贵州魔芋软腐病菌多基因分子鉴定及其致病力分析
2023-08-05黄露周兵正安星宇等
黄露 周兵正 安星宇等




关键词 魔芋;软腐病;病原鉴定;致病力
中图分类号:S 436.32 文献标识码:A DOI:10.16688/j.zwbh.2022328
魔芋Amorph,ophallus konjac是天南星科的一種多年生草本植物,主要分布在中国、日本及东南亚等国家和地区,其块茎富含葡甘聚糖,被广泛用于食品、医药等领域,是一种重要的经济作物,也是贵州省的特色产业之一[1-2]。随着魔芋大面积规模化种植,病害发生也越来越重,其中魔芋软腐病是魔芋生产中危害最严重的病害,可造成产量损失30%~50%,甚至绝收[3]。魔芋软腐病普遍发生于贵州魔芋种植区,已成为限制贵州魔芋产业发展的主要因素之一。
魔芋软腐病是由果胶杆菌Pectobacterium和迪基氏菌Dickeya等引起的细菌性病害,有学者认为其致病菌为胡萝卜欧文氏菌胡萝卜亚种Erwinia carotovora subsp.carotovora[4-5],修建华等通过16S- 23S rDNA转录间隔区分析,将魔芋软腐病菌分为胡萝卜欧文氏菌胡萝卜亚种、菊欧文氏菌Erwinia chrysanthemi及一株未能确定的菌株[6]。吴金平认为我国魔芋软腐病的病原菌主要为胡萝卜欧文氏菌胡萝卜亚种和菊欧文氏菌,同时分离到一株新的引起软腐病的细菌一肠杆菌属细菌En-terobacter sp.[7]。卢美欢等发现陕南地区魔芋软腐病的病原菌为胡萝卜果胶杆菌Pectobacterium ca-rotovorum[8]。孙苗苗研究发现引起湖北省魔芋软腐病的主要为海芋果胶杆菌Pectobacteriumaroidearum和方中达迪基氏菌Dickeya fang-zhongdai[9]。魏环宇等发现云南省珠芽魔芋软腐病的病原菌为海芋果胶杆菌[10-11]。而对贵州魔芋软腐病菌的研究,仅有本研究组前期发现黔东南魔芋软腐病菌为菊欧文氏菌E.chrysanthe-mi[12]。目前贵州省内许多地区引起软腐病的病原菌种类和分布仍不清晰,本研究采集贵州省主要魔芋种植基地的软腐病样,进行病菌的分离鉴定及致病力分析,明确贵州魔芋软腐病菌种类、致病力及分布特点,旨在为魔芋软腐病的防治工作提供科学依据。
1材料与方法
1.1试验材料和培养基
从贵州省威宁、水城、赫章、雷山等魔芋种植地采集发生软腐病的魔芋块茎、叶柄,装进采样袋中,标记采集信息,带回试验室备用。
