不同辣木种质资源对辣木瑙螟生长发育和繁殖的影响
2023-08-05桂华敏方雪峰张祖兵等
桂华敏 方雪峰 张祖兵等


关键词 辣木;种质资源;辣木瑙螟;生命表;生长发育
中图分类号:S 763.7 文献标识码:A DOI:10.16688/j.zwbh.2022364
辣木lVIoringa Adans.,又名鼓槌樹、奇迹树等[1],为辣木科Moringaceae辣木属Moringa多年生热带落叶乔木,其营养价值丰富,健康功效独特,可作为粮食、蔬菜,也可作为油料植物、蜜源植物、畜牧饲料、薪材、水质净化资源、化妆品及医疗用途[2-7]。辣木原产于印度,1910年引种到我国台湾,随后云南、广东、海南、广西、福建、贵州、四川等省份亦有引种[8]。目前我国已经引种栽培的有多油辣木M. oleifera(包括传统种,印度改良种‘PKM1’以及‘PKM1’的改良种‘PKM2’等品种)、狭瓣辣木M.stenopetala、北方辣木Mconcanensis[9]等,其中多油辣木的种植面积最大。2012年,辣木叶获得了新种质资源食品批准,并于2015年列入国家有机食品认证目录。近年来,随着人民生活水平逐渐提高,辣木作为新兴的食品[10],种植规模逐年扩大,目前我国的种植面积约为10万hm,其中云南种植面积超过4万hm,约占全国的40%[11-12]。
辣木瑙螟Noorda blitealis属鳞翅目Lepidop-tera草螟科Crambidae,是辣木的重要害虫之一[13]。印度首先报道了该虫为害辣木[4],2015年国内首次在云南省玉溪市元江县发现辣木瑙螟为害辣木,在辣木园调查样地中植株被害率达94%以上[15]。目前,辣木瑙螟在云南省的辣木种植区内均有发生,单株虫量最高可达302头[15]。辣木瑙螟为寡食性昆虫,辣木科辣木属下的13个种:lVIoringa arborea、M.borziana、北方辣木M.concanensis、象腿辣木M.drouhardii、M. hildebrandtii、长管辣木M. lon-gituba、多油辣木M.oleifera、M.ovaliolia、M.peregrina、M.pygmaea、M.rivae、M.ruspoliana、狭瓣辣木M.stenopetala均受其为害[16]。