海南常见红树植物叶片脂肪酸组成
2023-08-09杨睿尧谢丽梅安文硕陈顺洋陈光程
杨睿尧,谢丽梅,安文硕,陈顺洋,陈 彬,4,陈光程,4*
(1.上海海洋大学海洋科学学院,上海 201306;2.自然资源部第三海洋研究所,福建 厦门 361005;3.厦门大学环境与生态学院,福建 厦门 361005;4.自然资源部北部湾滨海湿地生态系统野外科学观测研究站,广西 北海 536015)
分布在热带、亚热带地区潮间带的红树林通常会促进潮间带生境的复杂性和多样性,为诸多生物提供栖息和觅食的环境,为各种生物的生存繁衍提供了物质基础,维持着与其密切相关的底栖动物群落和生物多样性[1]。脂肪酸是植物细胞重要的组成部分,其组成与植物自身的生理功能密切相关,在不同物种和地理区域表现出明显的差异[2-3]。脂肪酸在动物摄食过程中能够相对保守地同化至消费者体内,随着食物链传递至下一营养级,通过比较脂肪酸成分,能够反映一段时间内动物的摄食情况[4]。一些学者也指出,红树植物体内多不饱和脂肪酸种类多,含有较多人体和动物必需的脂肪酸,是潜在的脂肪酸资源[5-6]。因此脂肪酸的组成分析对认识红树植物脂肪酸资源、红树林生态系统内部的营养关系、食物网的结构,以及红树植物对生境的适应等具有重要参考价值[7]。
目前我国针对秋茄(Kandeliaobovata)、白骨壤(Avicenniamarina)、桐花树(Aegicerascorniculatum)和无瓣海桑(Sonneratiaapetala)等广布种开展了植物体脂肪酸的研究[5,8-10],但总体上研究仍有限,对于热带地区红树物种的脂肪酸组成及差异的报道较少。本研究比较了我国海南三亚铁炉港和文昌清澜港常见红树植物叶片脂肪酸的组成,分析植物叶片脂肪酸在物种和区域之间的差异,以期丰富红树植物脂肪酸组成、资源利用潜力和生态学作用的信息。
1 材料与方法
1.1 研究区域
本研究的植物样品采集自海南岛南部的三亚市铁炉港(18°15′N,109°42′E)和东部的文昌市清澜港(19° 54′N,110° 84′E)。这两个港湾属于热带季风气候,三亚市的年均气温为 25.5 ℃,年均降水量为1 738 mm;文昌市的年均气温为24.1 ℃,年均降水量为2 106 mm[11-12]。铁炉港地貌上属沙坝-潟湖型港湾,潮汐为不规则日潮型;土壤以砂土、砂质粘土为主,多砂石及贝壳珊瑚碎屑[13]。清澜港属潟湖潮汐通道港湾,潮汐为不规则日潮型,土壤以细砂、砂-粉砂-粘土为主[14]。
1.2 植物样品采集与脂肪酸分析
本研究在铁炉港和清澜港分别采集红海榄(Rhizophorastylosa)、杯萼海桑(Sonneratiaalba)、木果楝(Xylocarpusgranatum)、角果木(Ceriopstagal)、榄李(Lumnitzeraracemosa)、木榄(Bruguieragymnorrhiza)和正红树(Rhizophoraapiculata)等7种植物的叶片。随机采集红树植株上的成熟叶片,在每株植物上采集3~5片成熟叶片合并为1份样品,每个物种采集3份样品。样品运回实验室后用蒸馏水洗净,于-40 ℃冷冻干燥后研磨待测。