营养琼脂培养基(NA):蛋白胨10g/L,牛肉膏粉5g/L,氯化钠5g/L,琼脂12g/L。营养肉汤培养基(NB):蛋白胨10g/L,牛肉膏粉3g/L,氯化钠5g/L。
1.2试验方法
1.2.1病原菌分离
采用平板划线法分离病原菌[13-14]。病样用75%乙醇表面消毒2 min,在无菌条件下挑取魔芋病样病健交界部分的小块组织,转移到含0.5 mL无菌生理盐水(0.85%NaCl)的无菌培养皿中,用灭菌玻璃棒将病组织捣碎,无菌条件下室温静置10~15 min,形成细菌悬浮液。用灭菌的移植环蘸取少量汁液在NA平板上划线,于28℃恒温培养箱中培养24h,挑取单菌落,转接纯化1~2次后,4℃和—80℃保存。
1.2.2病原菌的分子鉴定
1.2.2.1病原菌DNA提取及PCR扩增
采用天根细菌基因组DNA提取试剂盒提取病原菌基因组DNA,参考Bing等和Waleron等的方法[15-16],选用icdA、mdh、mtlD、proA、rpoS基因的引物(表1)进行PCR扩增。PCR扩增体系(25μL):2×Taq PCR Mix 12.5μL,10μmol/L的上、下游引物各1μL,DNA模板1μL,ddH20 9.5μL。PCR反应程序:95℃预变性5 min;95℃变性30s,退火温度下退火30s,72℃延伸1 min,35个循环;72℃延伸5min。PCR产物用1%琼脂糖凝胶电泳检测。将有清晰条带的样品送北京诺赛基因组研究中心有限公司测序。
1.2.2.2单基因和多基因序列分析及系统发育树构建
将测序获得的序列进行BLAST比对。采用BioEdit软件对从NCBI数据库中获取的可信度较高的相似序列进行多序列比对,基于icdA,mdh,mtlD,proA和rpoS 5个管家基因单个基因及多基因,用MEGA 6.0软件邻接法进行系统发育树构建和聚类分析。
1.2.3病原菌的致病力测定
分离的病原菌于NB培养液中,28℃,180 r/min培养24 h,配制成浓度1×10 cfu/mL的菌悬液,吸取2μL接种于用75%乙醇消毒的健康魔芋块茎(40 mm×40 mm)中央处,以无菌水为对照,2次重复,放于28℃培养箱中,培养24 h后观察发病情况,根据病斑的长度评价病菌的致病力,用Excel和DPS 7.5软件对数据进行统计分析,采用Duncan氏新复极差法(α=0.05)进行差异显著性分析。分级标准:病斑长度>30 mm为高致病(highly pathogenic level,H);10mm<病斑长度≤30 mm为中等致病力(moderately pathogenic level,M);病斑长度≤10 mm为低致病力(lowly pathogenic level,L),