辣木瑙螟幼虫有5龄[17],一般为乳白色或浅绿色,老熟幼虫为粉红色。幼虫取食叶片时,吐丝将2~3片叶缀在一起或纵卷单叶吃成缺刻,为害严重时可将嫩叶全部吃光,并造成全株落叶,仅留光杆。当无幼嫩部位可取食时,幼虫可吐丝下垂取食辣木表皮或木质部[17]。幼虫发育成熟后下树入土,在土壤中化蛹、羽化,幼虫虫口数在垂直方向差异较大,其中上、中层虫口数较多,下层较少[18]。辣木瑙螟繁殖力强,产卵量大,成虫有较强的趋光性[19-21]。幼虫历期7~15d;蛹期4~9d;完成一个生命周期需16~26d[17]。辣木瑙螟为一种隐蔽性较强且世代重叠严重的害虫,在不同国家、地区和气候条件下,该虫的发生规律存在较大差异[22-25]。目前,辣木瑙螟的防治包括农业防治(选用抗虫品种、田间管理、间套作[26])、物理防治(人工捕杀、黑光灯诱杀[27])、药剂防治[28]和生物防治(天敌防治[29]、性诱剂诱杀[30])。
该虫的为害特点、生活习性、防治技术在部分国家和地区已有报道,但不同辣木资源对辣木瑙螟生长发育及繁殖的影响未见报道。故本研究针对国内辣木生产上的实际问题,通过构建室内种群生命表,对不同辣木资源对辣木瑙螟的生存适合度进行评价,为辣木抗虫资源的选育和利用寄主抗虫性防治该害虫提供科学依据。
1材料与方法
1.1供试辣木种质资源
供试资源为农业农村部景洪辣木种质资源圃内的6种种质资源(表1)。
1.2供试昆虫
供试虫源采自云南省热带作物科学研究所中古辣木科技合作中心试验示范基地。将辣木瑙螟幼虫连同叶片一起采回在实验室饲养,待成虫羽化后,按1:1的雌雄比例多对放入养虫盒中。养虫盒为透明方形带盖塑料盒,长×宽×高=17.7cm×10.8cm×8.2cm,食品级PP(聚丙烯)材质。盒内放入含有5%~10%蜂蜜水的脱脂棉为其补充营养。用PE膜折叠成的小块(长×宽×高≈5cm×3cm×1.5 cm)供其产卵,饲养于温度(28±1)℃,相对湿度(75±10)%,光周期L∥D=14 h∥10 h的人工气候箱(BIC-300,上海博讯实业有限公司医疗设备厂;RQH-450,郑州生元仪器有限公司)内。待成虫交配产卵后,将卵取出放在盒子(透明方形带盖塑料盒,长×宽×高=15.5 cm×11.5 cm×6.5cm;材质同上)中,将盒子放入人工气候箱内待卵孵化,将初孵幼虫分别放入6个箱子(长×宽×高=64cm×46.5cm×36cm,材质同上)中,每个箱子分别放入不同资源新鲜辣木叶片,建立实验种群。