本研究中脂肪酸的表达式写为CA:BnXt或CA:BnXc,A表示脂肪酸碳链的碳数,B表示双键的数量,X表示第一个双键离甲基端的碳数,t表示反式脂肪酸,c表示顺式脂肪酸。碳链中含一个双键的脂肪酸为单不饱和脂肪酸,含两个及以上双键的脂肪酸为多不饱和脂肪酸。脂肪酸的提取和测定参考崔莹[15]和刘梦坛[16]的方法进行改进。取50 mg样品,加入C19:0作为内标,用体积比为2∶1的二氯甲烷与0.01%BHT-甲醇混合液在80 ℃水浴下加热2 h提取脂质,60 ℃水浴蒸干后加入3 mL甲醇和1 mL盐酸,50 ℃条件下水浴4 h进行甲酯化。结束后迅速冷却至室温,用正己烷萃取,样品定容至1 mL。脂肪酸组成和含量用气相色谱仪测定,使用HP88色谱柱(60 m×0.25 mm×0.2 μm),柱箱初始温度为 100 ℃,以 10 ℃/min 程序升温至210 ℃,保持2 min,再以 1.5 ℃/min 程序升温至240 ℃,保 持 3 min。采用峰面积归一化法得到各种脂肪酸成分在样品中的含量占比情况。样品加标回收率为70%~90%;脂肪酸含量检测限为1.01 μg/mL。
1.3 数据处理
运用SPSS 23.0软件,对铁炉港和清澜港红树植物样品中脂肪酸的含量在不同物种之间的差异进行单因素方差分析比较。基于样品的脂肪酸含量组成,在Primer软件中采用聚类分析对样品进行分组,再用相似性分析(One-way ANOSIM)和相似性百分比分析(Simper)检验植物样品处理组之间的相似性。不同植物样品中脂肪酸组成的差异用主成分分析(Principal Component Analysis,PCA)的多维度分析(OriginPro 2019b)。
2 结果与讨论
2.1 红树植物叶片脂肪酸种类组成
在铁炉港和清澜港采集的红树叶片中共检测出25种脂肪酸(表1、2),铁炉港的植物样品中检测到22种,十五烷酸(C15:0)、亚麻酸(C18:3n3)和芥酸(C22:1)未检出;清澜港的样品中检测到23种,葵酸(C8:0)和二十碳五烯酸(C20:5)未检出。月桂酸(C12:0)、肉豆蔻酸(C14:0)、棕榈酸(C16:0)、硬脂酯酸(C18:0)、油酸(C18:ln9c)、亚油酸(C18:2n6c)、二十碳烯酸(C20:ln9)和二十四烷酸(C24:0)等8种脂肪酸在所有样品中均有检测出。样品中共检测到16种饱和脂肪酸(saturated fatty acid,SFA),不饱和脂肪酸(unsaturated fatty acid,UFA)包括5种单不饱和脂肪酸(mono unsaturated fat acid,MUFA)和4种多不饱和脂肪酸(polyunsaturated fatty acid,PUFA)。
所有脂肪酸中含量最高的是SFA中的C16:0,在所有样品中占比均超过30.00%(30.06%~60.68%),除铁炉港的红海榄外,样品中C18:0(4.38%~12.05%)占比也高于其他饱和脂肪酸。含量较高的UFA有C18:ln9c、C18:2n6c和C20:ln9。其中C20:ln9在木榄和木果楝中的占比均超过20.00%。木果楝和清澜港的木榄样品中C18:2n6c的占比也超过20.00%。脂肪酸不饱和指数(以UFA/SFA总含量比值表示)在0.19~1.13之间,木果楝和清澜港的木榄样品中脂肪酸不饱和指数大于1。碳原子数小于12的脂肪酸只在木果楝和木榄中检测到,最短链脂肪酸为己酸(C6:0)。最长链脂肪酸为三十烷酸(C30:0),占比介于0.46%~5.22%,铁炉港的正红树中未检测到,清澜港的红海榄和杯萼海桑中也未检出。
