金门岛红树林湿地海洋线虫群落结构的初步探究
2023-08-09肖月平郭玉清施宜佳
肖月平,郭玉清,施宜佳
(集美大学水产学院,福建 厦门 361021)
小型底栖动物是一类数量多,分布广、生活周期短的后生动物,分选时能透过500 μm孔径的套筛而被42 μm孔径的套筛所截留,是一类评估海岸湿地中海洋沉积环境质量状况的重要指示生物[1]。海洋线虫是小型底栖动物中数量最多、分布最广泛的类群,通常占小型底栖动物数量的60%以上[2-3],是沿海红树林湿地底栖生态系统中重要的组成部分。在红树林碎屑食物链中,海洋线虫不仅是消费者,还在分解矿化有机碎屑方面扮演了重要的角色[4-5]。
金门岛地处厦门湾内,面积为124 km2,俯瞰形同哑铃。1999年12月,在古宁头西北海域设立了中国鲎保护区,保证了鲎幼体在潮间带的孵化率和成长率[6-7]。小型底栖动物是海洋碎屑食物链重要的中间环节,而海洋线虫作为其优势类群[8],为中国鲎提供了丰富的饵料资源[9-10]。迄今为止,金门红树林湿地底栖动物及其环境因子的研究暂无相关报道。2023年,本研究在金门岛发现了Parodontophora属的1新种[11]。本研究对金门岛海洋线虫群落,特别是优势属和摄食类型进行相关分析,以期为金门岛红树林湿地的生态系统健康评价提供数据参考。
1 材料与方法
1.1 站位设置与野外采样
2021年2月,在金门岛设置两个采样断面(图1),包括建功屿断面(24°25′12″N—24°25′38.28″N,118°18′3.24″E—118°18′38.52″E)和古宁头断面(24°29′9.24″N,118°18′15.84″E)。建功屿断面根据高、中、低潮位位设置H、M和L 3个站位,分别记为JH(24°25′38.28″N,118°18′38.52″E)、JM(24°25′37.2″N,118°18′34.92″E)、JL(24°25′12″N,118°18′3.24″E);古宁头断面仅在高潮位仅设置1个站位,记为GH(24°29′9.24″N,118°18′15.84″E)。每个站位只采集一个混合样。选择底质类型相对一致、无扰动的地点进行沉积物样品的采集,采样深度为0~5 cm。样品封装前倒入5%的福尔马林摇匀固定,然后常温保存带回实验室进行分析。采样均在白骨壤红树林区进行。
图1 采样站点Fig.1 Sampling station
1.2 样品处理