1.3辣木瑙螟的室内饲养及行为习性观察
以各资源饲喂的F3代新产的卵块为供试虫卵。早8点准时取出头天雌虫产的全部卵块,在显微镜下每个品种随机各数400粒,100粒为1组装入盒子中(透明密封带盖塑料盒,长×宽×高=3.7cm×2.9cm×3.3 cm,材质:食品级PP),放入温度(28±1)℃,相对湿度(75±10)%,光周期L∥D=14 h∥10 h的人工气候箱内饲养。每天早上8点观察1次,到次日早上8点记为1d,记录卵孵化情况。卵孵化后,将初孵幼虫分别接入放有足量新鲜辣木叶的培养皿(直径90 mm)中,100头为1组,每个培养皿接入1头幼虫,单头饲养并编号,设3个重复,即每份辣木资源饲养300头幼虫。每日更换新鲜辣木叶片,记录1~5龄幼虫的发育历期及存活情况;化蛹当天,用千分位电子天平(JJ623BC,Elektronische Waage)称量1日龄蛹重,记录预蛹期及蛹期。待成虫羽化后,记录其羽化时间、羽化数、雌雄虫数,然后以雌雄比1:1配对,放入含有5%~10%蜂蜜水的脱脂棉的盒子(长×宽×高=7.1cm×5.3cm×3.4 cm)内,并放入折叠成小块的PE膜供其产卵。观察记录产卵笼中每对成虫的产卵前期、产卵历期、逐日产卵量及雌雄虫死亡时间。
1.4数据统计与分析
2结果与分析
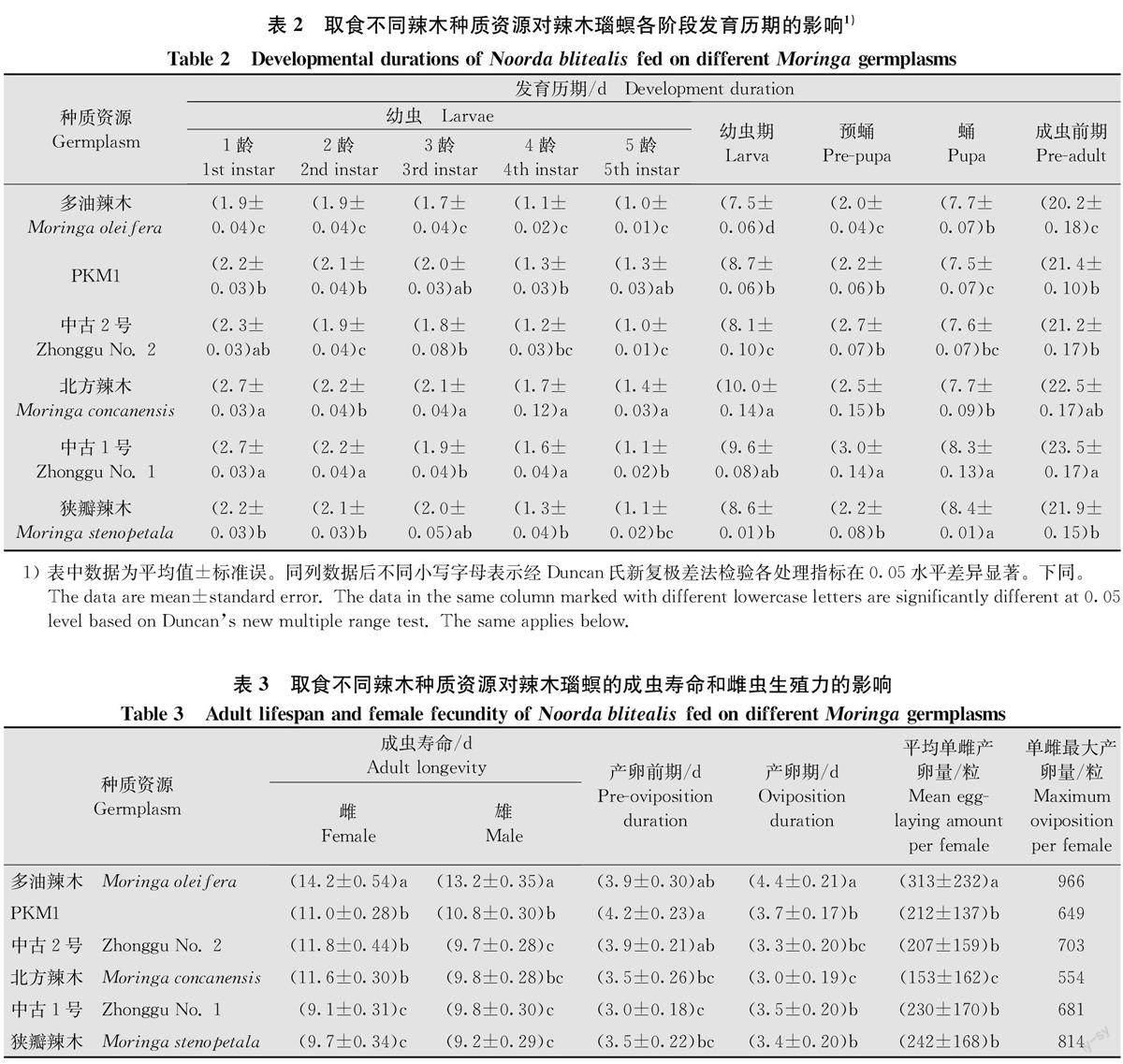
2.1取食不同辣木种质资源对辣木瑙螟各阶段发育历期的影响
在温度(28±1)℃,相对湿度(75±10)%,光周期L∥D=14h∥10 h条件下,取食6种辣木种质资源的辣木瑙螟卵期均为3.0d,辣木瑙螟幼虫在6种辣木种质资源上均为5龄,且均为1龄期最长,5龄期最短。取食不同辣木种质资源的幼虫历期存在显著差异(F=24.89,P=0.000),其中取食多油辣木的幼虫历期(7.5d)最短,分别比取食北方辣木和‘中古1号’短2.5d和2.1 d;取食其余3种资源的幼虫发育历期介于8.1~8.7d(表2)。
