降香黄檀幼苗在重度Mn2+胁迫下不同时期的光响应
2023-08-12杨红兰黄秋婵李晓东梁海红甘宁冯守富
杨红兰,黄秋婵,李晓东,梁海红,甘宁,冯守富*
(1.广西民族师范学院 化学与生物工程学院,广西崇左 532200;2.贵港市大将国际学校,广西贵港 537100)
环境中重金属离子浓度超过一定的值,就会成为一个逆境因子胁迫植物各个生理过程,抑制植物的生长。大量研究表明,重金属污染会直接影响到植物的水分代谢、光合作用、呼吸作用、碳水化合物代谢以及氮素代谢等[1]。重金属一旦污染环境,其在土壤中会产生累积效应导致难以降解,而不同重金属元素之间具有协同、拮抗等相互效应,会使重金属污染在治理、修复等方面变得更为复杂。土壤中重金属含量过高,会影响植物的生长、发育及产量[2]。植物根系吸收重金属离子并运输到地上部分,会导致植物叶片受到重金属毒害,破坏叶绿体结构,最终造成植物叶片光合能力下降。
Mn2+在维持叶绿体的正常结构方面起着重要作用。植物受到Mn2+胁迫,内部叶绿体结构会发生改变,导致叶绿素含量降低,从而抑制光合作用,降低净光合速率[3]。汪结明[4]的研究表明,Mn2+胁迫会导致藤本植物薜荔、忍冬、五叶地锦和络石叶绿素总含量下降。土壤中Mn2+浓度过高还会对植物的生长发育及其产量造成不利影响。王洋[5]的研究显示Mn2+浓度过高会制约植物生长发育,引起植物自身结构及功能的改变。曾琦[6]研究发现,在植物的生长环境中随着Mn2+浓度的不断增大,地上部分中的生物量显著减少,说明过量的Mn2+会抑制植物生长。
降香黄檀(Dalbergia odoriferaT.Chen)又名花梨木,属豆科黄檀属植物,常绿半落叶乔木树种,在广东、广西及云南等地均有引种栽培[7]。降香黄檀具有优良的耐干旱耐贫瘠特性,是林地恢复和人工造林的理想树种。降香黄檀适应力强、萌芽率高,大树移载存活率高,树势恢复快,在园林环境绿化中是极具开发潜力的优良树种[8]。推进珍贵树种的地区引种,矿区环境修复研究,可深入挖掘其经济价值。
