不同光质对坛紫菜果孢子和壳孢子萌发的影响
2023-08-09刘嘉嘉梁小君周琳琳钟晨辉郭辰涛唐隆晨
刘嘉嘉,梁小君,周琳琳,钟晨辉,郭辰涛,唐隆晨,林 琪*
(1.福建省水产研究所、福建省海洋生物增养殖与高值化利用重点实验室,福建 厦门361013;2.上海海洋大学、水产种质资源发掘与利用教育部重点实验室,上海 201306;3.海洋生物种业技术国家地方联合工程研究中心,福建 厦门 361013)
光是植物赖以生存的重要环境因子,不仅作为光合作用的能量来源,还参与调控植物的生长与发育,光的光谱组分(光质)是光因子的一种重要属性[1]。大部分藻类植物生长在水环境中,由于水层深度和水体组分差异造成了水体对不同波长光的选择性折射和吸收,不同水体的光质组分存在差异[2-3]。为适应水体环境的光谱组分,藻类在漫长的进化过程中获得了多样的光质适应机制[4]。大量研究指出,可见光区的不同光质(白、蓝、红、绿光)可对藻类的光合效率、形态建成、生长发育[5-7]、细胞结构[8]以及生化代谢[9-15]等多个方面产生显著的影响。例如,蓝光可提高硬石莼(Ulvarigida)、长珊瑚藻(Corallinaelongata)和海头红(Plocamiumcartilagineum)[10]以及角刺藻(Chaetocerossp.)[11]的叶绿素含量,促进真江蓠(Gracilariavermiculophylla)的氨基酸合成[12]以及海带(Saccharinajaponica)幼孢子体的生长、细胞分裂[6]和卵母细胞形成[7];红光处理可促进长茎葡萄蕨藻(Caulerpalentillifera)增重[13];绿光处理可提高龙须菜(Gracilariopsislemaneiformis)相对生长率和光合效率[5],并有利于紫球藻(Porphyridiumcruentum)[9]和海膜(Halymeniafloresii)[14]的生长及光合色素积累。另外,藻类具有丰富迥异的生活史形式,有些藻类在不同生活史阶段对光质的响应也存在差异,例如,蓝光可促进小球藻(Chlorella)突变体休眠细胞对氨基酸的吸收,但却抑制了生长期细胞对氨基酸的吸收[15]。
坛紫菜(Pyropiahaitanensis)是一种具异形世代交替生活史的大型海洋红藻,其果孢子与壳孢子是实现世代交替的关键发育细胞[16]。目前,尽管关于不同光质对紫菜丝状体及叶状体的影响已有报道[8,17-22],但光质是否在坛紫菜孢子萌发过程中发挥作用目前尚不太清楚。因此,本研究以坛紫菜的果孢子和壳孢子为实验对象,探讨白、蓝、红、绿光4种光质对坛紫菜果孢子和壳孢子萌发的影响,旨在为探究坛紫菜对光质的适应性响应提供理论参考。