取食不同辣木种质资源的辣木瑙螟预蛹期存在显著差异(F=44. 04,P=0.000),取食‘中古1号’的预蛹期(3.0d)最长,取食多油辣木的预蛹期(2.0 d)最短;取食不同辣木种质资源的辣木瑙螟蛹期存在显著差异(F=5.94,P=0.005),取食‘中古1号’和狭瓣辣木的蛹期相对较长,分别为8.3d和8.4d,取食其余4种资源的蛹期介于7.5~7.7d;取食不同辣木种质资源的辣木瑙螟成虫前期存在显著差异(F=13.28,P=0.003),其中取食多油辣木的成虫前期最短,仅为20.2 d,分别比取食‘中古1号’和北方辣木短3.3 d和2.3d(表2)。
2.2取食不同辣木种质资源对辣木瑙螟成虫寿命和雌虫生殖力的影响
取食不同辣木种质资源的辣木瑙螟雌雌成虫寿命均存在显著差异(雌成虫:F=70.34,P=0.000;雄成虫:F=69. 69,P=0.000),其中取食多油辣木时雌雄成虫寿命分别为(14.2±0.54)d和(13.2±0.35)d,显著高于取食其余资源(P<0.05);取食不同辣木种质资源的辣木瑙螟产卵前期和产卵期均存在显著差异(产卵前期:F=9.42,P=0.000;产卵期:F5,12=18.74,P=0.000),其中取食‘PKM1’的成虫产卵前期(4.2 d)最长,取食其余5种种质资源的成虫产卵前期介于3.0~3.9 d;取食多油辣木的成虫产卵期(4.4 d)最长,取食北方辣木时最短(3.0 d);取食不同辣木种质资源的辣木瑙螟的平均单雌产卵量存在显著差异(F=82.36,P=0.000),其中取食多油辣木的单雌产卵量最高,为313粒,取食北方辣木时最低,为153粒,取食其余4种资源时单雌产卵量介于207~242粒;取食多油辣木的成虫最大单雌产卵量为966粒,取食其余5种资源时成虫最大单雌产卵量介于554~814粒(表3)。
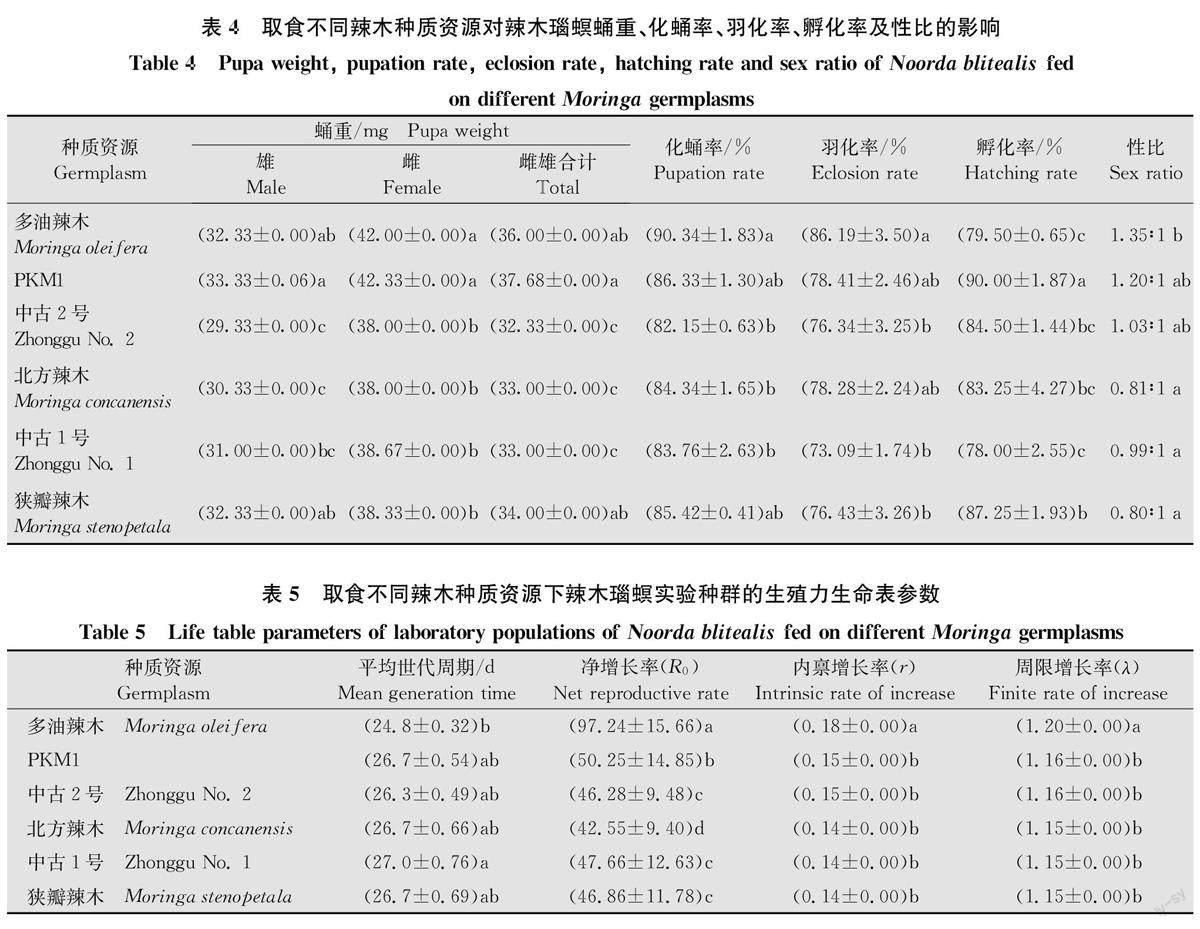
2.3取食不同辣木种质资源的辣木瑙螟蛹重、化蛹率、羽化率、孵化率及性比
取食不同辣木种质资源的辣木瑙螟蛹重存在显著差异(F=12.99,P=0.000),其中取食‘PKM1’的辣木瑙螟雌(42.33 mg)、雄(33.33 mg)虫蛹重均最重,取食‘中古2号’的雌(38mg)、雄(29.33 