小型底栖动物室内分选及海洋线虫的制片方法见文献[12-13]。将野外采集获得的泥样放入由500 μm与42 μm孔径网筛组成的套筛内,用过滤后的自来水冲洗,直至透过套筛的水流清澈。将截留下的样品,用Ludox-TM 硅胶溶液(密度为1.15 g/mL)转入离心管,以4 000 r/min转速离心 10 min,重复两次,提取上清液。利用解剖镜(Nikon SMZ800)进行类群鉴定,利用微分干涉显微镜(Nikon ECLI-PSE-80i)进行海洋线虫的鉴定,鉴定主要参照文献[13-14]和https://nemys.ugent.be/数据库上的文献。
根据海洋线虫口腔类型和摄食方式,将其划分为4个功能类群(摄食类型)[15-16],即:选择性沉积食性者(selective deposit feeders,1A型);非选择性沉积食性者(non-selective deposit feeders,1B型);刮食者或底上硅藻食性者(epigrowth feeders,2A型);捕食者/杂食者(omnivores/predators,2B型)。
2 结果与讨论
2.1 小型底栖动物类群组成
研究区小型底栖动物类群组成及占比见表1。2个断面共发现10个小型底栖动物类群,包括海洋线虫、桡足类、寡毛类、多毛类、涡虫、有孔虫、腹足类、涟虫、双壳类和缓步类。海洋线虫为最优势类群,占小型底栖动物总数的93.93%,其次桡足类占5.31%,其他类群占0.75%。腹足类仅在建功屿断面JH、JL站位出现,占0.10%;缓步类仅在建功屿JM站位出现,占0.02%;有孔虫、涟虫和双壳类仅在古宁头断面GH站位出现,占0.05%。从不同站位小型底栖动物类群数来看,古宁头GH站位小型底栖动物类群数最多,而建功屿JL站位最少。海洋线虫在建功屿JM站位占比最高,达96.03%;其次为建功屿JL站位和古宁头GH站位;在建功屿JH站位占比最低(88.74%)。
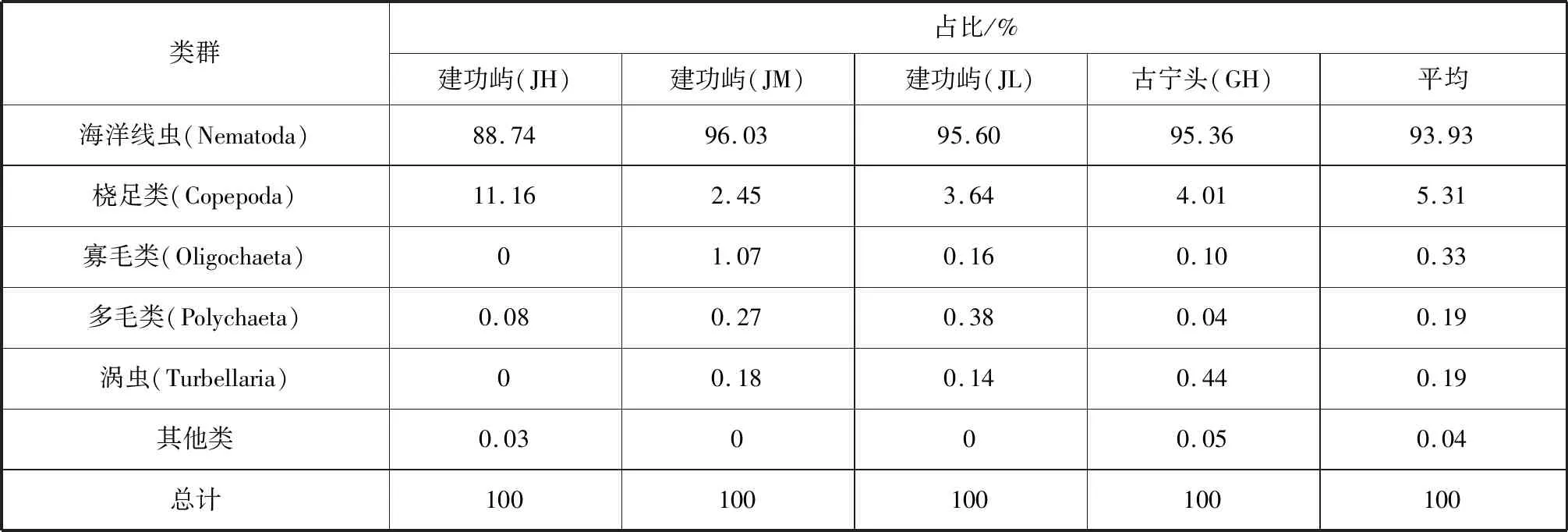
表1 金门红树林湿地各采样站位小型底栖动物类群占比Tab.1 Percentage of meiofaunal taxa at each sampling site in Jinmen mangrove wetland
本次金门岛小型底栖动物调查研究结果与厦门浔江湾小型底栖生物的研究结果较一致,浔江湾海洋线虫为最优势类群,占比为89.28%,其次为底栖桡足类,占比为2.76%[17]。海洋线虫占绝对优势,小型底栖动物为10个类群这一结果,也与厦门同安湾下潭尾红树林(人工林)、泉州洛阳江口红树林、广西防城港东湾红树林的调查结果相似[18-20]。
2.2 海洋线虫优势属(种)