2.2 红树植物叶片脂肪酸含量比较
在两个港湾,相同的红树物种叶片所含脂肪酸数量和组成有所不同,其中C16:0、C18:2n6c和C20:ln9的变化较明显(表1、2)。在脂肪酸的组成上,这3种脂肪酸为所有红树植物脂肪酸的主要组成。正红树、角果木和木果楝的C16:0含量在两个港湾之间接近,红海榄和杯萼海桑的C16:0含量则表现为铁炉港低于清澜港。两个港湾木果楝的C18:2n6c和C20:ln9含量也相似,木榄的C20:ln9含量在两地之间也接近,而C18:2n6c含量在两地之间差异较大。除正红树、木果楝和木榄外,其他物种的饱和脂肪酸占比表现为清澜港显著高于铁炉港(P<0.05,图1)。清澜港和铁炉港红树植物脂肪酸含量也表现出不一样的种间差异。样品中C16:0含量在不同物种中的差异较大,两个港湾红海榄C16:0含量显著高于其他物种,木果楝的含量则最低。相反,木果楝中含有较高含量的C18:2n6c和C20:ln9(占比>20.00%),并且脂肪酸不饱和指数大于1,表现出与其他物种不同的脂肪酸组成特征。木榄中的C20:ln9占比在两港湾均超过20.00%,但C18:2n6c的含量在清澜港和铁炉港之间存在差异。
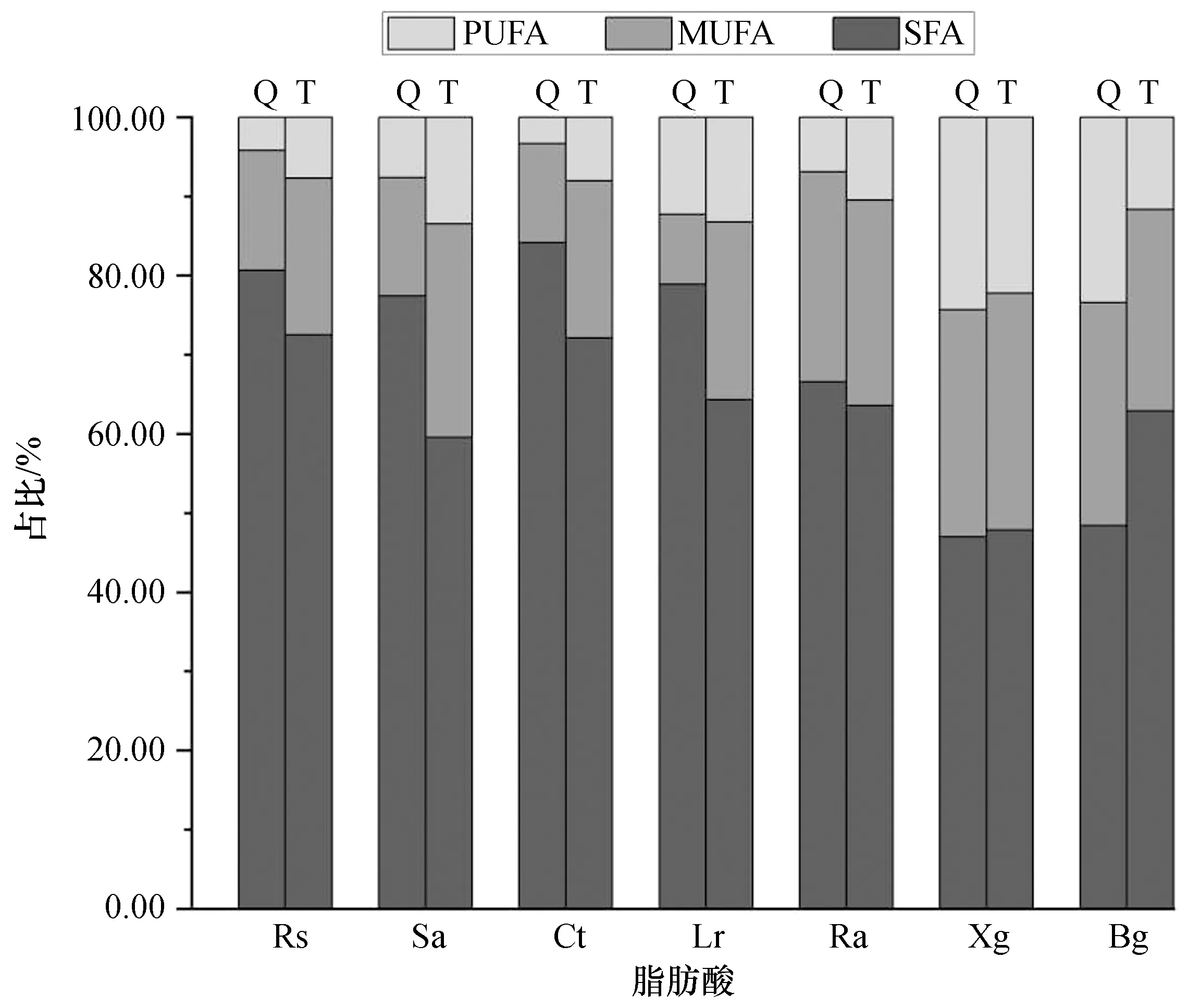
图1 海南铁炉港和清澜港红树植物叶片脂肪酸中各类脂肪酸的占比Fig.1 Proportion of various fatty acids in the leaves of mangrove plants in Tielugang and Qinglangang,HainanPUFA:多不饱和脂肪酸,MUFA:单不饱和脂肪酸,SFA:饱和脂肪酸;Rs:红海榄,Sa:杯萼海桑,Ct:角果木,Lr:榄李,Ra:正红树,Xg:木果楝,Bg:木榄;Q:清澜港,T:铁炉港,下同。
2.3 基于脂肪酸组成的树种差异和空间差异分析
红树叶片脂肪酸组成的聚类分析结果显示,红树植物脂肪酸组成因物种和采样地点而异(图2),在7种红树植物中,两个港湾样品中脂肪酸组成相似水平达90%以上的只有木果楝和正红树两个物种,其他物种在两个采样区域表现出明显的差异。基于聚类分析的结果,在相似性71%水平下区分为3组进行One-way ANOSIM分析。结果表明,组I与组II(R=0.434,P=0.012)、组I与组III(R=0.935,P=0.022)之间存在显著差异。对I、II组之间差异性贡献最大的脂肪酸是C16:0,其次是C20:ln9和C18:2n6c;对II、III组之间差异性贡献最大的同样为C16:0,其次是C18:0、C18:ln9c和18:2n6c;对I、III组之间差异性贡献最大的则是C20:ln9。