mg)虫蛹重均最轻;取食不同辣木種质资源的辣木瑙螟的化蛹率和羽化率均存在显著差异(化蛹率:F=3.67,P=0.03;羽化率:F=5.96,P=0.002),其中取食多油辣木时化蛹率和羽化率均最高,分别为90.34%和86.19%,取食其余5种资源时化蛹率介于82.15%~86.33%,羽化率介于73.09%~78.41% (P<0.05);取食不同辣木种质资源的辣木瑙螟的卵孵化率存在显著差异(F=10.79,P=0.000),取食‘PKM1’时卵孵化率最高,取食‘中古1号’和多油辣木时卵孵化率最低,取食其余3种资源时卵孵化率介于83.25%~87.25%(P<0.05)(表4)。以6种辣木种质资源饲养的辣木瑙螟的性比存在显著差异(F=56.21,P=0.000),雌雄性比介于0.80:1~1.35:1。
2.4取食不同辣木种质资源的辣木瑙螟实验种群生殖力生命表参数
辣木瑙螟取食6种辣木资源均能完成整个世代(表5),且周限增长率(λ)都大于1,说明取食这些资源后种群数量均以几何级数增长。幼虫取食多油辣木的世代周期(T)最短(24.8d),而取食‘中古1号’的最长(27.0d),二者相差2.2d。取食不同辣木种质资源的辣木瑙螟净增长率(Ro)差异显著(F=1294.70,P=0.000),取食多油辣木的R最高(97.24),是北方辣木的2.29倍,最有利于种群的增长。取食不同辣木种质资源的辣木瑙螟内禀增长率(r)差异显著(F=7.2,P=0.003),其中,取食多油辣木的辣木瑙螟r(0.18)和λ(1.20)均最高,显著高于取食其余5种资源。
3结论与讨论
实验种群生命表表明,虽然辣木瑙螟取食6种不同辣木种质资源均能完成整个世代发育,但其幼虫发育历期、预蛹期、蛹期、产卵前期、产卵期、世代历期、成虫寿命、产卵量均存在显著差异,说明这些种质资源作为辣木瑙螟寄主的适合度存在显著差异,即辣木瑙螟对6种辣木种质资源的适应性不同。
研究表明,存活率和发育历期可作为衡量寄主植物是否适宜植食性昆虫生长发育的指标[33]。取食多油辣木的辣木瑙螟的幼虫期最短,内禀增长率最高、产卵期长且繁殖力强,表明多油辣木对辣木瑙螟幼虫的适合度最高。鳞翅目昆虫蛹的质量是衡量其适应性的间接指标,一般情况下,蛹的质量越大其适应性越强[34]。如草地贪夜蛾的蛹重、雌成虫寿命、产卵量等都因取食玉米品种的不同而存在差异[35]。本研究通过对辣木瑙螟在6种资源上的生长发育情况进行分析亦得出类似结果,取食多油辣木的辣木瑙螟的蛹重、雌成虫寿命、化蛹率、羽化率、产卵量均高于取食‘中古2号’‘中古1号’和北方辣木的辣木瑙螟。辣木瑙螟对多油辣木的适应性最强,这与室外观察到辣木瑙螟对多油辣木辣木为害较严重相符。