本次调查共鉴定海洋线虫51种,隶属于4目15科31属。其中Daptonema为最优势属,占39.80%(表2);占比大于5%的优势属依次为Viscosia(11.02%)、Ptycholaimellus(9.62%)、Dorylaimopsis(9.54%)和Metachromadora(7.58%)。4个采样站位中,仅Daptonema为共有优势属,且在建功屿JL站位占比最高(72.61%),在JH、JM站位占比相近,不超过20%;在古宁头GH站位占比为51.34%。Anoplostoma仅在建功屿JH站位为优势属,Metachromadora仅在建功屿JM站位为优势属,Spilophorella仅在古宁头GH站位为优势属。比较2个断面的高潮位站位(JH、GH)站位,发现Ptycholaimellus、Viscosia、Daptonema和Parasphaerolaimus为共有优势属。
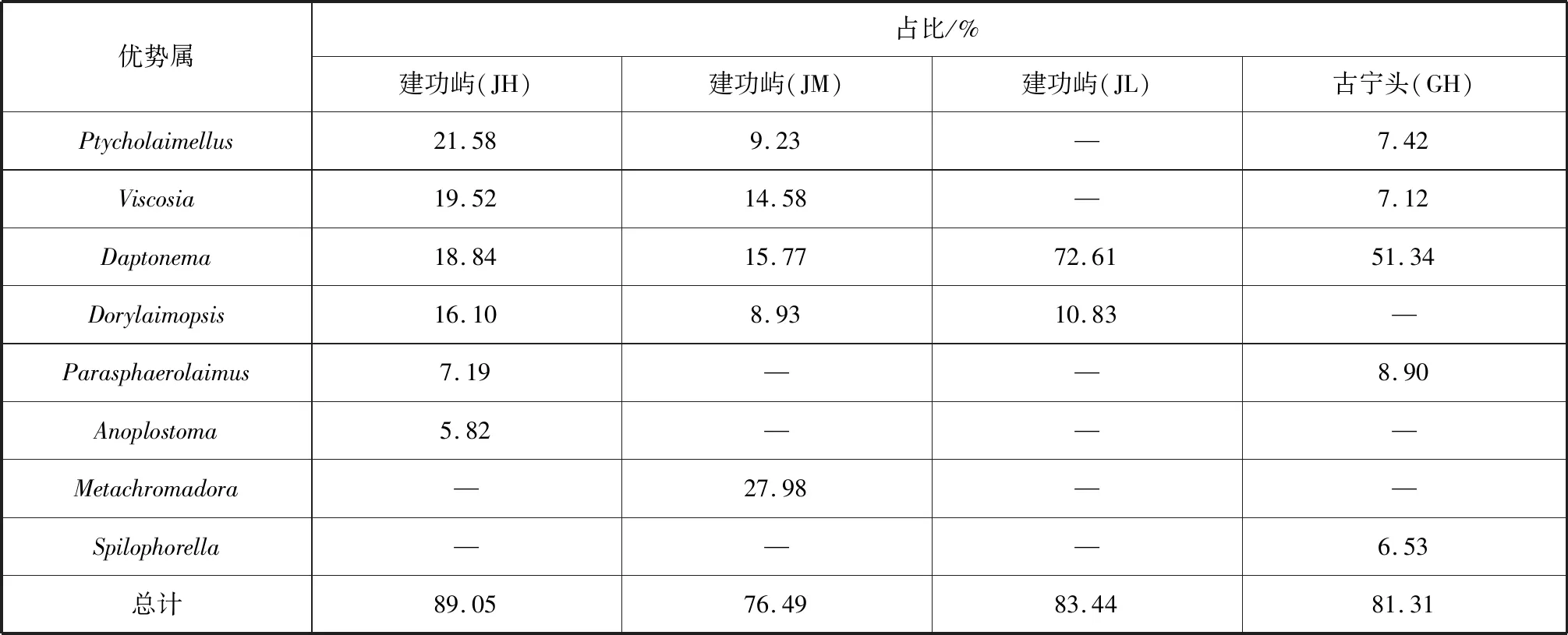
表2 金门红树林湿地海洋线虫优势属及其占比Tab.2 Dominant genera and dominance of marine nematodes in Jinmen mangrove wetland
4个站位中,Daptonemaquattuor的占比最高(14.78%),占比大于5%的优势种依次为Daptonemabaihaijiaoensis(12.74%)、Viscosiasp.1(10.95%)、Dorylaimopsissp.1(9.54%)、Ptycholaimellussp.1(7.97%)、Daptonemaungula(7.43%)和Metachromadorasp.1(7.35%),见表3。Ptycholaimellussp.1仅在2个断面高潮位站位为优势种,Ptycholaimellusocellatus和Metachromadorasp.1仅在建功屿JM站位为优势种,Ptycholaimellussp.1仅在建功屿JH站位为优势种,Dorylaimopsissp.1在建功屿3个站位为优势种,Anoplostomapapaviviparum仅在建功屿JH站位为优势种。

表3 金门红树林湿地海洋线虫优势种及其占比Tab.3 Dominant species and dominance of marine nematodes in Jinmen mangrove wetland
4个采样站位共有优势属仅Daptonema,但无共有优势种。表明各站位之间海洋线虫群落结构具有一定的相似性,但也存在差异,这可能与各站位所对应的沉积物类型有关。比较2个断面的高潮位站位发现,Ptycholaimellus、Viscosia、Daptonema和Parasphaerolaimus为共有优势属,而共有优势种仅Ptycholaimellussp.1和Viscosiasp.1;此外,古宁头GH站位不论在属还是种的水平上,其多样性都远超建功屿JH站位。由于缺乏相关环境因子的研究,2个断面高潮位站位海洋线虫群落差异原因尚不明确。