植物生长发育过程离不开光合作用,研究植物受重金属胁迫时的光响应,能够较为直接判断植物对重金属的适应性。目前关于降香黄檀引种栽培的研究很多,但关于降香黄檀对重金属胁迫光响应的研究较少。开展降香黄檀幼苗对重度Mn2+胁迫的光响应研究,对受Mn2+污染地区的适宜苗木的筛选具有重要意义,也对存在土壤Mn2+污染问题地区的人工育种、地区引种有着重要的实践意义。
1 材料与方法
1.1 材料与设备
实验所用降香黄檀种子采于广西民族师范学院校园,选择完好且饱满的种子,经泡种、发苗、培育得到胁迫实验所用幼苗。
YX1102 便携式光合测定仪,北京雅欣理仪科技有限公司。
1.2 幼苗培育方法
实验采用盆栽法,于口径15 cm 的种植盆各装取相同质量的混合壤(黑土∶红土∶营养土∶沙土=5 ∶2 ∶2 ∶1,质量比),每盆移栽5 株长势相似的幼苗(株高15 ~20 cm)。缓苗一周后,将MnSO4(结合前期实验0.4 mg/kg、2.0 mg/kg、10.0 mg/kg、50.0 mg/kg 浓度梯度结果,设定Mn2+浓度50 mg/kg 为重度胁迫)一次填埋施入(实验组),以不做填埋处理(Mn2+浓度为0 mg/kg)为对照组(CK),每个处理5 个重复。
1.3 光合参数的测定
分别于胁迫处理14 d(初期)、28 d(后期)测定各项光合指标,测定时间为8:00—15:00,从8:00 开始后每1 h 测一次。选取长势相同朝向一致的3 片成熟叶进行测量,每盆测量3 个重复,每次记录5 组数据。