电流刺激对钝齿棒杆菌厌氧条件下代谢通量分布的影响
2023-08-05陈小举张凤琴
陈小举,张凤琴
(巢湖学院生物与环境工程学院,安徽 巢湖 238000)
在食品、药品等行业中,琥珀酸与乳酸及其衍生物有着广泛应用。以琥珀酸与乳酸为前体可以合成具有广阔应用前景的可降解生物材料,因此,市场对琥珀酸与乳酸的需求旺盛。利用微生物发酵可再生资源生产琥珀酸与乳酸被认为是减少石化资源消耗和减少碳排放的有效手段,也是最具发展潜力的绿色工艺模式之一[1-4]。
钝齿棒杆菌(Corynebacterium crenatum)是生产精氨酸、谷氨酸等氨基酸的主要菌株之一。钝齿棒杆菌产氨基酸的发酵工艺多是有氧液体深层发酵,且对溶氧水平有较严格的要求[5]。将钝齿棒杆菌培养环境改为无氧时,琥珀酸与乳酸是其主要代谢产物。研究发现,C4途径是钝齿棒杆菌厌氧合成琥珀酸的主要途径,即由磷酸烯醇式丙酮酸至草酰乙酸,然后经苹果酸与富马酸生成琥珀酸[6](图1)。理论上,转化1 mol磷酸烯醇式丙酮酸可以生成1 mol琥珀酸,并需要2 mol还原型烟酰胺腺嘌呤二核苷酸(reduced form of nicotinamide-adenine dinucleotid,NADH)。糖酵解途径(Embden Meyerhof pathway,EMP)是钝齿棒杆菌代谢葡萄糖的主要途径,此途径每生成1 mol磷酸烯醇式丙酮酸仅伴随生成1 mol NADH,仅能满足由草酰乙酸至苹果酸或者由富马酸至琥珀酸所需的NADH。因此,厌氧发酵时,钝齿棒杆菌将70%左右的葡萄糖转化为乳酸,而琥珀酸的代谢通量相对较低,不利于联产乳酸与琥珀酸[7-8]。