前人对福建5片红树林湿地海洋线虫的调查中发现Sabatiera、Ptycholaimellus、Parasphaerolaimus、Terschellingia、Daptonema、Viscosia和Dichromadora为优势属[21];洛阳江口红树林湿地中Sabatieria、Parasphaerolaimus、Adoncholaimus、Parodontophora、Terschellingia和Viscosia为优势属[20];海坛岛长江澳沙滩中Metachromadora和Viscosia也为优势属[1]。这与本研究结果有一定的相似性。Metachromadora和Viscosia不仅在粉砂质泥为主的沉积环境中为优势属,在沙滩中也具有较高的占比。Al-Sofyani等认为每个特定的栖息地海洋线虫没有特定的优势属[22],也就意味着海洋线虫的分布模式可能不会受到不同生境的影响。邹朝中在厦门岛附近发现了Dorylaimopsis属的一新种,但在形态上与本研究所发现的Dorylaimopsissp.1不同,因此认为其是同一属内的不同种[23]。
Cobb于1920年建立了Daptonema属[24]。目前,Daptonema在nemys数据库中共记录有228种,其中有效种145种,疑问种25种,拼写或分类错误58种。该属分类地位尚不清楚,易与TheristusBastian,1865[25]、ParamonohysteraSteiner,1916[26]、PromonhysteraWieser,1956[27]、ParagnomoxyalaJiang &Huang,2015[28]等属混淆。Daptonema属中有41种从Theristus转移而来。本研究在金门岛红树林湿地发现Daptonema属的6个种,其中仅有2种发现雌性,未能给出种名,另外4种已确定种名。
Daptonema属具有机质污染指示作用,该属的线虫对有机质污染具有一定的耐受能力,其丰度和优势度会随环境中有机质含量的增加而增加[29-30]。其相对丰度大于10% 即表示生态质量状况较差或差[30-31]。Daptonema属在本研究中占绝对优势,总占比为39.80%。在建功屿断面JL站位,Daptonema属的个体数量占比超过70%,而在JH、JM站位占比均不足20%,表明建功屿断面JH站位沉积物有机质污染可能较严重,原因有待考究。另外,建功屿JH站位Daptonema属占比较古宁头断面GH站位低,表明后者受有机质污染可能更严重。总体而言,金门建功屿和古宁头红树林湿地可能面临严峻的有机质污染威胁,生态质量状况较差[6]。
2.3 海洋线虫的摄食类型