2结果与分析
2.1魔芋软腐病原菌分离
通过对贵州赫章、威宁、雷山、平坝、息烽、花溪、荔波、关岭、水城等魔芋种植地的软腐病进行调查,发现植株于展叶期染病,叶柄顶部及基部首先出现湿润状暗绿色或黑色小斑(图1a),后整个叶柄溃烂,植株倒伏,散发恶臭味;植株基部或块茎染病时,早期叶片仍保持绿色,后整株逐渐变黄、干枯、倒伏,地下块茎呈灰色或黑灰色腐烂,并散发恶臭味(图1b~c)。对采集的各魔芋软腐病样进行病原菌分离、纯化,共获得47株菌株,回接魔芋块茎后都造成块茎腐烂,但不同菌株对魔芋块茎的致病程度不同(图1d~f)。
2.2单基因序列分析
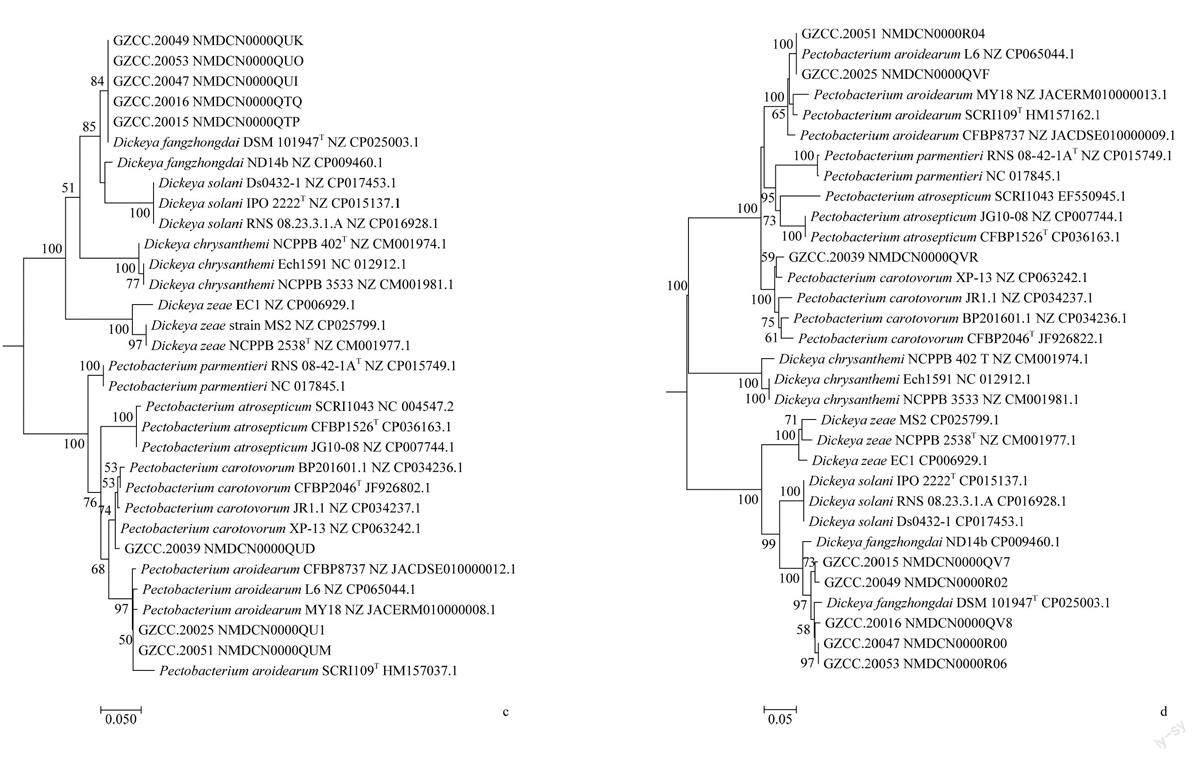
以分离的魔芋软腐病菌基因组总DNA为模板,分别进行icdA、mdh、mtlD、proA、rpoS等5个管家基因的PCR扩增及测序,并分别构建单基因系统发育树,发现不同基因构建的系统发育树对软腐病菌的鉴定结果有一定的差異,但所有菌株经鉴定均分属于为海芋果胶杆菌Pectobacterium aroidea-rum、胡萝卜果胶杆菌Pectobacterium carotovoorum或方中达迪基氏菌Dickeya fangzhongdai 3个种,其中根据mdh,mtlD,proA基因构建系统发育树结果基本一致,而采用icdA和rpoS基因时,菌株GZCC.20047、GZCC.20049和GZCC.20053鉴定结果有差异。基于icdA基因,GZCC.20049菌株与P.carotovorum聚为一支,GZCC.20053单独聚为一支;基于rpoS基因,菌株GZCC. 20047、GZCC.20053与P.aroidearum聚为一支(图2)。
2.3多位点序列分析
将分离的魔芋软腐病菌基因组的5个管家基因片段进行拼接、比对分析并构建多基因联合序列系统发育树(图3)。发现47个菌株有的聚在同个类群,有的则聚在不同类群,其中GZCC.20001、GZCC. 20003、GZCC. 20004等33个菌株与Pectobacterium aroidearum聚在同一个分支,同源性较高,均属于P.aroidearum;GZCC.20039菌株与Pectobacterium carotovorum聚类在同一个分支,属于P.carotovorum;而GZCC. 20002、GZCC. 20010、GZCC. 20011等13个菌株与Dickeya fangzhongdai聚类在同一个分支,为D.fangzhongdai。