图1 钝齿棒杆菌在厌氧条件下的代谢网络Fig.1 Metabolic network of C.crenatum under anaerobic conditions
如果要提高琥珀酸生成比例,利用代谢调控增加琥珀酸生产菌胞内的H还原力水平是可行的方法。比如,Zheng Pu等[9]利用基因组改组技术增加Actinobacillus succiniogenes胞内NADH通量,将琥珀酸产量提高了73%;Zhou Zhihui等[10]发现表达外源pntAB基因可以明显增加谷氨酸棒杆菌胞内的NADH通量与琥珀酸产量。另一提高琥珀酸产量的有效措施是通过添加电子供体、NAD前体物质调控胞内NADH/NAD+值。如姜岷等[11]通过流加还原剂改变发酵培养基的氧化还原电位水平,实现了对A.succinogenes胞内NADH代谢的调控,将琥珀酸生产效率提高了57.3%[11]。
除以上方法外,电刺激发酵也可用于调控微生物胞内的NADH代谢。电刺激发酵就是将惰性电极插入微生物培养液中,形成一种电解池系统[12]。当施加电流后,在阴极会产生电子,电子通过传递进入微生物胞内可以提高NADH/NAD+值[13]。电压达到一定值后,阴极还会发生还原反应而产生氢气。产生的氢气可降低胞外氧化还原电位,并作为电子传递供体使胞内NADH/NAD+值升高。电刺激发酵技术在发酵产氢、丁醇、丙二醇、琥珀酸等高还原性代谢产物的工艺中得到了初步应用。已有研究发现,低压、低电流刺激可以提高厌氧发酵产氢的能力,其主要原因是微生物胞内NADH可用水平的增加[14-15]。与琥珀酸相似,NADH不足也是导致丁醇产量较低的主要原因[16]。He Aiyong等[17]发现电刺激发酵可以提高Clostridium beijerinckiiIB4胞内的NADH/NAD+值,并导致丁醇的产量及生产效率分别增加17.4%与60.3%。Zhou Mi等[18]发现施加电流刺激可以提高甘油对1,3-丙二醇的转化率,代谢流分析结果显示NADH通量增加是1,3-丙二醇产量升高的主要原因。Wang Zhen等[19]研究结果表明,对A.succinogenes发酵过程施加电流刺激可以将琥珀酸提高22.4%。
电化学技术在微生物发酵领域的应用为发酵过程控制及微生物代谢调控提供了新的研究方向[13,20],但电刺激对钝齿棒杆菌厌氧发酵特性的影响尚不明确,菌株胞内哪些节点的代谢流分布对电流刺激胁迫产生了应激响应尚不清楚。且目前为止,此方面的研究甚少。因此,本研究以钝齿棒杆菌为发酵菌株,通过改变厌氧发酵过程中的电流强度对胞内NADH代谢进行调控,以提高琥珀酸产量,并对不同电流条件下的代谢通量及基因表达水平进行分析,考察电刺激对钝齿棒杆菌代谢流分布的影响,找出可以联产乳酸与琥珀酸的电流刺激条件。本研究结果将有助于在理论方面掌握电流刺激对钝齿棒杆菌厌氧发酵特性的影响,在应用方面可建立提高琥珀酸生产效率的电刺激发酵工艺,同时对推动发酵工程与电化学学科的交叉融合也具有重要意义。
1 材料与方法
1.1 材料与试剂
1.1.1 菌株
钝齿棒杆菌CICC20219,目前由巢湖学院生物工程实验室保藏。
1.1.2 培养基
培养基A主要用于微生物的快速繁殖,主要成分:葡萄糖40 g/L,尿素2 g/L,酵母提取物2 g/L,酪蛋白氨基酸7 g/L,(NH4)2SO47 g/L,KH2PO40.5 g/L,K2HPO40.5 g/L,MgSO40.5 g/L,FeSO40.006 g/L,MnSO40.0042 g/L,生物素0.0002 g/L、硫胺素0.0002 g/L[21]。
培养基B 主要用于厌氧发酵产乳酸与琥珀酸,主要成分:葡萄糖60 g/ L,NaHCO316.8 g/ L,KH2PO40.5 g/ L,K2HPO40.5 g/L,MgSO40.5 g/L,FeSO40.006 g/L,MnSO40.0042 g/L,生物素0.0002 g/L、硫胺素0.0002 g/L[21]。
1.1.3 试剂盒
NADH与NAD+浓度检测试剂盒(产品序号:A114)、RNA提取试剂盒(产品序号:NO65)、cDNA合成试剂盒(产品序号:N118)南京建成生物工程研究所。
1.2 仪器与设备
ACQUTIY UPLC H-Class plus超高效液相色谱系统 美国Waters公司;FQD-96A实时荧光定量聚合酶链式反应(real-time polymerase chain reaction,real-time PCR)仪 杭州博日科技股份有限公司;CHI660E电化学工作站 上海辰华仪器有限公司;ACQUTIY UPLC HSS T3色谱柱(100 mm×2.1 mm,1.8 μm)、ACQUTIY UPLC BEH Amide Carbohydrate Analysis C18色谱柱(100 mm×2.1 mm,1.7 μm)爱尔兰Waters公司;全温摇瓶柜 苏州培英公司。
1.3 方法
1.3.1 实验设计
本研究采用的发酵工艺可分为2 个阶段:有氧阶段与厌氧阶段。有氧阶段主要用于钝齿棒杆菌的快速繁殖,目的是获得大量的菌体;厌氧阶段主要为代谢葡萄糖生产有机酸的发酵阶段。主要过程如下:挑取钝齿棒杆菌接种于含30 mL培养基A的250 mL摇瓶中,于30 ℃、200 r/min培养12 h,按照10%接种量转接于含100 mL培养基A的500 mL摇瓶中,然后于30 ℃、200 r/min培养10~12 h。将培养液于5000 r/min、4 ℃离心10 min,弃上清液并收集菌体,然后将菌体接种于厌氧发酵培养基B,接种量:生物量干质量浓度约为8 g/L。
厌氧发酵阶段使用100 mL的H型电解池为发酵容器(图2)。H型电解池为3电极体系,铂片电极(10 mm×10 mm×0.1 mm)为工作电极,Ag/AgCl电极为参比电极,且置Ag/AgCl电极于阴极电解池,并使用质子膜将阴极与阳极隔开。阳极与阴极电解池均含有80 mL培养基B,唯一不同的是:添加3 g碱式MgCO3于阴极电解池,用于中和产生的有机酸。电流通过电化学工作站提供,开环条件下的电解池系统为对照组,默认电流大小为0 mA。