本研究中,海洋线虫群落以非选择性沉积食性者(1B型)占优,有18种,其次刮食者或底上硅藻食性者(2A型)和捕食者/杂食者(2B型)分别有12种和11种,选择性沉积食性者(1A型)10种。表明1B型在研究区域占绝对优势,2A型2B型居中,1A型为最少。从摄食类型在该站位摄食类群的占比看(表4),4个站位中除建功屿JM站位以2A型为主,其他3个采样站位皆以1B型占绝对优势;且1B型在建功屿JL站位占比最为突出(85.35%)。
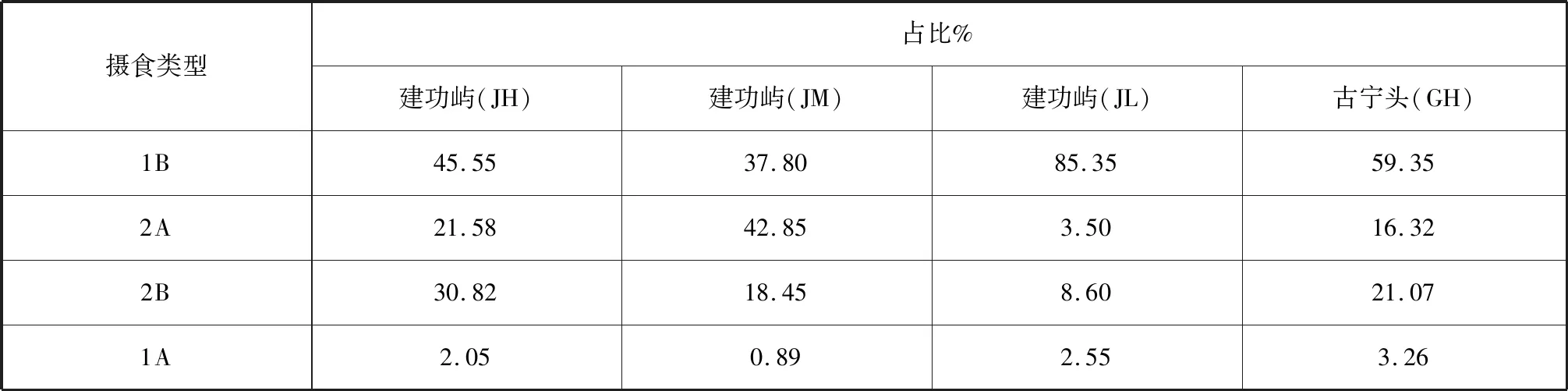
表4 金门红树林湿地海洋线虫的摄食类型及其占比Tab.4 Feeding types and percentage of marine nematodes in Jinmen mangrove wetland
海洋线虫几乎能利用周围环境中所有的营养源,如细菌、有机碎屑、微藻、真菌、微型底栖生物和其他小型底栖动物[32]。海洋线虫摄食类型与所处的沉积物类型密不可分,其摄食类型的变化反映了沉积物中线虫食物来源的不同,反映了沉积物中有机物存在形式的不同[33]。Tietjen也认为,食物的可获得性与沉积物类型之间存在密切关系[34]。Nguyen在越南Can Gio红树林湿地发现海洋线虫摄食类型以1B型为优势,其次为1A型[35]。杨培培等在漳江口、九龙江口和凤林等红树林湿地发现海洋线虫均以1B型占主导,而在洛阳江口和湾坞红树林湿地中以2B和2A型为优,1A型为最少[21]。这与本研究结果相似,可能是金门红树林有机质含量较高,海洋线虫主要以腐烂的有机碎屑为食。
3 结论
本次金门岛红树林湿地小型底栖动物研究中发现小型底栖动物共10个类群,海洋线虫占绝对优势(93.93%)。共鉴定出海洋线虫4目15科31属51种,其中优势属(占比从高到低)依次为Daptonema、Viscosia、Ptycholaimellus、Dorylaimopsis和Metachromadora;优势种为Daptonemasp.2、Daptonemasp.1、Viscosiasp.1、Dorylaimopsissp.1、Ptycholaimellussp.1、Daptonemasp.3和Metachromadorasp.1。4个站位中共发现10个共有种,分别为:Daptonemasp.1、Daptonemasp.2、Daptonemasp.3、Daptonemasp.4、Ptycholaimellussp.1、Viscosiasp.1、Dorylaimopsissp.1、Sabatieriasp.1、Parasphaerolaimussp.4和Sphaerolaimussp.3。摄食类型占比从高到低依次为1B>2A>2B>1A。