1 材料与方法
1.1 实验材料
实验所用果孢子由福建省霞浦海区养殖筏架上具成熟果孢子囊的坛紫菜叶状体经阴干刺激后放散获得[23],壳孢子由实验室内保存的坛紫菜纯系丝状体经扩培促熟后放散获得[24]。
1.2 实验条件设定
实验所用组合光源(白光)及单色光源(蓝、红、绿光)由功率为11 W的发光二极管(light emitting diode,LED)提供,其波长范围分别为:白光400~760 nm、蓝光455~475 nm、红光580~630 nm、绿光510~550 nm。通过增减LED灯管的数量及其与实验材料的距离对光量子通量密度进行调节,并采用PLA-20光谱分析仪测量实验光源的波长与光量子通量密度,培养果孢子和壳孢子的光量子通量密度分别为(15±1) μmol photons/(m2·s)和(40±2 ) μmol photons/(m2·s);温度与光周期均统一设置为(22±1)℃、10L∶14D。
1.3 孢子的培养与摄片记录
实验使用的培养液均为MES(modified ES)培养基[25]。将含有果孢子的海水经过200目筛绢滤去杂质后搅拌均匀,取等体积果孢子悬浊液于无色培养皿(Φ=9 cm)中黑暗静置12 h,待孢子附着后分别置于不同光质的光源下培养,每个光质组设3个生物学重复。每3 d更换1/3培养液,并用Leica DMI8倒置显微镜进行观察、摄片,随机统计20个果孢子萌发体的萌发管分枝数目及萌发管总长度。壳孢子的培养与摄片方法同上,培养过程中统计60个壳孢子萌发体的细胞数目。另外,在培养皿中随机选取30个视野(Φ=2 mm),统计区域内的孢子数量,在不同光质下培养15 d后,再次以同样的方法统计随机30个视野的孢子数量,并计算孢子的存活率。
1.4 数据处理与分析
采用图像测量软件Digimizer V4.3.4测量果孢子萌发管总长度,Excel 2013进行数据整理与统计,Origin 9.0作图,IBM SPSS 19进行实验组间的单因素ANOVA检验,设置差异显著水平为P<0.05。
2 结果
2.1 不同光质对坛紫菜果孢子萌发的影响
2.1.1 果孢子萌发的细胞学观察