图2 H型电解池发酵系统示意图Fig.2 Schematic view of H-type electrolytic fermentor
1.3.2 分析检测
细胞数量通过检测OD600nm和干质量进行评估。NADH与NAD+浓度由辅酶I检测试剂盒测定。利用超高效液相色谱对有机酸和残糖含量进行分析。检测有机酸时,采用二极管阵列检测器,色谱柱为ACQUTIY UPLC HSS T3色谱柱(100 mm×2.1 mm,1.8 μm),流动相为0.003 mmol/L硫酸溶液,流速为0.2 mL/min,检测波长为210 nm。检测残糖时,采用蒸发光检测器,色谱柱为ACQUTIY UPLC BEH Amide Carb ohydrate Analysis C18色谱柱,流动相为80%乙腈溶液,流速为0.4 mL/min。
1.3.3 基因表达水平分析
钝齿棒杆菌在厌氧条件下培养4 h后,利用TRIzol法提取总RNA,然后反转录得到cDNA,以16S RNA为内参基因,利用real-time PCR仪对6-磷酸葡萄糖脱氢酶基因(zwf)、磷酸烯醇式丙酮酸羧化酶基因(pepc)、苹果酸脱氢酶基因(mdh)及乳酸脱氢酶基因(ldhA)的表达水平进行分析。RNA提取与cDNA合成分别使用相关试剂盒完成。使用的引物如表1所示,引物合成委托生工生物工程(上海)股份有限公司完成。

表1 real-time PCR使用的特异性引物Table 1 Specific primer sequences used for real-time PCR
1.3.4 代谢通量分析
钝齿棒杆菌在厌氧条件下培养时,生物量基本没有变化[8,22]。因此,本研究不考虑生物量对代谢通量分布的影响。代谢通量计算过程中,假设胞内代谢反应的中间代谢产物处于拟稳态,并能及时转化为下游产物,同时设定NADH与NADPH可无限制相互转换。钝齿棒杆菌在厌氧条件的代谢网络如图1所示,主要由EMP、磷酸戊糖途径(pentose phosphate pathway,HMP)、琥珀酸、乳酸、乙酸等合成途径组成。由图3可知,代谢通量计算时共有27 个变量,围绕6-磷酸葡萄糖、6-磷酸果糖等代谢中间体可列出代谢通量方程21 个(表2),围绕NADH(NADPH)平衡、三磷酸腺苷(adenosine triphosphate,ATP)平衡可列出代谢通量方程2 个,即共有23 个代谢通量方程,方程自由度为4,因此,需要测定4 个变量才能对代谢通量方程进行求解。本研究取厌氧发酵4 h的发酵液进行代谢产物分析,通过离线精确检测残糖、乳酸、琥珀酸和乙酸等变量,以求解代谢通量方程。

表2 代谢通量方程Table 2 Equations of metabolic flux balance

图3 变量分布示意图Fig.3 Schematic diagram of variables
2 结果与分析
2.1 电流刺激对发酵结果的影响
本研究以H型电解池为发酵容器,在发酵过程中通过施加电流刺激改变钝齿棒杆菌胞外培养环境,研究电流刺激对其代谢特性的影响,结果如图4所示。研究发现,通入电流对钝齿棒杆菌在厌氧条件下发酵特性产生了较为明显的影响。在对照组(未通电),钝齿棒杆菌在8 h内共消耗葡萄糖58.7 g/L(图4a),葡萄糖消耗速率为7.3 g/(L·h)(图5)。施加-2 mA与-5 mA电流刺激后,在相同时间内,钝齿棒杆菌消耗的葡萄糖分别达到65.7 g/L(图4b)与74.7 g/L(图4c),葡萄糖消耗速率分别为8.2 g/(L·h)与9.3 g/(L·h),与对照组相比,分别增加12.3%与27.4%。以上结果说明,通入电流对葡萄糖的代谢效率具有显著促进作用。导致钝齿棒杆菌在电流刺激条件下糖代谢效率增加的可能原因是细胞膜通透性的提升。因为,钝齿棒杆菌吸收利用葡萄糖的过程主要分为跨膜运输与糖酵解,葡萄糖只有通过细胞膜进入胞内才能被微生物进一步利用,而研究显示适度的电流刺激会增加细胞膜的通透性[23]。另外,细胞膜通透性增加还会加速胞内代谢产物(琥珀酸与乳酸)向胞外的转运效率,减弱胞内代谢产物反馈抑制,提升胞内的整个代谢速率。与本研究结果相似,Wang Kai等[24]也发现,施加电流刺激可以增加硫酸盐还原细菌细胞膜的通透性,并加速底物与代谢产物的跨膜转运。

图4 不同电流刺激条件下的发酵结果Fig.4 Fermentation characteristics under different electrical current stimulation conditions

