈剂量依赖性,Bcl-2蛋白表达水平总体上随佛手散浓度的升高而升高,说明佛手散具有很好的抗凋亡作用,且浓度越高,其抗凋亡作用越显著。
综上所述,佛手散可以通过提高细胞的抗氧化能力和减少细胞凋亡,来抑制谷氨酸钠诱导的PC12细胞氧化应激损伤,从而发挥保护神经的作用。