如图1所示,果孢子经12 h避光附着后,可见细胞内具明显的凝集状黄褐色色素体[图1(a)、(g)、(m)、(s)]。遇光后的果孢子迅速萌发,并很快在各光质处理组间出现颜色差异。在白光与蓝光下的果孢子萌发体始终色泽饱满,呈鲜红色[图1(b)至(f)、(h)至(l)],而经红、绿光培养3 d的果孢子萌发体已呈黄褐色,并随着实验的进行,其色泽愈发暗淡,与白光组和蓝光组的萌发体颜色形成鲜明对比,最终呈现浅黄褐色[图1(n)至(r)、(t)至(x)]。在培养后期观察到红光组萌发体的萌发管明显较其他光质组少而短[图1(q)、(r)],而绿光组的萌发体虽然色泽与红光组类似,呈暗淡的浅黄褐色,但其萌发管较其他光质组更长,且具更多的分枝[图1(w)、(x)]。
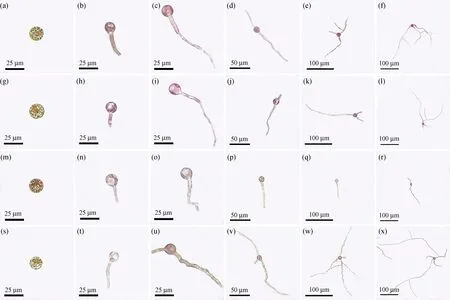
图1 不同光质下培养的坛紫菜果孢子的形态变化Fig.1 Morphogenetic developments of carpospores of P. haitanensis cultured under different light qualities(a)至(f)分别为白光下培养0、3、6、9、12、15 d的坛紫菜果孢子和萌发体;(g)至(l)分别为蓝光下培养0、3、6、9、12、15 d的坛紫菜果孢子和萌发体;(m)至(r)分别为红光下培养0、3、6、9、12、15 d的坛紫菜果孢子和萌发体;(s)至(x)分别为绿光下培养0、3、6、9、12、15 d的坛紫菜果孢子和萌发体。
2.1.2 萌发管分枝形成的时序
对果孢子萌发体萌发管分枝形成时序的量化统计结果显示,绿光下果孢子萌发管的分枝形成速度最快,白光下次之,蓝光下再次之,红光下最慢。在培养至第3天时,红光组萌发体的平均萌发管分枝数目已显著低于白光对照组(P<0.05),其余各组间未形成显著差异(P>0.05)。在培养进行至第9天时,各组间平均萌发管分枝数目开始出现较明显的差异,其中绿光组最多,白光组次之,红光组、蓝光组则显著低于其余各组(P<0.05),二者间暂无显著差异(P>0.05)。从第12天起至培养结束,各光质组萌发体的平均萌发管分枝数目表现出绿光组>白光组>蓝光组>红光组,且在各组间差异显著(P<0.05)。其结果如图2所示。
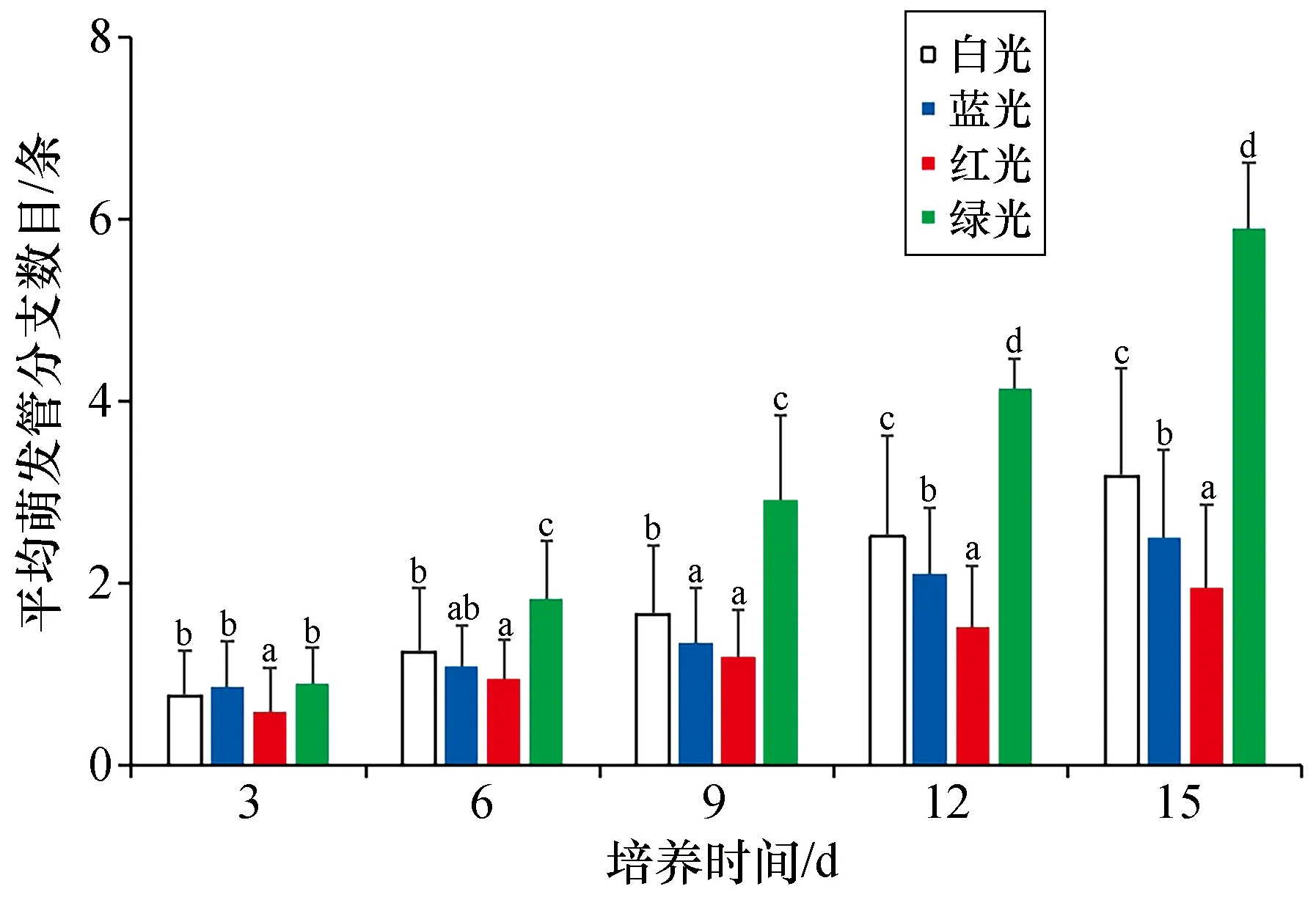
图2 不同光质下培养的坛紫菜果孢子的平均萌发管分枝数目Fig.2 Average numbers of germ tube branches of carpospores of P. haitanensis cultured under different light qualitiesn=20,不同字母代表同一时间内组间差异显著(P<0.05),下同。
2.1.3 萌发管长度比较