图5 不同电流刺激条件下的葡萄糖消耗速率Fig.5 Glucose consumption rates under different electrical current stimulation conditions
施加电流刺激后,发酵液中的乳酸与琥珀酸质量浓度均高于对照组。无电流刺激条件下,发酵液中的乳酸质量浓度在8 h达到43.9 g/L(图4a)。当电流为-2 mA与-5 mA时,乳酸质量浓度分别为45.0 g/L(图4b)与46.5 g/L(图4c),与对照组比,分别增加了2.5%和5.9%。对照组实验的乳酸质量浓度在3 组中虽然是最低的,但其乳酸产量(每代谢1 g葡萄糖产生的乳酸质量)最高,达到0.75 g/g(图6),分别比-2 mA与-5 mA实验组高出10.3%与21.0%。通入电流可以明显提高发酵液中琥珀酸质量浓度及琥珀酸产量,电流越小,琥珀酸质量浓度与产量越高。当电流由0 mA(对照组)变为-5 mA时,琥珀酸质量浓度由13.8 g/L增加到28.9 g/L,琥珀酸产量由0.24 g/g增加到0.39 g/g(图6),与对照组相比,质量浓度与产量分别增加了109.4%和62.5%。同时,琥珀酸与乳酸产量比由0.32增加到0.63。以上结果说明,电流刺激可以改变钝齿棒杆菌在厌氧条件下的代谢物组成,并有利于提高需要较多H还原力的代谢产物的合成,如琥珀酸。

图6 不同电流刺激条件下的有机酸产量Fig.6 Organic acid yields under different electrical current stimulation conditions
微生物培养过程中,向反应系统施加电流刺激可以在培养液中产生更多的电子,电子通过传递进入细胞会对胞内NADH代谢产生影响,继而影响微生物整体代谢特性[13,20,25],比如代谢产物通量分布。为了深入分析电流刺激调控钝齿棒杆菌代谢特性的原因,本研究对不同电流刺激条件下的胞内NADH/NAD+值进行分析。由图7 可以看出,通入电流后,钝齿棒杆菌胞内的NADH/NAD+值明显升高。当电流为-2 mA与-5 mA时,NADH/NAD+值分别为0.79与0.85,比对照组高出21.5%和30.8%。由图1可以看出,辅因子NADH/NAD+是6-磷酸葡萄糖脱氢酶、苹果酸脱氢酶、琥珀酸脱氢酶与乳酸脱氢酶等的辅酶,表明胞内H还原力可用水平对钝齿棒杆菌代谢通量分布有重要的调节作用。因此,电流刺激导致钝齿棒杆菌胞内NADH通量增加是导致钝齿棒杆菌代谢特性发生变化及琥珀酸产量增加的主要原因之一。与本研究结果相似,其他研究也表明改变微生物培养环境可以调节其胞内NADH的可用水平,随之调控胞内产物代谢通量分布[26]。
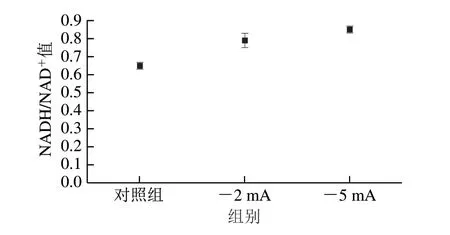
图7 不同电流刺激条件下的NADH/NAD+值Fig.7 NADH/NAD+ ratio under different electrical current stimulation conditions
除了乳酸与琥珀酸,乙酸是钝齿棒杆菌厌氧发酵时的主要副产物。施加电流刺激后,乙酸质量浓度也呈现上升趋势,由1.3 g/L(0 mA)增加到2.3 g/L(-5 mA),但其质量浓度与产量相对较低。
2.2 电流刺激对代谢通量分布的影响
NADH(或NADPH)主要由EMP与HMP产生。与EMP相比,1 分子葡萄糖经由HMP代谢后会产生额外2 分子H还原力(图1)。本研究中,施加电流刺激后,消耗单位葡萄糖产生的NADH明显增加,表明胞内EMP与HMP两个途径的代谢通量分布在施加电流刺激后发生改变。另外,磷酸烯醇式丙酮酸节点的代谢流分布对琥珀酸、乳酸产量也有重要调节作用[27]。通过代谢通量分析,可以更深入了解电流刺激对6-磷酸葡萄糖、磷酸烯醇式丙酮酸等关键节点代谢流分布的影响,有助于通过电流刺激调控琥珀酸与乳酸生成比,实现钝齿棒杆菌厌氧发酵联产乳酸与琥珀酸。代谢通量分析结果如表3所示。