图2 海南铁炉港和清澜港红树植物叶片脂肪酸组成的聚类分析结果Fig.2 Cluster analysis results of fatty acid composition of mangrove leaves collected from Tielugang and Qinglangang,Hainan
PCA分析将红树植物的脂肪酸组成分为4组(图3),两个港湾相同物种的脂肪酸组成上只有木果楝、正红树、红海榄和木榄处于相同分组,在相似性上两个港湾红海榄和木榄的相似程度低于木果楝和正红树,结果与聚类分析结果相似。对两个港湾其余物种Simper分析表明,造成各组内部物种之间差异贡献最大的脂肪酸是C16:0(贡献率35.47%~56.67%);C16:0是造成组4与其他3组之间差异性贡献最大的脂肪酸(贡献率12.43%~25.51%);C20:ln9是造成1、2组与2、3组之间差异性贡献最大的脂肪酸(贡献率21.94%、19.23%);1、3组之间的差异性则由18:2n6c 引起(贡献率25.94%)。
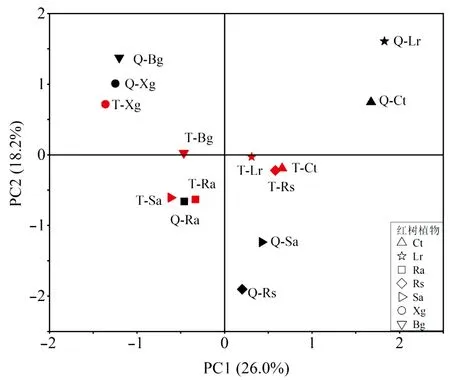
图3 海南铁炉港和清澜港红树植物叶片的脂肪酸组成PCA分析结果Fig.3 PCA analysis results of fatty acid composition of the mangrove leaves collected from Tielugang and Qinglangang,Hainan红色图形为铁炉港,黑色图形为清澜港。
2.4 讨论
本研究调查了海南三亚铁炉港和文昌清澜港7种红树植物叶片的脂肪酸组成及其差异。在样品中检测到25种脂肪酸,脂肪酸碳链长度集中在C12~C20之间,与前人对秋茄、桐花树、木榄、无瓣海桑等红树叶片脂肪酸的研究相一致[8-10]。本次研究中红树植物脂肪酸以C20以下脂肪酸为主,其中C20脂肪酸含量低于C16和C18脂肪酸含量。植物脂肪酸的合成是乙酰辅酶A在乙酰辅酶A羧化酶和脂肪酸合成酶的共同作用下发生连续的聚合反应,最终合成C16至C18的饱和脂肪酸,大多数高等植物脂肪酸碳链长度在C18以下[2],由于普通高等植物缺少脂肪酸碳链延长的相关酶(如Δ6脂肪酸去饱和酶),所以高等植物C18以上的多不饱和脂肪酸含量较少[17]。脂肪酸组成上,C16:0、C18:0、C18:ln9c、C18:2n6c和C20:1在各样品中均有较高的含量,在所有样品中含量最高的C16:0已被证实为红树植物叶片脂肪酸组成中的主要成分。一些研究指出红树植物叶片中不饱和脂肪酸含量通常高于饱和脂肪酸含量[5,10]。本研究则发现,除木果楝外的其他红树植物叶片脂肪酸的组成以饱和脂肪酸为主,木榄的不饱和脂肪酸含量在铁炉港和清澜港之间也存在明显的差异,说明红树植物脂肪酸的组成上因地区和物种等因素而异。这种差异在以往的研究中也被报道,例如,珠海淇澳岛的木榄样品中饱和脂肪酸含量低于不饱和脂肪酸,但湛江特呈岛的饱和脂肪酸含量可达到不饱和脂肪酸的两倍[10]。除物种和地理因素外,谢丽梅等发现,红树叶片衰老和腐烂后脂肪酸不饱和指数出现明显的变化[18]。