与萌发管分枝数目相似,在不同光质条件培养下果孢子萌发体的萌发管长度也存在较明显的差异。在培养至第3天时,绿光组萌发体的平均萌发管长度已开始明显大于其他光质组(P<0.05),其余各组间则无显著差异(P>0.05)。从第6天起,红光组萌发体的平均萌发管长度显著小于白光组(P<0.05),蓝光组则与白光组之间暂未形成显著差异(P>0.05),但从第9天起至培养结束,蓝光组萌发体的平均萌发管长度一直处于略大于红光组,但与之无显著差异的水平(P>0.05),而二者均显著小于白光组(P<0.05),并呈现出绿光组>白光组>蓝光组>红光组(P<0.05)。其结果如图3所示。

图3 不同光质下培养的坛紫菜果孢子萌发体的平均萌发管长度Fig.3 Mean lengths of germ tubes of carpospore germlings of P. haitanensis cultured under different light qualities n=20。
2.1.4 果孢子存活率
在白、蓝、红、绿光4种不同光质下培养15 d后的坛紫菜果孢子其存活率分别为87.53%、88.28%、82.19%和89.68%,红光组的坛紫菜果孢子的存活率略低于其他3个光质组,但各组间无显著差异(P>0.05),其结果如图4所示。

图4 不同光质下培养15 d的坛紫菜果孢子的存活率Fig.4 Survival rates of carpospores of P. haitanensis cultured under different light qualities for 15 days n=30。
2.2 不同光质对坛紫菜壳孢子萌发的影响
2.2.1 壳孢子萌发的细胞学观察
萌发前的坛紫菜壳孢子通常呈球状,深黄褐色或暗橙黄色(图5)。在培养初期,各光质组的壳孢子萌发过程相似,细胞发生横向分裂,并形成明显的假根。培养至第6天时,蓝光组的萌发体细胞颜色开始逐渐变红[图5(i)];而培养至第9天时,各光质条件下的萌发体色泽出现明显差异,蓝光组的萌发体细胞颜色加深并呈现鲜红色[图5(j)],白光组则呈现红棕色[图5(d)],而红、绿光组仍然保持暗淡的黄褐色[图5(p)、(v)]。培养至第15天时,各组中以蓝光组的萌发体颜色最深,呈深紫红色[图5(k)、(l)],白光组次之,呈鲜红色[图5(e)、(f)],红光组和绿光组萌发体均呈暗淡的浅黄褐色,且红光组色泽较绿光组稍浅[图5(q)、(r)、(w)、(x)]。