表3 代谢通量计算结果Table 3 Calculated metabolic flux distribution
可以看出,除琥珀酸、乳酸、乙酸等最终产物外,电流刺激对6-磷酸葡萄糖、磷酸烯醇式丙酮酸等节点的代谢流分布产生了明显影响。对照组中,6-磷酸葡萄糖节点流向6-磷酸果糖的流量J2为 92.4 mmol/(L·g·h),至6-磷酸葡萄糖酸的流量J9为7.6 mmol/(L·g·h),两者比例为12∶1,说明碳源主要经由EMP 进行代谢。同时,在磷酸烯醇式丙酮酸节点,流向丙酮酸的流量J7为159.7 mmol/(L·g·h),流向草酰乙酸(C4途径)的流量J22为37.7 mmol/(L·g·h),两者比例约为4:1,说明碳代谢流主要流向丙酮酸-乳酸合成途径。施加-2 mA与-5 mA电流刺激后,J9分别增加到14.0、19.7 mmol/(L·g·h),与对照组相比,分别增加了84.4%和150.6%。同时,J7流量由 159.7 mmol/(L·g·h)减少到146.9、134.2 mmol/(L·g·h)。相比之下,J22由37.7 mmol/(L·g·h)增加到48.4、59.3 mmol/(L·g·h)。以上结果说明,施加电流刺激可以明显增加HMP与C4途径的代谢通量。其原因可能是:微生物胞内的氧化还原态势处于一个动态平衡状态,在钝齿棒杆菌胞内H还原力因外界环境改变而升高后,微生物会调节胞内的代谢产物组成,以合成还原态势更高的代谢产物,如琥珀酸,让NAD+快速重生,使胞内氧化还原态势处于新的稳定状态[7,28]。
2.3 电流刺激对基因表达水平的影响
为进一步明确电流刺激对钝齿棒杆菌代谢通量分布影响,本研究对关键酶基因的表达水平进行了分析。由图8可以看出,施加-5 mA电流刺激后,6-磷酸葡萄糖脱氢酶基因(zwf)、磷酸烯醇式丙酮酸羧化酶基因(pepc)与苹果酸脱氢酶基因(mdh)的表达水平均上调,而乳酸脱氢酶基因(ldhA)表达水平下调。zwf与pepc表达水平上调,可以进一步佐证电流刺激可以增加HMP与C4途径代谢通量的结果。苹果酸脱氢酶是琥珀酸合成途径中的关键酶,NADH是其辅酶[29]。mdh表达水平上调,ldhA表达水平下调,说明C4途径中草酰乙酸至苹果酸的代谢活动增加,丙酮酸至乳酸的代谢活动下降,与琥珀酸产量增加、乳酸产量减少的发酵结果一致。

图8 不同电流刺激条件下的基因表达水平Fig.8 Gene expression levels under different electrical current stimulation conditions
厌氧条件下,钝齿棒杆菌利用C4途径合成1 mol琥珀酸,需要2 mol NADH。EMP产生的NADH无法满足高琥珀酸合成通量对NADH的需求,而HMP则可产生更多的H还原力。在本研究中,施加电流刺激后,阴极发生还原反应产生电子,甚至析氢,并经过传递链进入钝齿棒杆菌胞内,电流刺激提供的额外电子致使6-磷酸葡萄糖节点处的HMP代谢通量增加,并合成更多的H还原力,结果导致胞内的NADH/NAD+值显著升高[25]。因为胞内可用于合成琥珀酸的NADH增加,钝齿棒杆菌调节了磷酸烯醇式丙酮酸节点的代谢流分布,使琥珀酸合成途径代谢通量增加,这可能就是施加电流刺激后,琥珀酸产量增加的主要原因。因此,6-磷酸葡萄糖是影响琥珀酸产量的关键节点,只有该节点出的代谢流发生改变,使更多的碳经由HMP进行代谢产生更多的H还原力,才能有效提高琥珀酸产量。
3 结论
研究电流刺激对钝齿棒杆菌厌氧发酵特性的影响,结果显示,施加-2 mA与-5 mA电流刺激可以提升钝齿棒杆菌代谢葡萄糖的速率与琥珀酸的产量,有利于联产乳酸与琥珀酸。电流刺激提供的额外电子使HMP代谢通量增加,并合成更多的H还原力,是琥珀酸产量提高的主要原因。本研究结果有利于加强发酵工程与电化学学科的交叉融合,可为利用微生物发酵工艺联产琥珀酸及乳酸提供参考。