由于生物体自身代谢水平的差异,一些生物类群会合成特异性的脂肪酸种类或是趋向于较多地合成某几种脂肪酸,这些脂肪酸就是该生物类群的特征脂肪酸[2]。在滨海湿地植物中,碳链长度大于24的脂肪酸被认定为红树植物的特征脂肪酸而被广泛用于指示红树植物对海洋动物的食物供给[7]。红树植物的特征脂肪酸组成存在明显的种间差异和区域差异,其长链脂肪酸的组成和含量存在很大的不确定性,在一些物种中存在两种及以上的长链脂肪酸,但一些物种中甚至未能检测到长链脂肪酸[8-10]。本研究中红树植物的特征脂肪酸组成也表现出种间和区域间的差异;清澜港的红海榄和杯萼海桑,以及铁炉港的正红树中未检测出特征脂肪酸。因此,采用脂肪酸示踪研究红树林动物食源时,需根据植物脂肪酸的组成选择特征脂肪酸,并结合胃含物分析和稳定同位素示踪等方法提高结果的准确性[7]。
除自身的遗传特性决定外,植物体内脂肪酸组成也受多种环境因素的影响[19]。低温、光照、病害和盐度等环境因素对植物不饱和脂肪酸的合成有一定的调控作用[20]。由于植物脂肪酸组成与植物细胞膜的结构和功能密切相关,不饱和脂肪酸含量增加则会提高膜的流动性,提高植物对环境的抗逆性和适应性[21]。本研究中C18:ln9c、C18:2n6c和C20:1是造成不同红树物种之间差异性的主要脂肪酸,铁炉港和清澜港红树叶片中不饱和脂肪酸的含量也存在差异,除木果楝和木榄外其他物种总体上表现出清澜港低于铁炉港。光照时长和气温可能是造成两个研究区域之间红树叶片脂肪酸组成差异的因素。三亚年日照时长(2 500 h)高于文昌(1 800 h)[14]。光照时长已被证实会使植物脂肪酸的含量和组成产生明显差异,随着光照时间的延长,不饱和脂肪酸如C18:ln9c的占比增加[22-23]。植物的脂肪酸不饱和指数与植物对温度的适应性密切相关[24],低温能够诱导植物中脂肪酸的去饱和相关酶表达,使脂肪酸不饱和指数升高,提高植物的耐寒性[25-26]。本研究样品采集于低纬度的热带地区,这些植物叶片的脂肪酸不饱和指数总体上低于其他地区报道的秋茄、桐花树、老鼠簕(Acanthusilicifolius)等物种的结果[5,10],说明这些红树林物种的地理分布可能与它们体内脂肪酸不饱和指数关系密切。以红海榄为例,其脂肪酸不饱和指数在福建九龙江口和广东湛江(特呈岛)分别为1.12和0.76[5,27],在清澜港和铁炉港则分别为0.38和0.24,总体上随纬度降低而降低。在环境温度的变化中能否调节体内脂肪酸组成的能力也关系着植物对低温条件的适应性[28],因此,我们推测一些红树林广布种可能具有较强的脂肪酸调控能力以适应不同的气温条件,但还需要进一步的验证。
3 结论
本研究结果表明,海南三亚铁炉港和文昌清澜港7种红树植物的脂肪酸以偶数碳原子脂肪酸为主,其中C16:0在所有植物中含量最高,红树植物中存在C26:0和C30:0等特征脂肪酸。不同红树物种之间脂肪酸的组成不同,除木榄和正红树外的其他红树植物的脂肪酸在两个区域之间也存在差异,这些差异主要归因于植物叶片中C16:0、C18:ln9c和C18:2n6c含量的变化。总体上,铁炉港红树植物中不饱和脂肪酸占比高于清澜港,但两地的红树植物中脂肪酸不饱和指数较低,可能与温度和光照时长等环境因素有关。因此,脂肪酸的组成可应用于红树植物对环境因素生理响应的研究,而不同物种在其适宜分布的地理区域内对环境适应性的差异、脂肪酸组成是否反映热带地区的植物种源北移的抗寒性等问题还需要进一步探索。