研究表明,草地贪夜蛾在甘蔗、水稻、玉米上都可完成生活史,但在甜质型玉米上的生存适合度更好[36-37];小菜蛾在萝卜品种‘红衣天使’上的r、R和λ最小,说明该品种不利于小菜蛾的生长发育和繁殖[38]。本试验通过对实验种群生命表参数进行分析可知,取食6份辣木资源的辣木瑙螟均能完成整个世代,且周限增长率都大于1,说明取食这些资源后辣木瑙螟种群数量均以几何级数增长,但从反映种群世代增长数量的指标看,取食多油辣木的净增长率最高,为97.24,其次是‘PKM1’,为50.25,取食北方辣木的净增长率最低,为42.55,取食多油辣木的辣木瑙螟的平均世代周期最短(24.8d),且内禀增长率和周限增长率均为最高,表明取食多油辣木的辣木瑙螟种群增殖能力最强。
多油辣木原产于喜马拉雅山脉,是在世界范围内广泛引种栽培的品种[39],多油辣木有‘PKM1'和‘PKM2’2个栽培品种,其中‘PKM1’是通过传统方式选育的改良种[39],‘中古1号’是北方辣木和多油辣木的杂交种,‘中古2号’是‘中古1号’与多油辣木的杂交后代,北方辣木和‘中古1号’辣木的叶片比其余4种种质资源坚硬,结合6种种质资源生理特性及幼虫在各种质资源上的生长发育情况来看,辣木瑙螟在杂交种和改良种上的生存适合度显著低于多油辣木,多油辣木是6种辣木种质资源中最适合辣木瑙螟生长发育的种质资源,因此,可通过种质问的杂交创新来开展抗虫性研究,对辣木种质资源进行遗传改良,培育出更多的抗虫新品种。
综合生长发育及实验种群生命表参数来看,辣木瑙螟最适宜在多油辣木上生长发育,在此资源上辣木瑙螟种群具有更强的种群增殖能力,最不适宜在北方辣木上生长发育。结合辣木产量(叶片、果荚)及品质,可对‘PKM1’‘中古2号’‘中古1号’、狭瓣辣木和北方辣木这5种资源进行栽植推广。本试验结合辣木产业的发展,通过对辣木瑙螟在6种种质资源上生长发育情况进行分析,得出抗虫性较好的资源,可为辣木抗虫性品种的选育和推广提供一定的参考。