2.4魔芋软腐病菌致病力测定
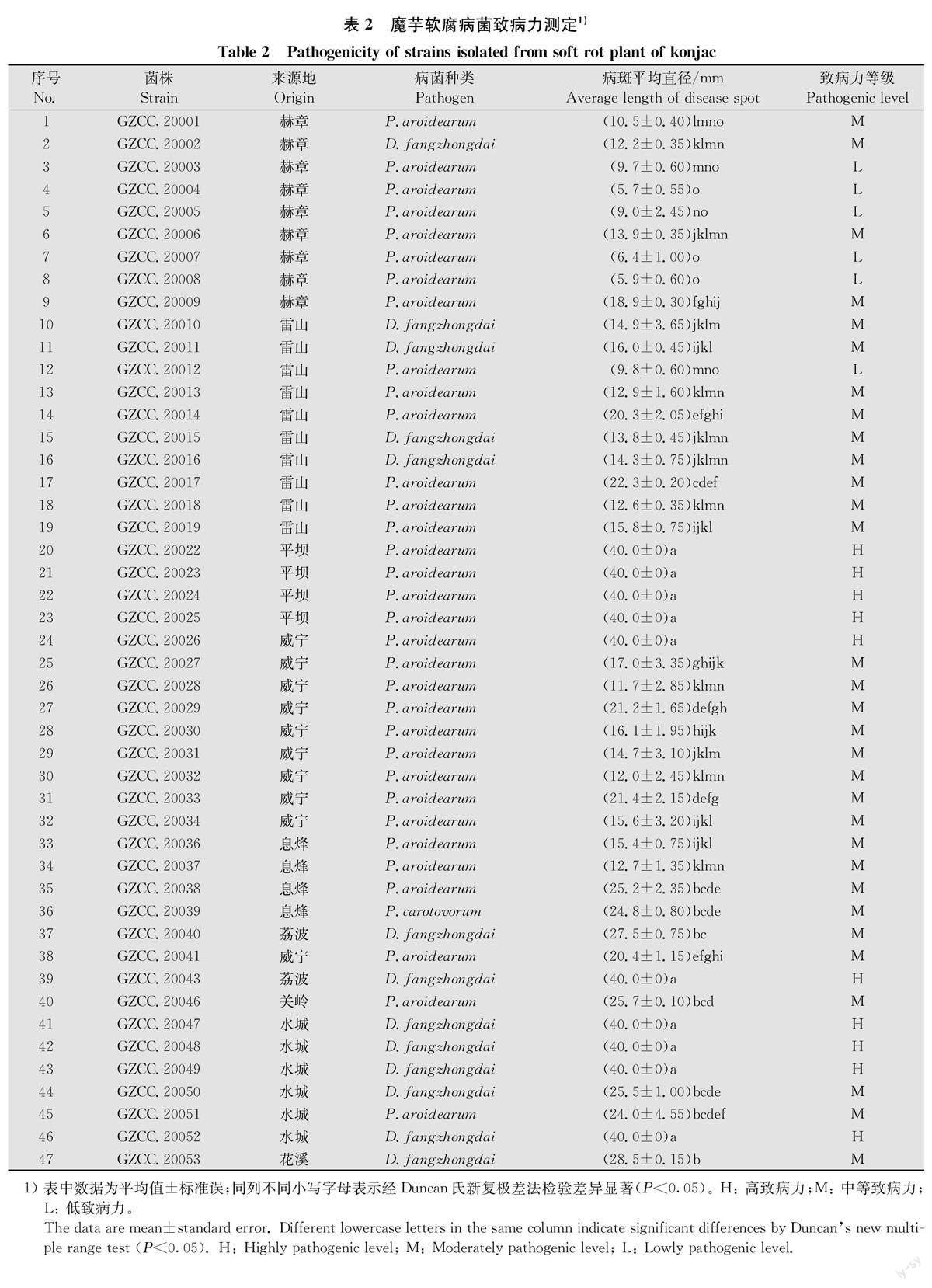
通过对魔芋软腐病菌进行致病力测定,发现不同软腐菌株的致病力有一定的差异。其中菌株GZCC. 20022、23、24、25、26、43、47、48、49和52号菌株具有强致病力,病斑直径达40 mm;GZCC. 20003、4、5、7、8、12号菌株致病力较弱,病斑直径在5. 7~9.8 mm;其余菌株的致病稍弱,病斑在10.5~28.5 mm间(表2)。
2.5贵州魔芋软腐病菌分布情况
通过对贵州魔芋软腐病病原菌进行鉴定,发现病菌分为3个种Pectobacterium aroidearum、P.ca-rotovorum和Dickeya fangzhongdai,其中P.aroidearum为贵州魔芋软腐病主要病原菌,约占分离菌株数的70%,广泛分布在多个地区;其次为D.fangzhongdai,占28%,也普遍存在于贵州魔芋种植区;P.carotovorum仅在息烽县魔芋种植地分离到1株,占2%。
3结论与讨论
随着分子生物技术的发展,近20年来,软腐病菌的分类发生很大的变化。1998年,欧文氏菌属Erwinia spp.有了重大修订,其许多种被归到果胶杆菌属Pectobacterium[17-18],随后在2005年Pecto-bacterium chrysanthemi被划定到新属迪基氏菌属Dicke ya[19-20]中。目前Pectobacterium属包括19个种,分别为猕猴桃果胶杆菌P.actinidiae,水生果胶杆菌P.aquaticum,海芋果胶杆菌P.aroidearum,黑茎果胶杆菌P.atrosepticum,甜菜果胶杆菌P.betavasculorum,巴西果胶杆菌P.brasiliense,仙人掌果胶杆菌P.cacticidum,胡萝卜果胶杆菌P.ca-rotovorum,泉水果胶杆菌P.fontis,气味果胶杆菌P.oderiferum,帕蒙蒂埃果胶杆菌P.parmentieri,微小果胶杆菌P.parvum,秘鲁果胶杆菌P.peruvi-ense,北极星果胶杆菌P.polare,波兰果胶杆菌P.polonicum,旁遮普果胶杆菌P.punjabense,广泛果胶杆菌P.versatile,山葵果胶杆菌P.wasabiae和马蹄莲果胶杆菌P.zantedeschiae;而Dickeya属有12个种,分别为菊迪基氏菌D. chrysanthemi,达旦迪基氏菌D.dadantii(subsp.dadantii