图5 不同光质下培养的坛紫菜壳孢子萌发过程的形态变化Fig.5 Germination morphogenesis of P. haitanensis conchospores cultured under different light qualities(a)至(f)分别代表白光下培养0、3、6、9、12、15 d的坛紫菜壳孢子和萌发体;(g)至(l)分别代表蓝光下培养0、3、6、9、12、15 d的坛紫菜壳孢子和萌发体;(m)至(r)分别代表红光下培养0、3、6、9、12、15 d的坛紫壳孢子和萌发体;(s)至(x)分别代表绿光下培养0、3、6、9、12、15 d的坛紫菜壳孢子和萌发体。
2.2.2 壳孢子的发育时序
与白光组相比,绿光组的壳孢子萌发体细胞分裂较快,而蓝光组和红光组则相对较慢。在前9天培养过程中,4种光质条件下培养的壳孢子萌发体的平均细胞数目未出现显著差异 (P>0.05)。但培养至第12天时,与白光组相比,蓝光组和红光组萌发体的平均细胞数目明显偏少(P<0.05),而绿光组则显著高于其余光质组(P<0.05)。培养至第15天时,蓝光组萌发体的平均细胞数目仍显著低于白光对照组(P<0.05),绿光组萌发体的细胞数目则仍保持显著最高的水平(P<0.05)。其结果如图6所示。
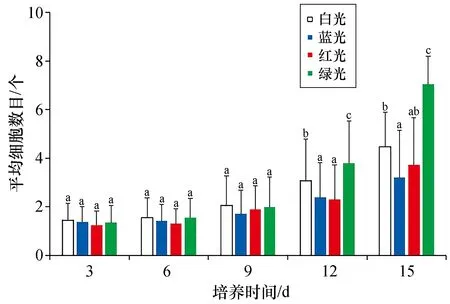
图6 不同光质下培养的坛紫菜壳孢子萌发体的平均细胞数目Fig.6 Average cell numbers of conchospore germlings of P. haitanensis cultured under different light qualities n=60。
2.2.3 壳孢子存活率
蓝、红、绿3种单色光下培养的壳孢子在第15天时的存活率分别为91.62%、93.03%和91.99%,均略低于白光组(96.71%),但各光质组间无显著差异(P>0.05)。其结果如图7所示。
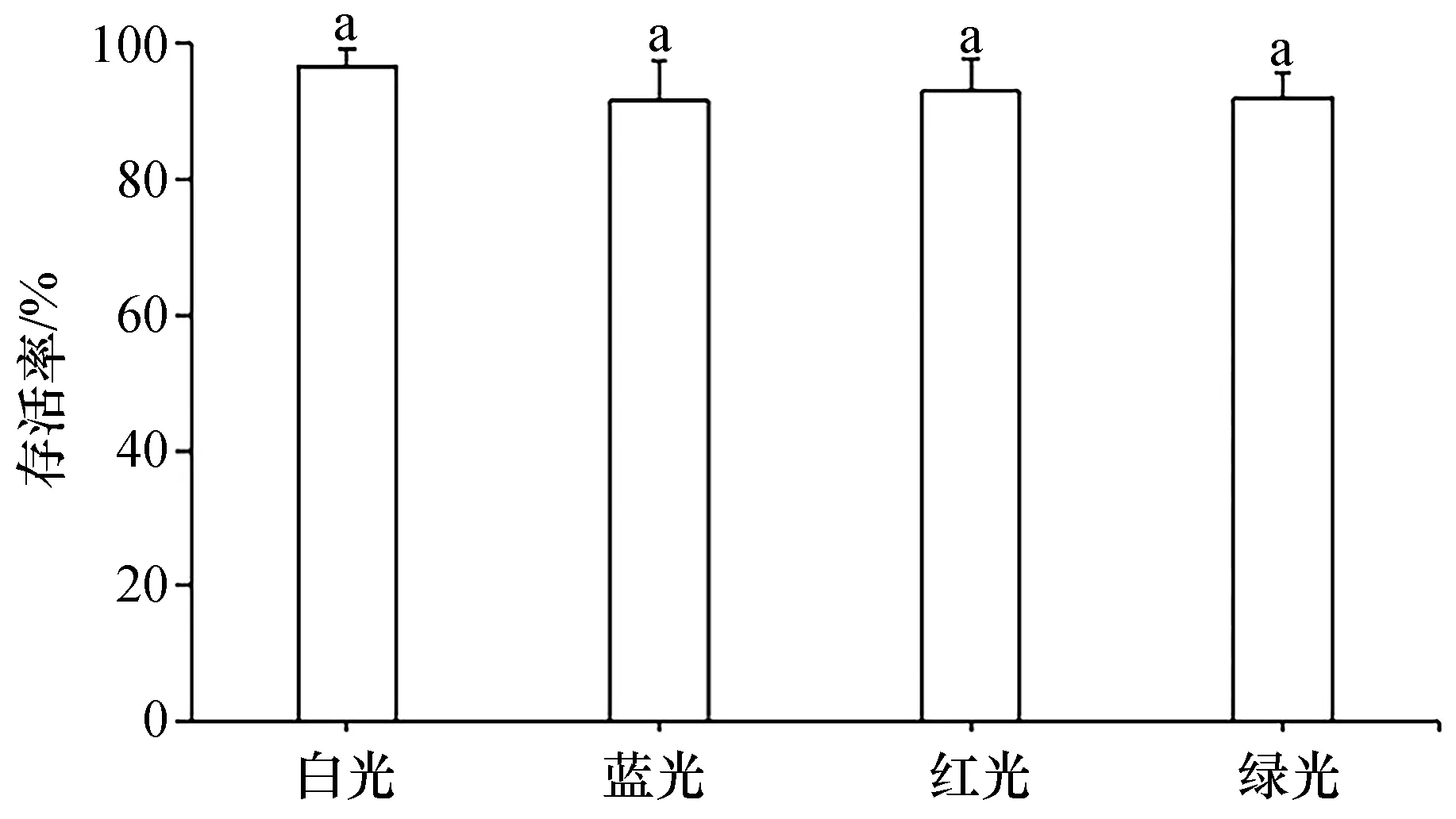
图7 不同光质下培养15 d的坛紫菜壳孢子的存活率Fig.7 Survival rates of conchospores of P. haitanensis cultured under different light qualities for 15 days n=30。
3 讨论