测定的光合指标包括幼苗的净光合速率(net photosynthetic rate,Pn)、蒸腾速率(Transpiration rate,Tr)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)、光合有效辐射(Photosynthetically Active Radiation,PAR)、大气相对湿度(Relative air Humidity,RH)、大气二氧化碳浓度(air CO2concentration,Ca)。
1.4 数据分析
所有数据采用Excel 2010 进行处理和作图分析。将5 组重复处理的数据去除波动较大的数值,最后取均值进行数据分析并作图。
2 结果与分析
各检测指标的日均值统计表见表1。
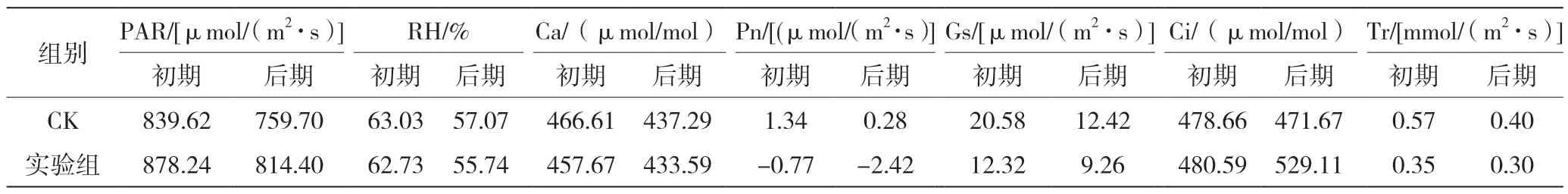
表1 各项指标日均值统计表
2.1 环境因子日变化
植物的光合作用,不单由内在生理因子制约,同时还会受外界环境因子的影响。由图1 可知,胁迫初期PAR 日变化和CK 的PAR 日变化均呈“双峰型”,实验组PAR 在9:00 出现第一个峰值,在12:00 出现第二个峰值,PAR 最大值为1 617.50 μmol/(m2·s),胁迫初期PAR 的日平均值为878.24 μmol/(m2·s)。胁迫后期,PAR 日变化曲线规律性不明显,实验组的PAR 最大值为1 376.00 μmol/(m2·s),峰值出现在15:00,日平均值为814.40 μmol/(m2·s)。胁迫初期、后期实验组的平均PAR 分别比CK 组高4.6%和7.2%。
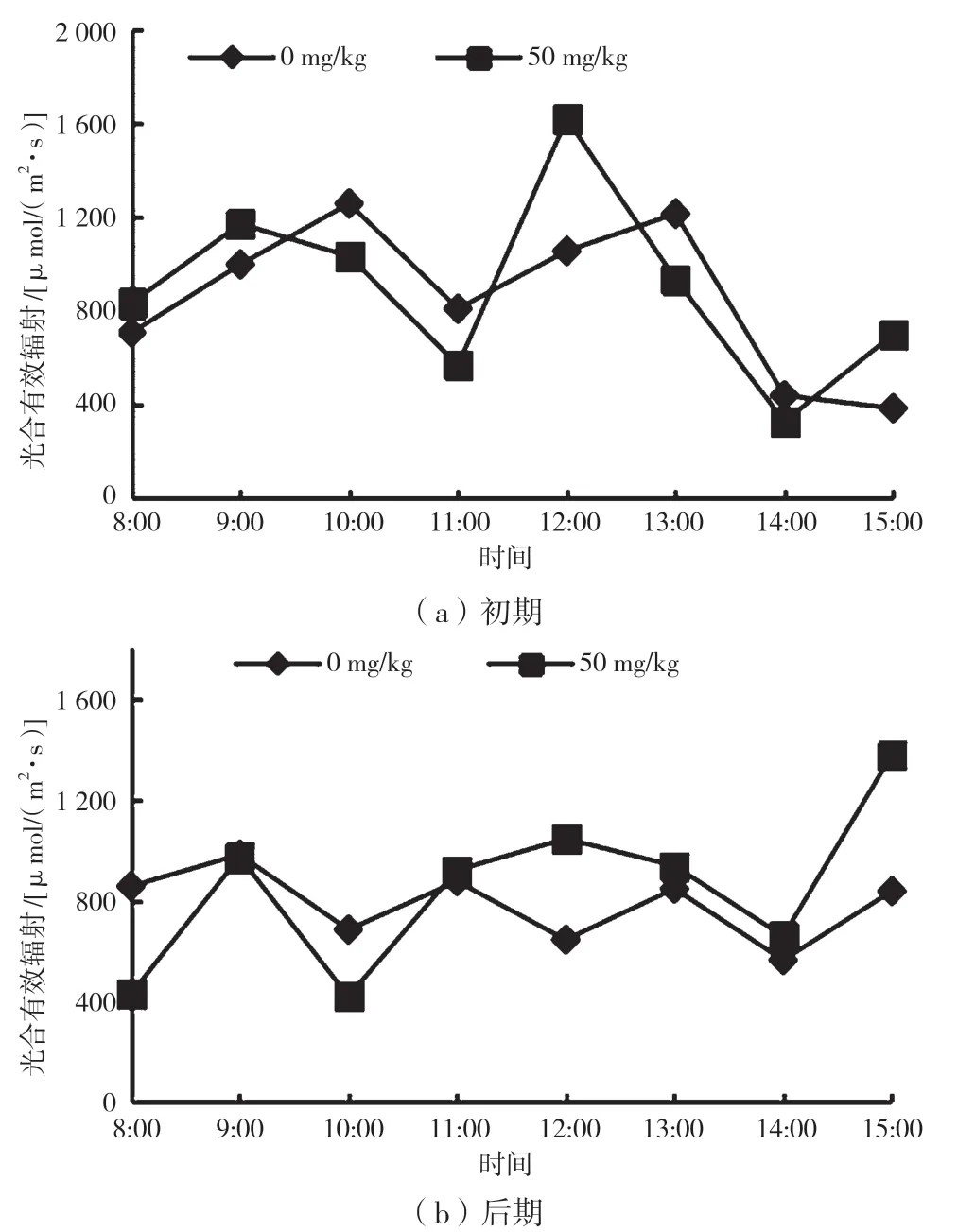
图1 不同时期光合有效辐射日变化曲线