and subsp.dieffenbachiae),石竹迪基氏菌D.dianthicola,玉米迪基氏菌D.zeae,水稻迪基氏菌D.oryzae,玉米迪基氏菌D.parazeae,茄迪基氏菌D.solani,方中达迪基氏菌D.fangzhongdai,水生迪基氏菌D.aquatica,湖泊迪基氏菌D.lacustris,水栖迪基氏菌D.undicola和喜禾本迪基氏菌D.poaceiphilan[18]。
本研究通过建立单基因和多基因系统发育树对贵州魔芋软腐病病原菌进行鉴定,发现贵州魔芋软腐病病原菌具有一定的多样性,包括海芋果胶杆菌P.aroidearum、胡萝卜果胶杆菌P.carotovorum和方中达迪基氏菌D.fangzhongdai 3个种,但一些菌株的单基因鉴定结果与多基因联合鉴定结果有一定差异,由于P.aroidearum、P.carotovorum和D.fangzhongdai 3个种比较相似,在历史分类地位上也常把其归为一类[17-19,21]。仅通过单个基因对其进行分类鉴定有一定的局限性,而采用多基因和全基因测序对其鉴定的准确性更好。本研究发现P.aroidearum为魔芋软腐病菌的主要种,广泛分布于贵州多个魔芋种植区,其菌株间致病力差异较大,低、中、高致病力菌株都有,而D.fangzhongdai菌株间差异较小,仅有中、高致病力菌株。
海芋果胶杆菌P.aroidearum在2013年被认定主要危害单子叶植物,但不仅限于单子叶植物的软腐病菌新种[22],近年来相继被报道危害马铃薯、西葫芦、白菜、魔芋、莴苣、辣椒、马蹄莲、胡萝卜、合果芋、甘蔗等作物[18,21,23-32]。对于魔芋软腐病的病原菌,以前报道多是胡萝卜欧文氏菌胡萝卜亚种Pecto-bacterium carotovora subsp.carotovora(P.carotovo-rum)和菊欧文氏菌Erwinia chrysanthemi (Dickeya sp.),但近年来通过全基因组及多基因序列分析,发现一些P.carotovorum其实是P.aroidearum,例如从陕西魔芋软腐样品中分离到一株细菌M8为P.aroidearum[21,33],魏环宇等发现云南省珠芽魔芋软腐病的致病菌也为P.aroidearum[10-11]。
本研究基于贵州魔芋软腐病菌种类不清的情况,确定了贵州魔芋软腐病菌的主要种类、致病力及分布情况,明确P.aroidearum、D.fangzhongdai是贵州魔芋软腐病的主要病菌,为魔芋软腐病的发生流行和科学防控提供了科学依据。