水对不同波段可见光的吸收具有明显的选择性[2-3]。受所处水层深度及水体成分的影响,不同藻类生境的光谱组分存在明显差异,因此,不同种类的海洋藻类在长期的适应性进化过程中对光质变化表现出丰富的响应模式[4,26]。在本研究中,不同光质下培养的坛紫菜果孢子和壳孢子均能萌发,但萌发速度与细胞颜色表现出明显差异,这表明不同波长的光在坛紫菜孢子的生长发育中不仅能够充当能量供应的角色来驱动光合作用的进行,还能影响孢子早期萌发时的形态建成和生长发育。
光合色素是藻类吸收光能的重要物质基础,包括叶绿素、藻胆素、类胡萝卜素等,这些色素的含量和组成决定了藻体的颜色[27],并可随着藻体对外界环境的光谱组分的适应而发生改变[10,28]。海藻所含光合色素种类、含量及其吸收光谱特性决定了它们对不同波段光的利用势必是不同的[29]。本研究观察到绿光与红光下的萌发体细胞出现明显的颜色变浅,而蓝光下的细胞颜色明显加深,这与坛紫菜丝状体和叶状体在这两种光质下的藻体色泽变化相类似,说明红光与绿光可能不利于藻胆色素的积累,蓝光则反之[17-18]。单一的红光或绿光照射下萌发体的色泽消退可能与天线蛋白的解耦和藻胆素的降解有关,这是由于紫菜的天线色素藻胆素对红绿光具有很强的吸收作用[28],光合作用的过量激发能会诱导产生有害的自由基,因此,天线蛋白解耦和藻胆素降解对避免这种过激是十分有利的[30-31]。相反,有研究表明脐形紫菜 (Porphyraumbilicalis)叶状体对蓝光的吸收作用较弱[21],在单一蓝光照射下细胞光合作用的光能捕获能力十分有限,但其适应性地提高了与光系统Ⅱ(PSⅡ)紧密相关的藻胆素的含量,这可能对维持藻体稳定的光合作用具有重要意义。目前,长期的红光或绿光照射已被证实不利于坛紫菜叶状体和丝状体的生长[17-18],且绿光还会抑制坛紫菜壳孢囊枝细胞的形成[32],但本研究中绿光照射下的孢子却表现出比白光下孢子的萌发速度更快的现象,这与坛紫菜雌性配子体在绿光下能够最早形成生殖细胞并促进单性生殖孢子的发生与萌发相似[18]。事实上,绿光已被发现能以最低光照强度促使多种红藻孢子的萌发[33],一些富含藻红素的红藻和蓝藻种类也表现出对绿光的利用效率比其他光质更高效的现象[34-36]。本研究中,绿光下培养的坛紫菜果孢子、壳孢子及其萌发体生长速度明显优于其他光质条件下的生长速度,暗示坛紫菜可能对绿光具有较高的利用效率。然而目前有关紫菜对不同光质光源的适应性响应机制还知之甚少,还有待进一步研究。
4 结论
光质显著影响坛紫菜果孢子和壳孢子萌发过程中细胞的色泽变化。白光和蓝光能使果孢子和壳孢子的萌发体颜色加深,最终变为鲜红色或深紫红色,红光和绿光则使两者颜色变浅,最终呈浅黄褐色。绿光能明显促进坛紫菜孢子萌发体的生长,暗示坛紫菜可能对绿光具有更高的利用效率。