由图2 可知,胁迫初期大气相对湿度的日变化呈不明显的“W”型。两组试验的RH 均在10:00 出现最低值;胁迫后期RH 总体呈现下降的趋势。由图3 可知,胁迫初期Ca 呈“单峰型”曲线,最大值出现在9:00,之后持续下降,胁迫初期Ca 日均值为457.67 μmol/mol;胁迫后期Ca 没有出现明显的峰值,总体呈下降趋势,日均值为433.59 μmol/mol。
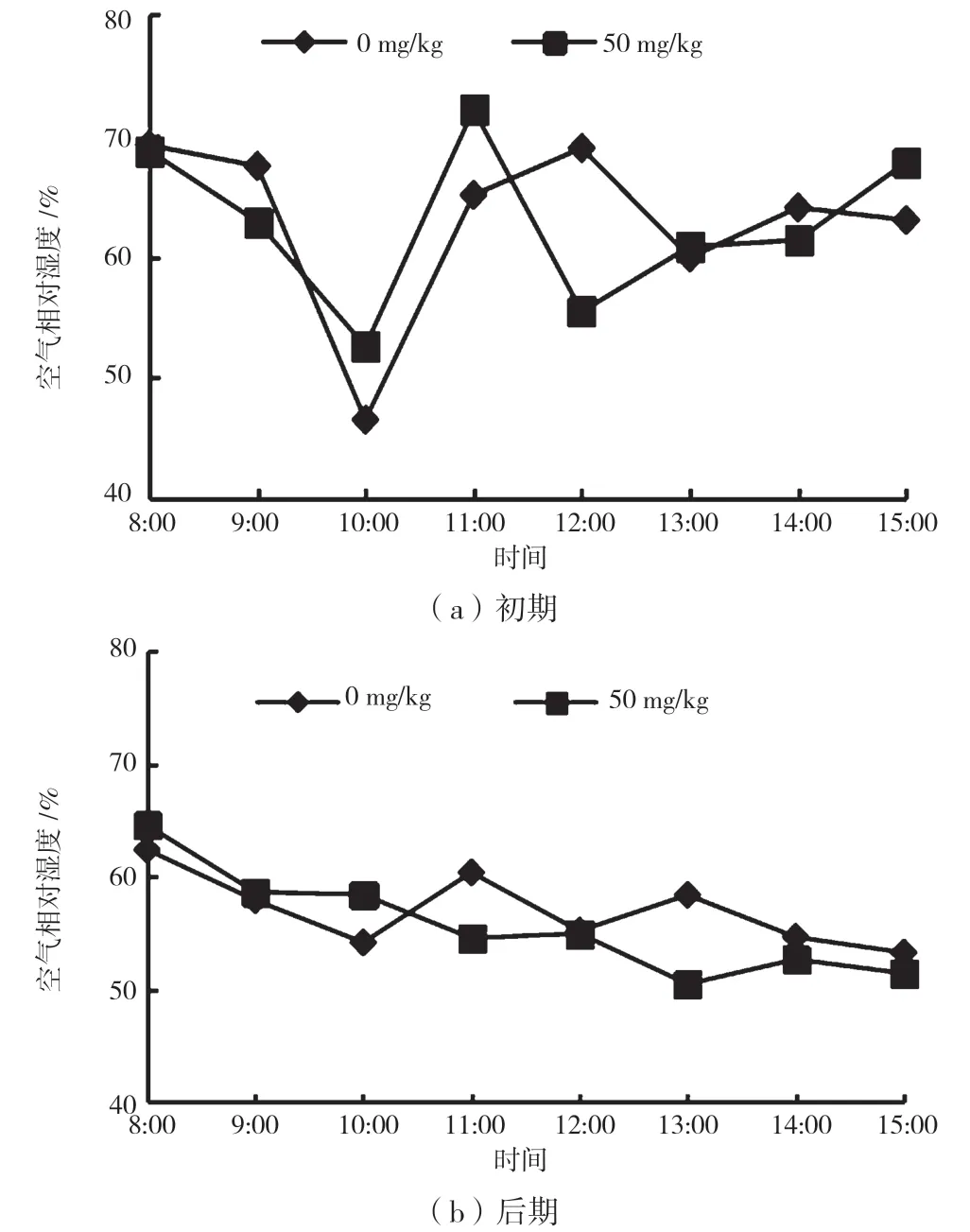
图2 不同时期空气相对湿度日变化曲线

图3 不同胁迫时期大气CO2 浓度日变化曲线
2.2 生理因子日变化
2.2.1 净光合速率(Pn)日变化
光合速率日变化的午休现象在自然界中普遍存在,是植物对环境的一种适应机制,植物光合作用的日变化曲线大体有单峰型和双峰型两种。从图4 可以看出,降香黄檀幼苗在胁迫初期、后期两组的净光合速率(Pn)日变化模式相似。
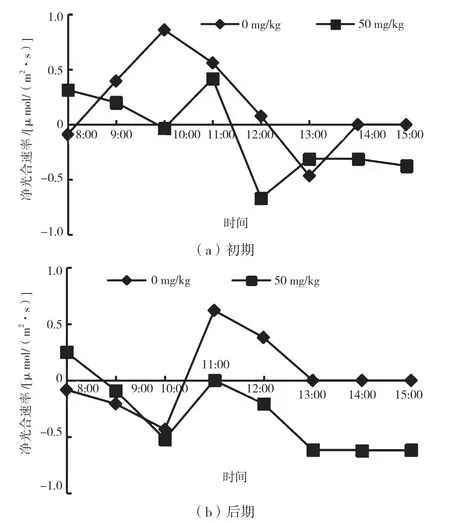
图4 不同胁迫时期净光合速率日变化曲线
光是植物进行光合作用的重要能量型原料,但不同植物对光的响应不同[9]。在重度Mn2+胁迫初期,两组的Pn 值呈不明显的“双峰型”曲线。实验组从8:00到10:00,PAR 先增大后有所降低,而Pn 逐渐减小;在10:00—12:00,PAR 先减小后增大(见图1),而Pn 先上升后下降,且在12:00 出现最低值;在胁迫初期Pn 日变化曲线中,Pn 最大值为0.42 μmol/(m2·s),CK 和实验组Pn 峰值出现的时间有所不同。在胁迫初期,与CK 组相比,受重度Mn2+胁迫的降香黄檀幼苗对PAR 变化的影响更为敏感,且光合能力有所下降。
在胁迫后期CK 组和实验组Pn 日变化趋势基本一致,表明受一段时间的重度Mn2+胁迫之后,实验组植株对重度Mn2+的适应性增强。胁迫后期CK 组的Pn 峰值为0.62 μmol/(m2·s),出现在11:00;实验组Pn 最大值为0.25 μmol/(m2·s),出现在8:00。在10:00 之后,随着PAR 的变化,实验组Pn 始终明显低于CK 组,表明受重度Mn2+胁迫的降香黄檀幼苗,光合作用能力明显下降,这可能是由于植物光合作用被抑制[10]。
2.2.2 气孔导度(Gs)日变化
由图5 可知,在重度Mn2+胁迫初期,CK 组和实验组最大值均出现在9:00,CK 组的Gs 最大值为50.44 μmol/(m2·s),实验组Gs最大值为22.83 μmol/(m2·s)。实验组气孔导度日变化曲线在11:00—15:00 与CK组气孔导度日变化曲线变化呈现相反趋势。CK 组平均气孔导度为20.58 μmol/(m2·s),实验组平均气孔导度为12.32 μmol/(m2·s),比CK 组下降了40.1%。
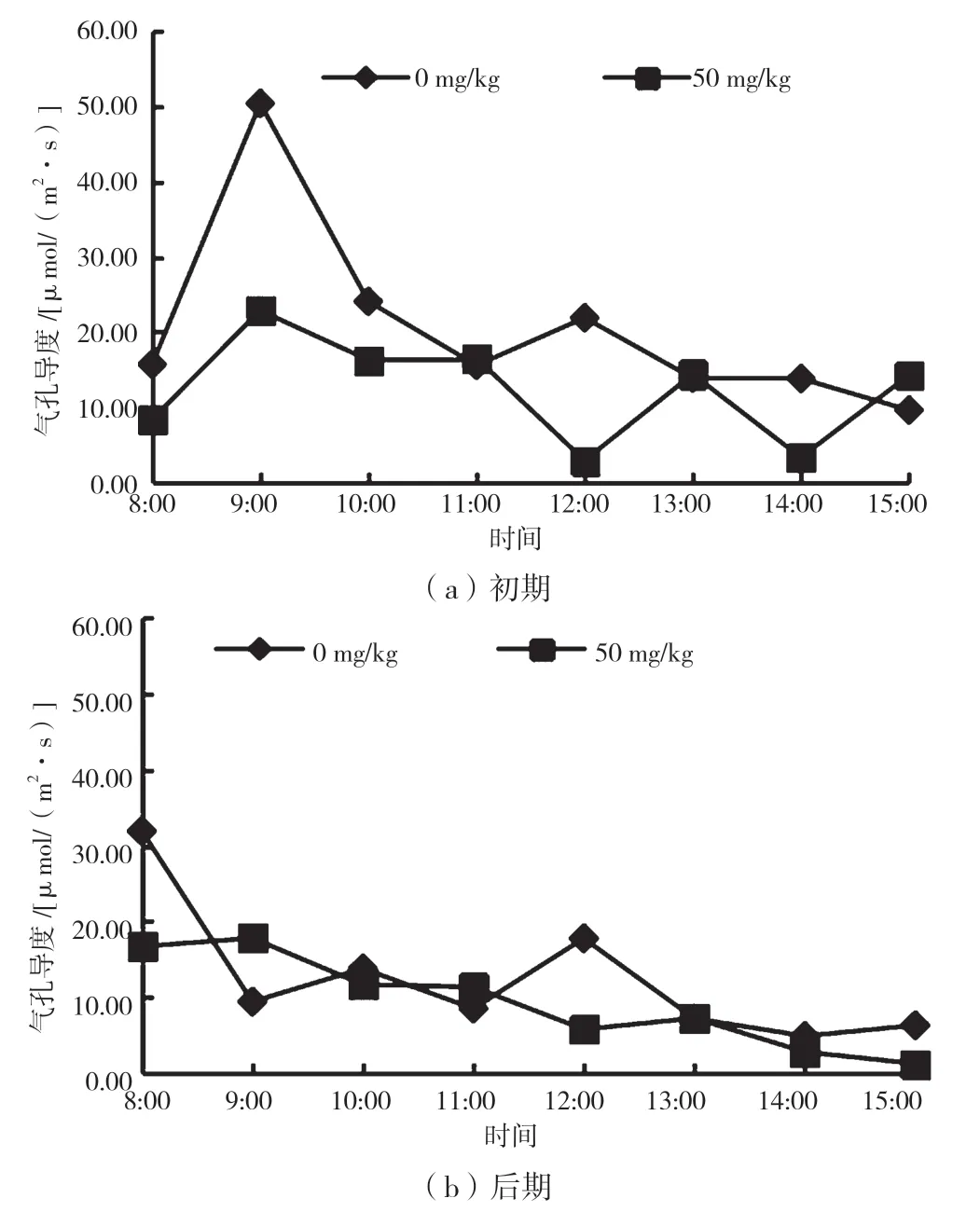
图5 不同胁迫时期气孔导度日变化曲线
在重度Mn2+胁迫后期,CK 组和实验组的Gs 日变化曲线趋势相似,最大值均在8:00 出现,CK 组平均气孔导度为12.42 μmol/(m2·s),实验组平均气孔导度为9.26 μmol/(m2·s),与CK 组相比下降了25.4%。整体看来,胁迫后期实验组气孔导度日变化曲线波动更为平缓。
2.2.3 胞间CO2 浓度(Ci)日变化
光是主要的能量原料也是主要的环境因子,光的变化会影响内部因子变化,从而影响光合速率[9-10]。结合图1、图4 和图6 可知,CK 组在PAR 升高的初始阶段,叶片Pn 增加,消耗CO2量增大,导致Ci 下降。胁迫初期和后期的8:00—9:00,CK 组的Ci 值均高于实验组,10:00 之后随着PAR 的变化,实验组Ci 浓度开始高于CK 组。胁迫初期,CK 组的Ci 日均值为478.66 μmol/mol,实验组Ci 日均值为480.59 μmol/mol,较CK 组高0.4%;胁迫后期,Ci 日变化曲线呈“单峰型”,CK 组的Ci 日均值为471.67μmol/mol,实验组Ci日均值为529.11 μmol/mol,较CK组高10.8%。可见,随着胁迫时间的增加,降香黄檀幼苗通过增大胞间二氧化碳浓度适应高锰环境。
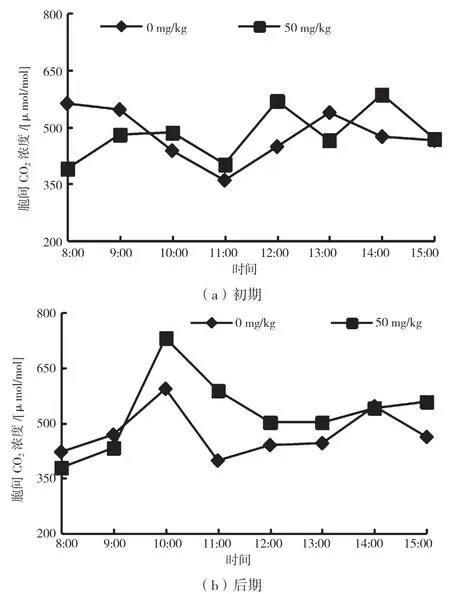
图6 不同胁迫时期胞间CO2 浓度日变化曲线
2.2.4 蒸腾速率(Tr)日变化
如图7 所示,Tr 的最大值出现的时间与Gs 最大值出现的时间相似,胁迫初期蒸腾速率峰值出现在9:00,且实验组峰值低于CK 组。胁迫初期CK 组的Tr 日均值为0.57 mmol/(m2·s),实验组Tr 日均值为0.35 mmol/(m2·s),较CK 组下降了38.6%。胁迫后期CK 组Tr 日均值为0.4 mmol/(m2·s),实验组Tr 日均值为0.3 mmol/(m2·s),较CK 组下降了25.0%。在整个胁迫周期中,实验组降香黄檀幼苗Tr值基本上均小于CK 组,且后期下降较前期少。
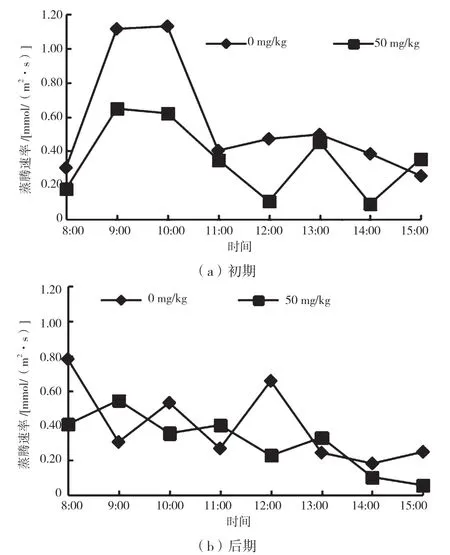
图7 不同胁迫时期蒸腾速率日变化曲线
3 讨论与结论
Mn 是植物生长必需的微量元素之一,缺少或者过量都会影响植物的正常生长。本实验对降香黄檀幼苗在重度Mn2+胁迫下的光合生理日变化进行分析,结果发现受重度Mn2+胁迫的降香黄檀幼苗的日蒸腾速率(Tr)、气孔导度(Gs)、净光合速率(Pn)较CK均有所下降,胞间CO2浓度(Ci)相反,较CK 有所上升。
Mn2+与植物的光合作用关系密切,受重度Mn2+胁迫的降香黄檀幼苗Pn 较CK 明显下降,这可能与降香黄檀幼苗叶片中叶绿素含量下降有关[11]。降香黄檀幼苗光合过程同时受到诸多生理因子和环境因子的调控,PAR 是环境因子中主要的影响因素,随着PAR 的变化,RH 和Ca 受到影响。在外界环境因子的影响下,Ci、Gs、Tr 等光合生理指标发生变化,最终引起Pn 的变化。
Gs、Tr 在胁迫初期,较CK 分别下降了40.1%、38.6%,随着胁迫时间的增加,幼苗表现出了对重度Mn2+的适应性;在胁迫后期,Gs、Tr 较CK 分别下降了25.4%和25.0%。Ci 的变化趋势则相反,Ci 较CK数值有所增加,由胁迫初期上升0.4%到胁迫后期上升10.8%,表明重度Mn2+胁迫对降香黄檀幼苗的光合作用造成一定抑制。受重度Mn2+胁迫的降香黄檀幼苗Pn 日峰值同样低于CK,在胁迫初期,CK 的Pn 日峰值为0.86 μmol/(m2·s),到12:00 时接近于零,实验组的Pn 日峰值为0.42 μmol/(m2·s);胁迫后期,CK 的Pn 日峰值为0.62 μmol/(m2·s),实验组的为0.25 μmol/(m2·s),两个时期的实验组Pn 值基本都小于CK 组,说明重度Mn2+胁迫对降香黄檀幼苗的Pn 有明显抑制作用。
综上,重度锰胁迫下降香黄檀幼苗在两个不同的时期光响应表现出明显差异。环境因子PAR 和生理因子Ci日均值在两个时期均表现为实验组高于CK组。而Gs 和Tr 日均值则体现出两个时期均是实验组低于CK 组。Pn 对重度Mn2+胁迫则较敏感。以上结果说明,重度锰胁迫对降香黄檀幼苗的光响应具有重要影响。
